
Sadia Saeed
Laboratory of Molecular Genetics, Department of Microbiology,
University ofKarachi-75270, Pakistan
Sheikh Ajaz Rasool
Laboratory of Molecular Genetics, Department of Microbiology,
University ofKarachi-75270, Pakistan
Samia Ahmad
Laboratory of Molecular Genetics, Department of Microbiology,
University ofKarachi-75270, Pakistan
Sohail Zahoor Zaidi
Public Health Laboratories, National Institute of Health, Islamabad, Pakistan
Shafqat Rehmani
Sindh Poultry Research Institute, Karachi, Pakistan
Research Journal of Microbiology
Year: 2007 | Volume: 2 | Issue: 11 | Page No.: 796-806







ABSTRACT
Staphylococcus aureus AB188 has been found to produce bacteriocins or/and bacteriocin-like inhibitory substance (BLIS) tentatively termed as staphylococcin 188. The staphylococcin 188 was purified to homogeneity by 80% ammonium sulfate precipitation and conventional size exclusion gel chromatography using Sephadex G-75 column equilibrated and eluted with 50 mM sodium phosphate buffer pH 7.0. This separation profile resulted in two major and well separated peaks designated as peak I and peak II. Bacteriocin activity was trailed in peak II with minor activity in peak I. Staphylococcin 188 had an estimated molecular weight of 4 kDa as indicated by activity detection after SDS-PAGE. The in vivo anti-viral studies of purified staphylococcin 188 against animal viruses were performed by chick embryo technique using New Castle Disease Virus (NCDV). Accordingly, the ELD50 (50% egg lethal dose) in case of virus injected system was found to be 10-9 while in the presence of staphylococcin 188; the ELD50 of virus dropped to 10-4. The in vitro anti viral studies of purified staphylococcin 188 against polio virus (propagated on Human Rhabdosarcoma cells (RD) and L20B cell lines (Mouse L cells cloned by human poliovirus receptors) were performed by serial tube dilution and 96 well plate method. Staphylococcin 188 did not show anti-viral activity against this single stranded Picorna (polio) virus.
PDF Abstract XML References
How to cite this article
Sadia Saeed, Sheikh Ajaz Rasool, Samia Ahmad, Sohail Zahoor Zaidi and Shafqat Rehmani, 2007. Antiviral Activity of Staphylococcin 188: A Purified Bacteriocin Like Inhibitory Substance Isolated from Staphylococcus aureus AB188. Research Journal of Microbiology, 2: 796-806.
URL: https://scialert.net/abstract/?doi=jm.2007.796.806
URL: https://scialert.net/abstract/?doi=jm.2007.796.806
INTRODUCTION
Antimicrobial peptides, (the bacteriocins), are non-replicating post translationally modified polypeptides, which have bactericidal or bacteriostatic effect, usually against bacteria that are closely related to the producer strain but not lethal to producer cells (Diep and Nes, 2002; Cleveland et al., 2001). Antimicrobial peptides have been shown to display both antiviral and antifungal activities in vitro and have also been shown to be effective in experimental infections with multidrug resistant Staphylococcus aureus and M. tuberculosis (Oakey et al., 2003). Bacteriocin-producer cells identified to date have exhibited specific immunity to the action of their own antimicrobial peptide. Staphylococci produce several types of antibacterial substances, which differ in their chemical nature and mode of action. Apart from lytic enzymes (such as lysostaphin and lysozyme), there exist two main groups of antagonists: I) bacteriocins (staphylococcins) and ii) low molecular weight antibiotic like substances. These staphylococcal bacteriocins are generally termed as staphylococcins or class I, (bacteriocins including lantibiotics containing modified amino acids) and class II, bacteriocins/BLIS which are small heat-stable non-lantibiotics. These two classes have been the subject of intensive study over the past few years. Several reports describe their production, purification and characterization. Similar to their structural heterogenities, staphylococcins produced by staphylococci show wide functional diversity and are active not only against other staphylococci but also against many other unrelated microorganisms. Nascimento et al. (2005) worked on Coagulase-Negative Staphylococcus (CNS) strains which were isolated from bovine mastitis cases and among them twelve CNS strains (6.4%) exhibited antagonistic activity against a Corynebacterium species and Listeria monocytogenes, an important food-borne pathogen and several strains of Streptococcus agalactiae associated with bovine mastitis, suggesting a potential use of these bacteriocins either in the prevention or in the treatment of streptococcal mastitis. The purification methods pertain to protein biochemistry and a wide variety of different combinations of procedures (usually applied with varying success). Often the crude bacteriocin preparations are first concentrated (precipitated) with acids, salts, ethanol or various solvent mixtures. Subsequent purification processes are based on size differences (gel chromatography, ultrafiltration and centrifugation) or charge differences [(ion-exchange chromatography, electrophoresis, isoelectric focusing) (Modak et al., 2004)]. New Castle Disease Virus (NCDV), a paramyxovirus is usually thought to be an avian (most notably related to chicken) virus, but it also infects humans. NCDV causes a potentially fatal, non-cancerous disease in birds; it causes only minor illness in humans (Seal et al., 2000). Poliovirus can enter the body either through the upper respiratory tract or the gastrointestinal tract (Grist et al., 1979). Individual strains of NCDV are classified as lytic or nonlytic. Viruses of both strain types can kill cancer cells, but lytic strains have the potential to do this more quickly because they damage the plasma membrane of infected cells. Nonlytic strains appear to kill by interfering with cell metabolism. The genetic material of NCDV is RNA rather than DNA (Schirrmacher et al., 1998). As with other types of viruses, essentially all of NCDV’s replication cycle takes place inside infected cells, which are also known as host cells. (Schirrmacher et al., 1997). During a replication cycle, new virus proteins and copies of the NCDV genetic material (i.e., genome) are made in the host cell’s cytoplasm. NCDV is also an enveloped virus, which means that progeny virus particles are released from infected cells by budding off from them. In this process, single copies of the NCDV genome become wrapped in an outer coat (i.e., an envelope) that is made from a small piece of the host cell’s plasma membrane (Eaton et al., 1973).
The conventional approach to the control of virus-associated diseases is to develop effective vaccines, but this is not always possible. At present, the viruses and bacteria are charging relentlessly more than 16 million lives per year everywhere and many countries, especially the poorest, constitute the main cause of death among children and adult youths (Modak et al., 2004). There is an increasing need of new compounds with antiviral activity as the treatment of viral infections with the available antiviral drugs is often unsatisfactory due to the problem of viral resistance coupled with the problem of viral latency and conflicting efficiency in recurrent infections in immunocompromised patients (Lin et al., 2001). In this situation, more attention needs to be paid to the search for antiviral agents present in natural products (Jassim and Naji, 2003). A large number of the biological activities of peptides isolated from different sources have been studied and that includes the antiviral activity. Enterocin CRL 35 produced by Enterococcus faecium CR 35 (isolated from cheese) inhibited viral multiplication of Herpes Simplex virus type 1 and 2 (HSV-1 and HSV-2) on Vero-cell lines in a dose dependent manner. Enterocin CRL35 also has anti-viral activity against Herpes Simplex virus type 1 and 2 in Vero and BHK-21 cells at concentrations 15-fold lower than the cytotoxic concentration (Waschmann et al., 1999).
MATERIALS AND METHODS
Partial Purification of Staphylococcin 188
For partial purification of staphylococcin 188, S. aureus AB188 was grown in brain heart infusion broth at 37°C for 24 h. The bacterial cells were removed by centrifugation at 10,000 rpm for 20 min at 4°C. The supernatant was concentrated 3 to 5 fold using a rotary evaporator followed by filter sterilization by passing it through 0.22 μm filters. Ammonium sulfate was added slowly to the supernatant with constant stirring at 4°C till the level of 80% concentration to attain the optimum precipitation of the protein. The system was held overnight and the precipitate was recovered by centrifugation at 15,000 rpm for 50 min at 4°C. The resulting pellet was solubilized in 200 mL of 50mM sodium phosphate buffer pH 7.0 and designated as crude preparation (Iqbal et al., 2001).
Molecular Weight Estimation of Staphylococcin 188
Size Exclusion Chromatography
The ammonium sulfate precipitated fraction was further subjected to conventional gel chromatography using Sephadex G-75 column equilibrated and eluted with 50 mM sodium phosphate buffer, pH 7.0. The flow rate was maintained at 3 mL/10 min and the elute was monitored at 280 nm. The active fractions thus obtained were collected and pooled for further bioassay for antimicrobial activity by agar well diffusion method (Magnusson and Schnurer, 2001; DeCourcy, 2004).
SDS-PAGE and Assay for Antibacterial Peptide
Fractions from ammonium sulfate precipitation and gel filtration chromatography were analyzed by sodium dodecyl sulfate-polyacrylamide gel electrophoresis [(SDS-PAGE) (Laemmli, 1970; Perbal, 1988)]. Calibration markers were used as molecular weight standard (Sigma), which include carbonic anhydrase (29 kDa), trypsinogen (24 kDa), trypsin inhibitor (21 kDa), myoglobin, horse heart (17 kDa), α-lactalbumin, bovine milk (15.5 kDa), bovine trypsin (6.2 kDa) and insulin chain B, oxidized, bovine (3.5 kDa). A 40 μL of the sample (with antibacterial activity), the sample treated with trypsin (Sigma) and the molecular weight standards were subjected to SDS-PAGE in vertical slab gels by the method of Laemmli (1970).
Gel Overlay Assay for Antimicrobial Activity Directly on SDS-PAGE
Direct detection of the antimicrobial activity by gel overlay method was performed as described by Bhunia et al. (1987). Briefly, after running a 12% gradient gel electrophoresis (SDS-PAGE) the gels were removed and cut into two vertical parts. One half (containing the samples and molecular weight standards) was stained with Coomassie Brilliant Blue R-250. The second half of the gel was assayed for antimicrobial activity by gel overlay method by placing the gel on a brain heart infusion agar plate and overlaid with 5 mL top agar (0.8% agar) containing 100 μL of exponential culture of indicator strain Enterococcus faecalis SA-74. After overnight incubation at 37°C, the plate-gel combination was examined for zones of inhibition.
Anti-viral Activity of Staphylococcin 188
Anti-viral activity of staphylococcin 188 against animal viruses
The in vivo anti-viral activity of staphylococcin 188 was performed by chick embryo technique (Beard et al., 1975). Ten fold dilutions of purified staphylococcin 188 and Moctasver New Castle Disease Virus (MNDV) were made in sterile PBS. For staphylococcin 188, dilutions were made up to 10-4 and for virus, dilutions were made up to 10-6. From each dilution of virus, 0.1 mL was injected into five eggs (6x5 = 30), which served as positive control. In eight tubes, 0.5 mL of bacteriocin from 10-3 tube was taken and mixed with 0.5 mL from each viral dilution. All tubes were incubated at 4°C for 30 minutes. After incubation, 0.1 mL was taken from each tube and injected into 5 eggs. Thus, from each tube, 0.5 mL was used to inject 5 eggs (8x5 =40). Five eggs without injection served as plain control. Five eggs were incubated as negative control (injected with 0.1 mL of PBS). All eggs were incubated at 37°C and mortality was observed after every 24 h (as per Brugh and Beard, 1980). After 96 h, haemagglutination (HA) was performed by taking allantoic fluid from eggs and 50% egg lethal dose (ELD50) was calculated.
Anti-viral Activity of Staphylococcin 188 Against Human Viruses
The in vitro anti-viral activity of staphylococcin 188 was performed by serial tube dilution and 96 well plate methods. The cell lines i.e., L20B (a genetically engineered mouse cell line expressing the human poliovirus receptor) and RD (derived from a human Rhabdosarcoma) were requested from Public Health Laboratories Division, National Institute of Health (NIH), Islamabad, Pakistan. Cell lines were grown as a monolayer culture in Hank’s Minimal Essential Medium (MEM) supplemented with Hank’s balanced salt solution (BSS-10) (ICN Biomedicals Inc.), 10% Fetal Calf Serum (FCS), 1 M HEPES and 200 mM L-glutamine (Gibco Invitrogen Corporation, Auckland UK), 100 IU mL-1 penicillin, 100 μg mL-1 streptomycin, 7.5% NaHCO3 solution and 0.4% phenol red solution (ICN, Sigma). The maintenance medium contained the same material as the growth medium contained but the concentration of fetal calf serum was set as 2%. The cells were maintained at 36°C in a humidified atmosphere with 5% CO2 and were subcultured twice a week (Waschmann et al., 1999; Lin et al., 2001).
Cytotoxicity Assay of Staphylococcin 188
To establish the most appropriate working concentration of staphylococcin 188 (for its anti-viral activity against human viruses), the cytotoxicity assay was performed. Virus titer was determined by the 50% Tissue Culture Infectious Dose [(TCID50) (the amount of viruses required to kill 50% cells in a cell line)] assay using monolayers of L20B and RD cell lines in 96-well microtiter plate. The two-fold and four-fold dilutions of purified staphylococcin 188 were made in cell broth of Hanks minimal essential medium supplemented with 2% fetal calf serum and added to the monolayers of L20B and RD cell lines. The control cell lines were run alone (without adding staphylococcin 188). All cell lines were incubated at 36°C and observed for 5 days. All the experiments were run in triplicate (Murinda et al., 2003; Modak et al., 2004).
Anti-viral Activity Assay of Purified Staphylococcin 188
The anti-viral activity assay of purified staphylococcin 188 was performed according to standard procedure in 96 well tissue culture plates (Rubinstein et al., 1981). For this, two-fold serial dilutions of the purified staphylococcin 188 were made. The growth medium was aspirated from established monolayers of cells in 98-well tissue culture plates. Viruses with known TCID50 were added to each well except those to be used for cell controls. After the viruses were adsorbed at 36°C, the appropriate dilutions of staphylococcin 188 were added to each test wells (3 wells/dilution). Tissue culture medium without drug was added to cell control wells (without virus) and virus control wells (with virus). Test medium containing drug without virus was added to all toxicity control wells. The plates were incubated at 37°C in an atmosphere of CO2 until virus control wells had Cytopathic effects (CPE) near 100%. Morphological changes in the test wells were compared with the control wells and examined microscopically under inverted microscope. Morphological changes were graded on a scale of 0-4, with 0 being no effect and 4 being 100% cell death. All experiments were performed in triplicate (Modak et al., 2004).
RESULTS
The addition of ammonium sulfate to a final concentration of 80% resulted in precipitation of 80% of the total staphylococcin 188 activity and 4 fold increase in the specific activity. Precipitates were concentrated in a minimal volume by rotary evaporator in a minimal volume and diluted to a known protein concentration in 50 mM sodium phosphate buffer pH 7.0 and designated as Staphylococcin 188 as assayed by agar well diffusion using Enterococcus faecalis SA-74 as indicator strain (Table 1). The purification steps resulted in specific activity of 250 AU mg-1 and a recovery of 78% of total activity. The crude ammonium sulfate precipitate was further subjected to conventional gel filtration chromatography on Sepahdex-G75 column equilibrated and eluted with 50 mM sodium phosphate buffer, pH 7.0. This separation profile resulted in two major and well separated peaks designated as peak I and peak II. Bacteriocin activity was trailed in peak II with minor activity in peak I (Fig. 1). After chromatography, antibacterial activity of both these peaks was checked by agar well diffusion method using Enterococcus faecalis SA-74 (Fig. 2) as indicator culture. This purification step resulted in maximum specific activity of 5000AU mg-1 and a recovery of 7.8% of the total activity (Table 1). Ammonium sulfate precipitated and purified fractions of staphylococcin 188, eluted from size exclusion gel chromatography were subjected to SDS-PAGE. Figure 3 shows the electrophoregram of staphylococcin 188. In the case of ammonium sulfate precipitation, six major and other bands were observed with molecular weights ranging from 29 to 3.5 kDa (Fig. 3a). The bacteriocin activity against Enterococcus faecalis SA-74 corresponded to a low molecular weight band [(with an estimated molecular mass of 4 kDa) (Fig. 3b)]. Purified staphylococcin 188 (fraction II) from gel chromatography migrated in CBB-stained SDS-PAGE as a single band (Fig. 3c). The bacteriocin activity on SDS-PAGE against Enterococcus faecalis SA-74 (Fig. 3d) was exhibited as a clear zone of inhibition on the corresponding band (4 KDa). The in vivo anti-viral activity of purified staphylococcin 188 in chick embryo system was evaluated against NCDV by 50% egg lethal dose (ELD50). Accordingly, the ELD50 in case of virus injected system was found to be 10-9 while in the presence of staphylococcin 188; the ELD50 of viruses was found to be 10-4.
| Table 1: | Summary of purification of staphylococcin 188 from culture supernatant of Staphylococcus aureus AB188 |
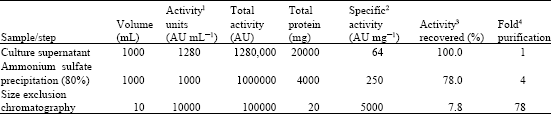 | |
| 1Activity unit (AU mL-1) = Reciprocal of the highest dilution x 1000/volume of bacteriocins added. 2Specific activity (AU mg-1) = Total activity of the subsequent purification step/Total protein of the same step. 3Recovery (%) = Total activity of subsequent step x 100/Total activity of crude preparation. 4Fold purification = Specific activity of subsequent step/ Specific activity of crude preparation. Note: The specific activity (AU mg-1) and activity recovered (%) and times purified are expressed in appropriate whole numbers | |
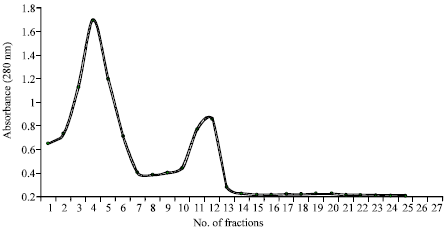 | |
| Fig. 1: | Size Exclusion Chromatography (SEC) of partially purified precipitates of staphylococcin 188 subjected to Sephadex G-75 gel filtration column. Column was equilibrated and eluted with 50 mM Sodium Phosphate buffer, pH 7.0. Flow rate of the column was maintained at 3 mL/10 min and the elute was monitored at 280 nm. Bioactivity of both peaks was checked by agar well diffusion method against Enterococcus faecalis SA-74 |
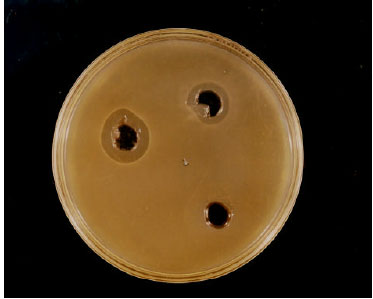 | |
| Fig. 2: | Staphylococcin 188 profile of 2 peaks obtained after size exclusion gel chromatography. Both peaks were collected separately and activity of each peak was checked by agar well diffusion assay. The well 1 contains sample obtained from peak I and well 2 contains sample obtained from peak II. Well 3 served as control (PBS). The target organism was Enterococcus faecalis SA-74 |
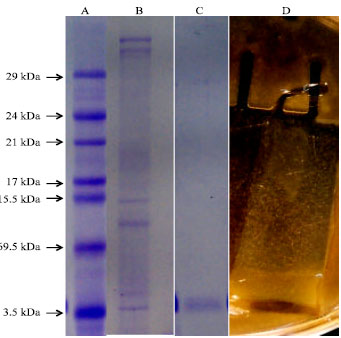 | |
| Fig. 3: | Twelve percent gradient polyacrylamide gel electrophoresis of the staphylococcin 188 preparation after ammonium sulfate precipitation and size exclusion gel chromatography. A: Coomasie brilliant blue R-250 stained gel: lane 1, molecular weight markers, B: Lane 2, ammonium sulfate precipitated staphylococcin 188, C: Lane 3, active fraction obtained after size exclusion gel chromatography and D: Detection of staphylococcin 188 activity on SDS-PAGE against Enterococcus faecalis SA-74 |
(The difference between both ELD50 doses is approximately thousand fold). These observations indicate that in the absence of bacteriocin, the viruses had infected chick embryo at high dilution (less viral load) while in the presence of bacteriocin, low titre of virus (high viral load) was required (ELD50) to infect chick embryo (Table 2 and 3).
| Table 2: | Haemagglutination (HA) profile using eggs inoculated with NCDV (New Castle Disease Virus) and NCDV + Staphylococcin 188 system |
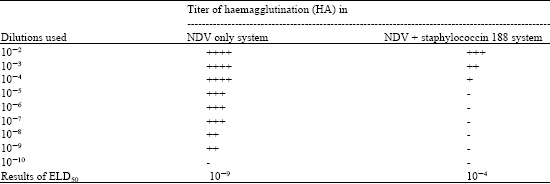 | |
| ++++ = Prominent agglutination; +++ = Moderate agglutination; ++ = Fair agglutination; + = Less agglutination and - = No agglutination | |
| Table 3: | In vivo anti-viral activity of purified staphylococcin 188 on Newcastle Disease Virus (NDV) by chick embryo technique |
 | |
| Note: The numbers indicates the number of eggs killed after every 24 h (after injection of different dilutions of staphylococcin 188 and NDV) | |
| Table 4: | Cytotoxic effects of staphylococcin 188 on RD and L20B cell lines (to check the antiviral activity against Polio virus) |
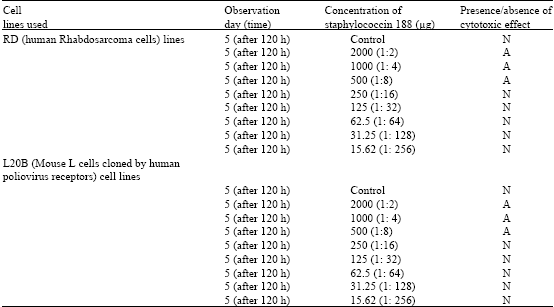 | |
| Control = cell lines without staphylococcin 188, A = Abnormal cell lines, N = Normal cell line | |
The in vitro anti-viral studies of purified staphylococcin 188 against polio viruses propagated on RD and L20B cell lines were performed by serial tube dilution and 96 well plate method at a concentration non toxic to these cell lines. The experiments were performed in triplicate. The results obtained indicated that staphylococcin 188 did not show antiviral activity against this single strand RNA (polio) virus (Table 4).
DISCUSSION
Bacteriocins of Gram-positive bacteria have been examined extensively in the recent years (Jack et al., 1995; Ennahar et al., 2000). Bacteriocins produced by Gram-positive bacteria exhibit a wider spectrum of inhibitory activities, which may favour a broader industrial application (Eijsink et al., 1998). The staphylococcin 188 was purified from culture supernatant of Staphylococcus aureus AB188 by sequential ammonium sulfate precipitation and size exclusion gel chromatography. Salting out or ammonium sulfate precipitation is useful for concentrating dilute solutions of proteins (Whitford et al., 2001). It is also useful for fractionating a mixture of proteins. Usually an ammonium sulfate cut is taken in order to obtain a higher degree of purification. Precipitation of culture supernatant of Staphylococcus aureus AB188 with ammonium sulfate resulted in 80% recovery of the total staphylococcin 188 activity. The specific activity in crude culture supernatant was 64 (AU mg-1) which was increased to 250 (AU mg-1) after ammonium sulfate precipitation and the purification was 4-fold with a recovery of 78%. Ogunbanwo et al. (2003) reported that bacteriocins of L. plantarum F1 and L. brevis OG1 were recovered following the 60% saturation of the culture broths with ammonium sulfate with an increased specific activity of 9.4 and 5.2 AU mg-1 protein, respectively. The ammonium sulfate precipitates were further subjected to conventional gel filtration chromatography on Sepahdex-G75 column equilibrated and eluted with 50 mM sodium phosphate buffer, pH 7.0. The antibacterial activity was determined by analysis on Enterococcus faecalis SA-74 plate and the bacteriocin activity resided in peak II with minor activity in peak I. Size exclusion gel chromatography resulting increased in the specific activity (5000AU mg-1) compared to that obtained after ammonium sulfate precipitation (250 AU mg-1) and with a recovery of 7.8% of the total antibacterial activity from 78% following the ammonium sulfate precipitation (Table 1). The recovery of the activity of staphylococcin 188 decreased with the subsequent step in purification. In case of staphylococcinT, the specific activity of ammonium sulfate precipitates was 801AU mg-1 which was increased up to 4690 AU mg-1 after gel filtration chromatography and the final purification was 82-fold from 14-fold (Furmanek et al. 1999). The method used for detecting bacteriocin activity directly in SDS-PAGE is the simplest, sensitive, rapid and the most widely used protocol (Bhunia et al., 1987; Crupper and Iondolo, 1996). The corresponding band giving inhibitory activity can be eluted from the gel for further studies. During the course of detection of staphylococcin 188 activity directly in the gel, same methodology (Bhunia et al., 1987) with slight modifications was followed. The electrophoregram of CBB-stained gel showed a heterogenous protein population ranging from 29 to 3.5 kDa. In the gel used for detection of antibacterial activity, a clear zone of inhibition of the indicator cells (Enterococcus faecalis SA-74) corresponds with a 4 kDa protein, indicating that staphylococcin 188 could be associated with low molecular mass protein. The low molecular weight proteins have been reported in staphylococci and other Gram-positive bacteriocins such as staphylococcin from S. aureus IYS2 [(5 kDa) (Nakamura et al., 1983)], Bac1829 from S. aureus KSI1829 [(6.4 kDa) (Crupper and Iondolo, 1996)], staphylococcin BacR1 from S. aureus UT0007 [(303 kDa) (Crupper et al., 1997)], staphylococcin T from Staphylococcus cohnii T strain [(2.1 kDa) (Furmanek et al., 1999)]. The in vivo anti-viral studies of staphylococcin 188 against animal viruses were performed by chick embryo technique using NCDV. According to the results, the ELD50 (50% egg lethal dose) in case of virus injected system was found to be 10-9 while in the presence of staphylococcin 188; the ELD50 of viruses was dropped to 10-4 indicating that in the absence of bacteriocin, the viruses had infected chick embryo at high dilution (less viral load) and in the presence of bacteriocin, low titer of virus (high viral load) was required (ELD50) to infect chick embryo. This is the first report in which the bacteriocin from Staphylococcus aureus was studied against any animal (avian) virus. New Castle Disease (NCD) is a highly contagious viral disease of domestic poultry, cage and wild birds. It is characterised by digestive, respiratory and/or nervous signs, inflammation of respiratory tract and of either the brain or the gastrointestinal tract. NCD virus is infective for almost all avian species (Schirrmacher et al., 1997, 1998). The anti-viral activity of staphylococcin 188 against NDV is an interesting finding and these results may convey that this bacteriocin may be used for the treatment of avian diseases (hens or ducks) to eliminate or minimize the heavy economic losses occuring due to infections of chicken (caused by NDV). The in vivo anti-viral studies of purified staphylococcin 188 were conducted against human virus. For these studies, polio virus was used as a source of human associated virus. Initially, cytotoxic studies were done. It is very important to check the cytotoxic level of the bacteriocin before its use as a preservative or in the treatment of infections. In case of staphylococcin 188, two fold and four fold dilutions were made and incubated with Human Rhabdosarcoma cells (RD) and L20B cell lines using 96 well plate method. Cytotoxic effect was checked after every 24 h and up to 120 h. Addition of staphylococcin 188 up to a concentration of 250 μg mL-1 to the culture medium had no significant cytotoxic effect on cell lines. The results indicated toxic effects of bacteriocins in dose-dependent manner (i.e., 500, 1000 and 200 μg mL-1). Murinda et al. (2003) have used different preparations of partially purified bacteriocins (Nisin, pediocin, colicins E1, E3, E6, E7 and K) to check their cytotoxic effects on Simian virus 40-transfected human colon (SV40-HC) and Vero monkey kidney (Vero) cells. After estimating the cytotoxic level of staphylococcin 188, the anti-viral activity of staphylococcin 188 against polio virus was studied on RD and L20B cell lines by serial tube dilution and 96 well plate method. Staphylococcin 188 did not show anti-viral activity against this single strand RNA (polio) virus. In conclusion, we could say that staphylococcin 188 has no anti-viral activity against the single stranded RNA virus. Bacteriocin with demonstrable anti-viral activity has been reported mostly against HSV (Herpes Simplex virus) because of its structure and super sensitivity to many antiviral agents (Waschmann et al., 1999; Kott et al., 1999). To our knowledge no reports about the anti-viral activity of bacteriocins against polio virus (single stranded RNA virus) have been published. A large number of the biological activities including the anti-viral ones of peptides isolated from different sources have been studied. Enterocin CRL35, a bacteriocin isolated from Enterococcus faecium had strong antiviral activity against HSV-1 and HSV-2 (Wachsman et al., 2003).
ACKNOWLEDGMENT
This project has been supported by Pakistan Science Foundation (PSF-S-KU/CHEM-363) and Higher Education Commission (HEC No. 20-598/R and D/06/1650) research grants to Sheikh Ajaz Rasool.
REFERENCES
- Beard, C.W., S.R. Hopkins and J. Hammond, 1975. Preparation of Newcastle disease virus hemagglutination-inhibition test antigen. Avian Dis., 19: 692-699.
CrossRefDirect Link - Bhunia, A.K., M.C. Johnson and B. Ray, 1987. Direct detection of an antimicrobial peptide of Pediococcus acidilactici in sodium dodecyl sulfate-polyacrylamide gel electrophoresis. J. Ind. Microbiol., 2: 319-322.
CrossRefDirect Link - Brugh, M.J. and C.W. Beard, 1980. Collection and processing of blood samples dried on paper for microassay of Newcastle disease virus and avian influenza virus antibodies. Am. J. Vet. Res., 41: 1495-1498.
PubMed - Cleveland, J., T.J. Montville, I.F. Nes and M.L. Chikindas, 2001. Bacteriocins: Safe, natural antimicrobials for food preservation. Int. J. Food Microbiol., 71: 1-20.
CrossRefPubMedDirect Link - Crupper, S.S. and J.J. Iandolo, 1996. Purification and partial characterization of a novel antibacterial agent (Bac1829) produced by Staphylococcus aureus KSI1829. Applied Environ. Microbiol., 62: 3171-3175.
Direct Link - Crupper, S., A.J. Gies and J.J. Iandolo, 1997. Purification and characterization of staphylococcin BacR1, a broad-spectrum bacteriocin. Applied Environ. Microbiol., 63: 4185-4190.
Direct Link - Diep, D.B. and I.F. Nes, 2002. Ribosomally synthesized antibacterial peptides in Gram-positive bacteria. Curr. Drug Targets, 3: 107-122.
PubMedDirect Link - Eaton, M.D., J.A. Heller and A.R. Scala, 1973. Enhancement of lymphoma cell immunogenicity by infection with nononcogenic virus. Cancer Res., 33: 3293-3298.
Direct Link - Eijsink, V.G.H., M. Skeie, P.H. Middelhoven, M.B. Brurberg and I.F. Nes, 1998. Comparative studies of class IIa bacteriocins of lactic acid bacteria. Applied Environ. Microbiol., 64: 3275-3281.
Direct Link - Ennahar, S., N. Deschamps and J. Richard, 2000. Natural variation in susceptibility of Listeria strains to class IIa bacteriocins. Curr. Microbiol., 41: 1-4.
Direct Link - Furmanek, B., T. Kaczorowski, R. Bugalski, K. Bielawski, J. Bogdanowicz and A.J. Podhajska, 1999. Identification, characterization and purification of the lantibiotic staphylococcin T, a natural gallidermin variant. J. Applied Microbiol., 87: 856-866.
PubMedDirect Link - Iqbal, A., S.A. Ali, A. Abbasi, W. Volter and S.A. Rasool, 2001. Production and some properties of Bac201: A bacteriocin like inhibitory substance from Staphylococcus aureus AB201. J. Basic Microbiol., 41: 25-36.
PubMedDirect Link - Jack, R.W., J.R. Tagg and B. Ray, 1995. Bacteriocins of gram-positive bacteria. Microbiol. Rev., 59: 171-200.
Direct Link - Jassim, S.A.A. and M.A. Naji, 2003. Novel antiviral agents: A medicinal plant perspective. J. Applied Microbiol., 95: 412-427.
CrossRefPubMedDirect Link - Kott, V., L. Barbini, M. Cruane, J.D. Munoz, E. Vivot, J. Cruanes, V. Martino and G. Ferraro, 1999. Antiviral activity in Argentine medicinal plants. J. Ethnopharmacol., 64: 79-84.
PubMedDirect Link - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Lin, S.B., C.H. Lin and Y.L. Hsu, 2001. Establishment of an antiviral activity assay and the partial purification of interferon-like protein from rainbow trout gonadal cells (RTG-2). Zoo. Stu., 40: 240-246.
Direct Link - Maganusson, J. and J. Schnurer, 2001. Lactobacillus coryniformis subsp. coryniformis strain Si3 produces a broad-spectrum proteinaceous antifungal compound. Appl. Environ. Microbiol., 67: 1-5.
CrossRefDirect Link - Modak, B., H. Galeno and R. Torres, 2004. Antiviral activity on Hantavirus and apoptosis of vero cells of natural and semi-synthetic compounds from Heliotropium fulifolium resin. J. Chil. Chem. Soc., 49: 143-145.
Direct Link - Murinda, S.E., K.A. Rashid and R.F. Robert, 2003. In vitro assessment of the cytotoxicity of nisin, pediocin and selected colicins on simian virus 40-transfected human colon and Vero monkey kidney cells with trypan blue staining viability assays. J. Food Prot., 66: 847-853.
PubMedDirect Link - Nascimento, D.S.J., P.C. Fagundes, D.P. Brito and K.R. Santos, 2005. Production of bacteriocins by coagulase-negative staphylococci involved in bovine mastitis. Vet. Microbiol., 106: 61-71.
CrossRefDirect Link - Ogunbanwo, S.T., A.I. Sanni and A.A. Onilude, 2003. Characterization of bacteriocin produced by Lactobacillus plantarum F1 and Lactobacillus brevis OG1. Afr. J. Biotechnol., 2: 219-227.
Direct Link - Schirrmacher, V., C. Haas and R. Bonifer, 1997. Virus potentiation of tumor vaccine T-cell stimulatory capacity requires cell surface binding but not infection. Clin. Cancer Res., 3: 1135-1148.
Direct Link - Schirrmacher, V., T. Ahlert and T. Probstle, 1998. Immunization with virus-modified tumor cells. Semin. Oncol., 25: 677-696.
Direct Link - Seal, B.S., D.J. King and H.S. Sellers, 2000. The avian response to New Castle Disease Virus. Dev. Comp. Immunol., 24: 257-268.
PubMedDirect Link - Wachsman, M.B., M.E. Farias, E. Takeda, F. Sesma, A.P. de Ruiz Holgado, R.A. de Torres and C.E. Coto, 1999. Antiviral activity of enterocin CRL35 against herpesviruses. Int. J. Antimicrob. Agents, 12: 293-299.
Direct Link - Whitford, W.F., M.A. McPherson, R.J. Foster and R.M. Teather, 2001. Identification of bacteriocin-like inhibitors from rumen Streptococcus spp. and isolation and characterization of Bovicin 255. Applied Environ. Microbiol., 67: 569-574.
CrossRefDirect Link