
Kustiati
School of Life Sciences and Technology, Institut Teknologi Bandung, Bandung, Indonesia
Marselina I. Tan
School of Life Sciences and Technology, Institut Teknologi Bandung, Bandung, Indonesia
Sri Yusmalinar
School of Life Sciences and Technology, Institut Teknologi Bandung, Bandung, Indonesia
Trisnowati B. Ambarningrum
Faculty of Biology, Universitas Jenderal Soedirman, Purwokerto, Indonesia
Intan Ahmad
School of Life Sciences and Technology, Institut Teknologi Bandung, Bandung, Indonesia
Journal of Entomology
Year: 2016 | Volume: 13 | Issue: 1-2 | Page No.: 40-47







ABSTRACT
House flies, Musca domestica L. is a major and the most common urban insect pest in Indonesia and is known to have the ability to develop resistance to insecticides. In order to determine the resistance status of this pest to permethrin and imidacloprid, the most commonly used insecticides to control this pest, 32 strains of house flies collected in 2013 and 2014 from 26 capital of provinces in Indonesia were tested. House flies were tested by topical application and feeding bioassay for resistance to permethrin and imidacloprid, respectively. To examine the resistance mechanism to permethrin, the synergist piperonyl butoxide was used. Majority of all strains, 26 of 32 field-collected strains showed very high levels of resistance to permethrin, the range of resistance ratios was 190-25190 fold for permethrin when compared with a susceptible strain. However, a few strains showed no to high levels of resistance to permethrin, with the range ratio was 0.3-143 fold. Meanwhile, for imidacloprid, all strains showed no to very low resistance (resistance ratios of 0.4-6.1 fold). Assay using permethrin in the presence of PBO in 12 of 14 selected strains were found to decrease the LD values, which suggests that the detoxifying enzyme Mixed Function Oxidase (MFO) might have a role in the development of resistance to permethrin. Our study is the first report of house flies resistance throughout much of Indonesia, although all strains were still susceptible to imidacloprid. The findings, together with house flies integrated management, can be used to prevent resistance development in other insecticides.
PDF Abstract XML References Citation
Received: December 14, 2015;
Accepted: January 27, 2016;
Published: February 15, 2016
How to cite this article
Kustiati, Marselina I. Tan, Sri Yusmalinar, Trisnowati B. Ambarningrum and Intan Ahmad, 2016. Monitoring Permethrin and Imidacloprid Resistance in Indonesian House Fly Musca domestica L. (Diptera: Muscidae). Journal of Entomology, 13: 40-47.
DOI: 10.3923/je.2016.40.47
URL: https://scialert.net/abstract/?doi=je.2016.40.47
DOI: 10.3923/je.2016.40.47
URL: https://scialert.net/abstract/?doi=je.2016.40.47
INTRODUCTION
The house fly, Musca domestica L. (Diptera: Muscidae) is considered a pestiferous nuisance and important public health threat due to their potential role as mechanical vectors of ≥100 diseases causing pathogens, e.g., typhoid, cholera, dysentery and diarrhea and viral infections (Keiding, 1986; Scott et al., 2014). Even though it was known that environmental sanitation was the most important aspect in controlling the house flies, the use of insecticides remain the first choice of controlling method. Unfortunately, the intensive use of insecticides has resulted in the development of resistance to not only all four major classes of insecticides (organochlorine, organophosphates, carbamates and pyrethorids) (Marcon et al., 2003; Akiner and Caglar, 2006; Cao et al., 2006; Memmi, 2010; Scott et al., 2013; Abbas et al., 2015a) but also to the relatively new class of insecticide, neonicotinoids.
Resistance to neonicotinoids in house flies has been reported in recent year in some countries. For example, in Denmark, moderate to high levels of resistance to imidacloprid were found in livestock units (Kristensen and Jespersen, 2008). Whereas, no to low levels of resistance to imidacloprid were found in five dairies farm in Florida (Kaufman et al., 2010b) and in five poultries facilities in Pakistan (Abbas et al., 2015b).
In Indonesia, since 1980s, home owners, public health officials and many professional Pest Control Operators (PCOs) used pyrethroids to control house flies due to their fast action and relatively low mammalian toxicity. However, due to the possibility that house flies in Indonesia had developed resistance to the pyrethroids and also environmental concerns, bait containing relatively new insecticide, imidacloprid was introduced as alternative in the late of 2000s.
Although some anecdotal reports from the PCOs has suggested house flies in some parts of Indonesia have developed resistance against pyrethroids, data about resistance status of house flies in Indonesia remain very limited or poorly documented. For example, Susanti in 2010, reported 6.2 fold resistance of houseflies, collected from Bandung to permethrin and also suggested that detoxifying enzyme Mixed Function Oxidase (MFO) played an important role in the development of resistance to permethrin. However, her findings in terms of resistance ratio (6.2 fold) might have been lower than the expected, due to the fact that the Bandung strain was not compared with the standard insecticide susceptible ones.
Therefore, given the limited information on the resistance of house flies to insecticides in Indonesia, the purpose of this study was conducted to determine the current resistance status of permethrin and imidacloprid to houseflies which were collected from 26 Indonesian provinces as well as testing the possible mechanisms that was involved in the development of resistance to permethrin by using piperonyl butoxide as synergist (Astari and Ahmad, 2005). In addition, this study was expected to provide baseline data for future monitoring effort that will help to define a control strategy for management of house flies resistance to insecticide in Indonesia.
MATERIALS AND METHODS
House flies: Larvae and adult house flies, Musca domestica were collected from garbage dumps from the 32 locations in 26 Indonesian provinces from September 2012 to August 2014. The WHOij2 strain obtained from Danish Pest Infestation Laboratory University of Aarhus Denmark in 2011 was used as the control (susceptible strain). All strains were reared in Entomology Laboratory, School of Life Sciences and Technology, Institut Teknologi Bandung, Bandung, Indonesia. Methods used to rear larvae and adults were those described by Keiding and Arevad (1964) with modification. House flies were reared in a stainless steel frame cage measuring 30×30×30 cm covered with tulle fabric and sleeve openings at the front. Dry food whole-milk powder mixed with refined sugar and dried yeast, with the ratio 100:100:2 was given ad libitum in petri dish. Small jar was filled with tap-water was provided for water-supply. A cup with rolls of tissue moistened in 5% milk was provided for eggs laying. After three days, the hatched larvae were transferred to larval rearing media which consist of 100 g bran, 50 g broiler chicken feed and 150 mL tap water mixture well in 1 L jar and covered with 3 cm rice husks. Pupae formed in seven days were sorted and placed in a new cage. Five-seven day old house fly adults emerged were used in the bioassay and provided refined sugar in petri dish and water on cotton ad libitum before the test. Male and female adult flies were separated in difference cage. Briefly, they were kept at 24-27°C and 50-75% RH before used.
Insecticides: The insecticides used were 94% permethrin (MIN, from PT. Triman Sentosa Jakarta), 95% imidacloprid (MIN TECH) and piperonil butoxide (PBO) diluted in analytical-grade acetone.
Insecticides bioassays: The bioassay procedure developed by Kristensen and Jespersen (2004, 2008) was followed for determining resistance in house flies. Topical application bioassay was carried out on female house flies and male house flies with feeding bioassay.
A total 30 of chilled house flies were treated with 1 μL of permethrin in acetone applied topically on the dorsal thorax using a micropipette. Five-Thirteen concentrations of permethrin ranging from 1.22×10–6 to 5×10–3% in acetone were used for estimating the LD50 and LD95. After application house flies were placed in 14 oz plastic jar, covered and secured with a tulle cloth. An absorbent cotton ball with 15% sucrose placed on the screened cloth as food source. Control groups were treated only with acetone. The numbers of house flies dead or paralyzed were recorded at 24 h after treatment. In the same method, the piperonyl butoxide (PBO) as a synergist was used to view the activity of detoxifying enzymes system of Mixed Function Oxidase (MFO) by adding to permethrin in ratio 1:5 for piperonyl butoxide: permethrin.
Susceptibility to imidacloprid was determined using a non-choice feeding bioassay with male house flies. Insecticide-treated refined sugars were prepared 24 h before treatment. Ten milliliter of imidacloprid dissolved in a mount of acetone with concentration ranging from 1.25×104 to 0.128% was added to 20 g refined sugar and stirred until the acetone evaporated. A total of 0.5 g of imidacloprid-treated refined sugar in small aluminum dishes as the only food and moistened cotton ball were placed in the bottom of 350 cm3 transparent plastic pot. Three replicates of 20 group of house flies were tested against 5-14 concentrations to determining the LC50 and LC95. Acetone-treated refined sugar was similarly provided to the control groups. Dead and ataxic house flies were recorded 72 h after treatment.
Statistical analysis: The lethal doses (LD50 and LD95) or lethal concentrations (LC50 and LC95) were calculated by means of probit analysis (POLO-PC, LeORA Software, 2004).
Resistance ratio: The Resistance Ratio (RR) was determined by comparing the LD50 or LC50 of field-collected populations to the LD50 or LC50 of susceptible strain. The RR was not considered significantly different if the 95% confidence interval overlapped with 1 (Robertson et al., 2007). The determination of resistance levels were based on the resistance classification of the house flies developed by WHO (1980), i.e., low (<10), moderate (10-40), high (40-160) and very high (>160).
RESULTS
The level of susceptibility of the widely used insecticides permethrin and imidacloprid were determined for 32 field-collected house flies strains from 26 provinces throughout Indonesia archipelago (Fig. 1) within two years, 2013 and 2014.
Toxicity of permethrin to different strains of house flies: According to the resistance classification of the house flies developed by WHO (1980), overall results of the assays to permethrin (Table 1 and 2) showed that the majority of field collected strains had very high level of resistance to permethrin, i.e., 25/32 (78.125%) with RR50 >160.
 | |
| Fig. 1: | Sample locations distribution of the M. domestica L. strains in Indonesia |
| Table 1: | Toxicity of permethrin to adult female house flies collected in 2013 from refuse sites in Indonesia |
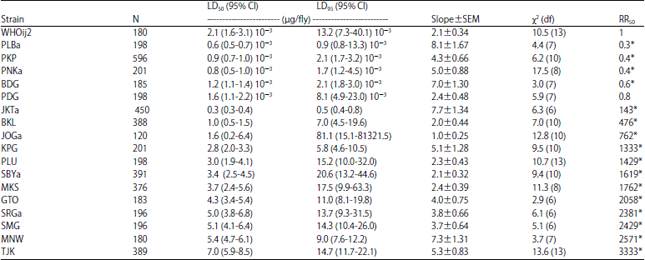 | |
| N: Number of house flies used in each bioassay, χ2: Value is calculated by probit analysis using POLO PC software, Resistance Ratio (RR): LD50 of field collected population/LD50 of susceptible strain, *Significantly different from 1.0 when 95% CL did not overlap | |
| Table 2: | Toxicity of permethrin to adult female house flies collected in 2014 from refuse sites in Indonesia |
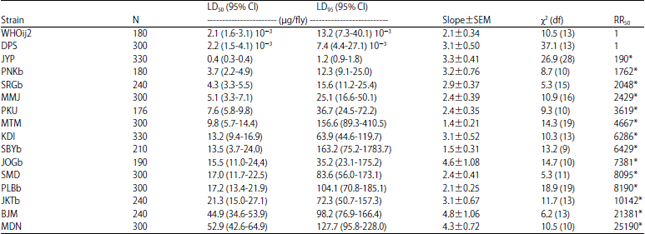 | |
| N: Number of house flies used in each bioassay, χ2: Value is calculated by probit analysis using POLO PC software, Resistance Ratio (RR): LD50 of field collected population/LD50 of susceptible strain, *Significantly different from 1.0 when 95% CL did not overlap | |
The slope of the probit line of three field strains (JOGa, MTM and SBYb) were <2.0, indicating the heterogeneity response of these strains to permethrin. In addition, the level of resistance of populations collected within 2014 was higher than strains collected in 2013. The addition of PBO in assay using permethrin (1:5) in 12 of 14 selected strains, tended to reduce the values of LD50 24 h as to those without the addition of PBO. Six strains, i.e., SRGb, MTM, KDI, PLB and SBYb, were greatlyreduced the LD50 value in the presence of PBO (Fig. 2).
Toxicity of imidacloprid to different strains of house flies: In contrast with the results of the permethrin bioassay, all of the field-collected strains of M. domestica (32 strains) in this study showed no to low levels of resistance to imidacloprid (0.4-6.1 fold) (Table 3 and 4). Fourteen strains had no resistance levels according to the criteria by Abbas et al. (2015a) with RR less than two when compared to the susceptible standard strain. The shallow slopes of the dose-response curve of three field populations (DPS, SBYb and PNKb) indicating the heterogeneity response of these populations to imidacloprid. The LC50 value or resistance ratio of the field strains collected in 2014 (Table 4) was with in the range of responses tested in 2013 (Table 3).
| Table 3: | Toxicity of imidacloprid to adult male house flies collected in 2013 from refuse sites in Indonesia |
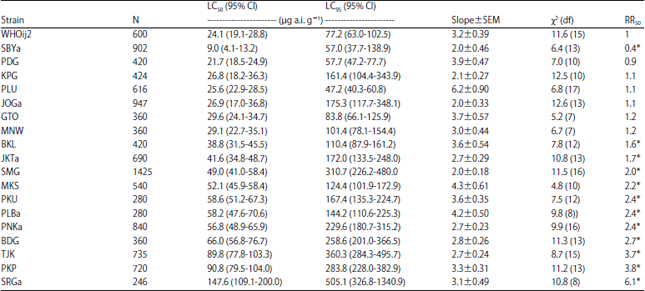 | |
| N: Number of house flies used in each bioassay, χ2: Value is calculated by probit analysis using POLO PC software, Resistance Ratio (RR): LC50 of field collected population/LC50 of susceptible strain, *Significantly different from 1.0 when 95% CL did not overlap | |
| Table 4: | Toxicity of imidacloprid to adult male house flies collected in 2014 from refuse sites in Indonesia |
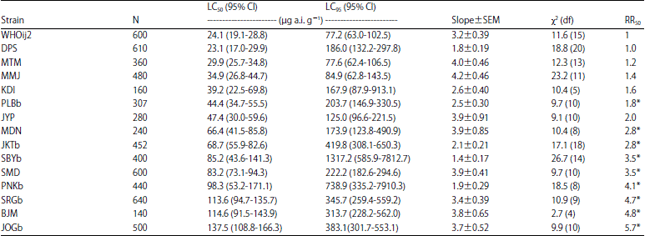 | |
| N: Number of house flies used in each bioassay, χ2: Value is calculated by probit analysis using POLO PC software, Resistance Ratio (RR): LC50 of field collected population/LC50 of susceptible strain, *Significantly different from 1.0 when 95% CL did not overlap | |
DISCUSSION
The findings that highly resistance to permethrin can be partially explained by the wide use of permethrin and other pyrethroids for house fly control in urban, agricultural and veterinary environments since early 1980s. In addition, the high levels of resistance might have been related to the cross resistance to previously used, now banned, insecticides such as DDT and dieldrin, since organochlorines and pyrethroids have similar mode of action. The results of the present study appears similar with the previous study conducted in our laboratory which showed that Aedes aegypti and Blattella germanica resistance to permethrin. For example A. aegypti from Bandung had a high resistance (79.3 fold) (Ahmad et al., 2007), whilst for Blatella germanica, the study revealed the existence of extremely high resistance to permethrin (RR50 1013.17 fold) (Rahayu et al., 2012).
This study also showed that the addition of PBO together with the application of permethrin, reduced the LD values for 12 strains. There was possibility that the PBO which works by inhibiting Mixed Function Oxidase (MFO) play important role in the occurrence of resistance.
 | |
| Fig. 2: | The Toxicity of permethrin and permethrin+PBO in 14 strains of house fly M. domestica L from Indonesia |
Nonetheless, our findings could not rule out, some other mechanisms that maybe responsible for the observed very high resistance to permethrin in majority of the strains, such as cuticular reduction penetration (pen) mechanism which was reported on multi resistant strains LPR which had 5900 times the level of resistance to permethrin (Shono et al., 2002). Moreover, the presence of kdr and super kdr mutation in the segment gen of voltage-sensitive sodiun channel (Vssc) has been known responsible to permethrin resistance in many regions (Huang et al., 2004; Rinkevich et al., 2006; Taskin et al., 2011; Al-Deeb, 2014; Mazzoni et al., 2015).
In contrast with our findings, Bong and Zairi (2010) reported that in the State of Penang Malaysia, permethrin was still the most effective insecticide in controlling M. domestica with resistance ratios ranging from 0.5-2.4 fold. Abbas et al. (2015b) in Pakistan reported resistance ratios for pyrethroids of 14-55 fold for cypermethrin, 11-45 fold for bifenthrin and 0.84-4.06 fold for deltamethrin. The high level of resistance to permethrin was also found by Kaufman et al. (2010a) in all field-collected strain from Florida. However, our findings also showed that about 5/32 (15.6%) of the strains demonstrated greater levels of susceptibility to permethrin than the susceptible strain (RR50<1). This finding was unexpected since it is unlikely that the flies in the areas surveyed had not been exposed to permethrin. Nonetheless, since we do not have historical records of permethrin or other pyrethroids used in those five areas, the question remains to be unanswered.
Therefore the findings from the present study which suggest that very high resistance on M. domestica to permethrin is widespread in Indonesia. It is rather alarming and need preventive actions, as it is suggested before that to avoid the possible control failure in the field, a permethrin halt for a certain period should be carried out (Rahayu et al., 2012). However, the fact that house flies in this study have developed resistance to permethrin, we have to admit that no study has been carried out to measure the operation impact of resistance. So we were not sure whether the very high resistance levels found in this study has led to any control failure or not. As confirmed by Marcon et al. (2003) that insecticide resistance was more due to the results of migration of resistant house flies rather than the exposure of insecticide. House flies were more intensely exposed to chemical treatment in domestic settings and resistant individuals may move to other site.
Imidacloprid was a new insecticide registered in Indonesia in the late 2000s and has not been widely used. Commonly, the use of imidacloprid in commercially insecticides was restricted in the poultry area, meanwhile the sampling locations of this present study were garbage dump in urban area. Nonetheless, the results showed that resistance to imidacloprid was emerged in the house fly in Indonesia.
Kaufman et al. (2006) also reported low levels of resistance to imidacloprid in the USA. Although imidacloprid had not been used previously, six colonies of house flies collected in 2004 exhibited limited cross resistance to imidacloprid (RR 3.1-8.0 fold). Recent report in Pakistan by Abbas et al. (2015b) also showed no to low level of resistance to imidacloprid with RR 2-14 fold. Inspite of resistance ratio remain relatively low, Kaufman et al. (2010a) noticed that it was significantly resistant to imidacloprid. Furthermore, Kaufman et al. (2010b) concluded that house flies were capable of developing high levels of resistance to imidacloprid when presented as sugar bait. The rapidity imidacloprid resistance can develop under high selection pressure and the early appearance of that resistance in field situations.
Regarding to the condition of low level of resistance, Gerry and Zhang (2009) supposed that house flies can develop resistance to imidacloprid baits through altered behavioral response to imidacloprid. House fly was able to elude against food containing imidacloprid in around. Moreover, imidacloprid may also have acted as a feeding deterrent in resistant house fly.
In Denmark, the situation was rather alarming and perplexing due to the fact that even though neonicotinoid insecticides had not been used in the area of collections, Markussen and Kristensen (2010) observed that two field populations demonstrated RR 20 and 140 fold. Even though there was no clear explanation about the findings, they suggested that the high resistance to imidacloprid was partly due to altered cytochrome P450 activity.
CONCLUSION
Results of this study, which is the first report of insecticides resistance status covering a wide range of field-collected populations of house fly in Indonesia, provide important baseline data on the susceptibility and resistance of field population of house fly to permethrin and imidacloprid in Indonesia. Thus, the findings have reminded us about the importance of current knowledge about the extent and geographic distribution of insecticide resistance. Monitoring of resistance to commonly used insecticides indicates a need to identify resistant mechanisms to identify alternative modes of house fly control.
ACKNOWLEDGMENTS
This work was funded the by Directorate General of Higher Education Ministry of Education Indonesia. The authors thank supporting personnel at Universitas Tanjungpura, Local Government of Kalimantan Barat, Biaya Operasioanl Perguruan Tinggi Negeri-Institut Teknologi Bandung (BOPTN-ITB) and Sekolah Ilmu dan Teknologi Hayati-ITB, Bandung for funding support. The authors also thank to the volunteers for assisting in house fly sampling and PT Triman Sentosatama, Jakarta, Indonesia for providing insecticides used in this study. The authors especially thank Dr. Michael Kristensen (Department of Agroecology, Aarhus University, Denmark) for valuable discussion, suggestions and encouragement.
REFERENCES
- Abbas, N., S.S. Shad and M. Ismail, 2015. Resistance to conventional and new insecticides in house flies (Diptera: Muscidae) from poultry facilities in Punjab, Pakistan. J. Econ. Entomol., (In Press).
CrossRefDirect Link - Abbas, N., S.A. Shad and R.M. Shah, 2015. Resistance status of Musca domestica L. populations to neonicotinoids and insect growth regulators in Pakistan poultry facilities. Pak. J. Zool., 47: 1663-1671.
Direct Link - Ahmad, I., S. Astari and M. Tan, 2007. Resistance of Aedes aegypti (Diptera: Culicidae) in 2006 to pyrethroid insecticides in Indonesia and its association with oxidase and esterase levels. Pak. J. Biol. Sci., 10: 3688-3692.
CrossRefPubMedDirect Link - Al-Deeb, M.A., 2014. Pyrethroid insecticide resistance kdr gene in the house fly, Musca domestica (Diptera: Muscidae), in the United Arab Emirates. Agric. Sci., 15: 1522-1526.
CrossRefDirect Link - Astari, S. and I. Ahmad, 2005. Insecticide resistance and effect of piperonyl butoxide as a synergist in three strains of Aedes aegypti (Linn.) (Diptera: Culicidae) against insecticides permethrin, cypermethrin and D-allethri. Buletin Penelitian Kesehatan, 33: 73-79.
Direct Link - Akiner, M.M. and S.S. Caglar, 2006. The status and seasonal changes of organophosphate and pyrethroid resistance in Turkish populations of the house fly, Musca domestica L. (Diptera: Muscidae). J. Vector Ecol., 31: 426-432.
CrossRefPubMedDirect Link - Bong, L.J. and J. Zairi, 2010. Temporal fluctuations of insecticides resistance in Musca domestica Linn (Diptera: Muscidae) in Malaysia. Trop. Biomed., 27: 317-325.
Direct Link - Cao, X.M., F.L. Song, T.Y. Zhao, Y.D. Dong, C.X. Sun and B.L. Lu, 2006. Survey of deltamethrin resistance in house flies (Musca domestica) from urban garbage dumps in Northern China. Environ. Entomol., 35: 1-9.
CrossRefDirect Link - Huang, J., M. Kristensen, C.L. Qiao and J.B. Jespersen, 2004. Frequency of kdr gene in house fly field populations: Correlation of pyrethroid resistance and kdr frequency. J. Econ. Entomol., 97: 1036-1041.
CrossRefDirect Link - Kaufman, P.E., A.C. Gerry, D.A. Rutz and J.G. Scott, 2006. Monitoring susceptibility of house flies (Musca domestica L.) in the United States to imidacloprid. J. Agric. Urban Entomol., 23: 195-200.
Direct Link - Kaufman, P.E., S.C. Nunez, C.J. Geden and M.E. Scharf, 2010. Selection for resistance to imidacloprid in the house fly (Diptera: Muscidae). J. Econ. Entomol., 103: 1937-1942.
CrossRefDirect Link - Kaufman, P.E., S.C. Nunez, R.S. Mann, C.J. Geden and M.E. Scharf, 2010. Nicotinoid and pyrethroid insecticide resistance in houseflies (Diptera: Muscidae) collected from Florida dairies. Pest Manage. Sci., 66: 290-294.
CrossRefDirect Link - Keiding, J. and K. Arevad, 1964. Procedure and equipment for rearing a large number of housefly strains. Bull. World Health Organiz., 31: 527-528.
PubMedDirect Link - Kristensen, M. and J.B. Jespersen, 2004. Susceptibility of spinosad in Musca domestica (Diptera: Muscidae) field populations. J. Econ. Entomol., 97: 1042-1048.
CrossRefDirect Link - Kristensen, M. and J.B. Jespersen, 2008. Susceptibility to thiamethoxam of Musca domestica from Danish livestock farms. Pest Manage. Sci., 64: 126-132.
CrossRefDirect Link - Markussen, M.D.K. and M. Kristensen, 2010. Cytochrome P450 monooxygenase-mediated neonicotinoid resistance in the house fly Musca domestica L. Pestic. Biochem. Physiol., 98: 50-58.
CrossRefDirect Link - Marcon, P.C.R.G., G.D. Thomas, B.D. Siegfried, J.B. Campbell and S.R. Skoda, 2003. Resistance status of house flies (Diptera: Muscidae) from Southeastern Nebraska beef cattle feedlots to selected insecticides. J. Econ. Entomol., 96: 1016-1020.
CrossRefDirect Link - Mazzoni, E., O. Chiesa, V. Puggioni, M. Panini, G.C. Manicardi and D. Bizzaro, 2015. Presence of kdr and s-kdr resistance in Musca domestica populations collected in Piacenza province (Northern Italy). Bull. Insectol., 68: 65-72.
Direct Link - Memmi, B.K., 2010. Mortality and knockdown effects of imidacloprid and methomyl in house fly (Musca domestica L., Diptera: Muscidae) populations. J. Vector Ecol., 35: 144-148.
CrossRefDirect Link - Rahayu, R., I. Ahmad, E.S. Ratna, M.I. Tan and N. Hariani, 2012. Present status of carbamate, pyrethroid and phenylpyrazole insecticide resistance to German cockroach, Blattella germanica (Dictyoptera: Blattellidae) in Indonesia. J. Entomol., 9: 361-367.
CrossRefDirect Link - Rinkevich, F.D., L. Zhang, R.L. Hamm, S.G. Brady, B.P. Lazzaro and J.G. Scott, 2006. Frequencies of the pyrethroid resistance alleles of Vssc1 and CYP6D1 in house flies from the Eastern United States. Insect Mol. Biol., 15: 157-167.
CrossRefDirect Link - Robertson, J.L., R.M. Russell, H.K. Preisler and N.E. Savin, 2007. Bioassay with Arthropods. 2nd Edn., CRC Press, New York, USA., ISBN-13: 9781420004045, Pages: 224.
Direct Link - Scott, J.G., C.A. Leichter, F.D. Rinkevihc, S.A. Harris and C. Su et al., 2013. Insecticide resistance in house flies from the United States: Resistance levels and frequency of pyrethroid resistance alleles. Pestic. Biochem. Physiol., 107: 377-384.
CrossRefDirect Link - Scott, J.G., W.C. Warren, L.W. Beukeboom, D. Bopp and A.G. Clark et al., 2014. Genome of the house fly, Musca domestica L., a global vector of diseases with adaptations to a septic environment. Genome Biol., Vol. 15.
CrossRefDirect Link - Shono, T., S. Kasai, E. Kamiya, Y. Kono and J.G. Scott, 2002. Genetics and mechanisms of permethrin resistance in the YPER strain of house fly. Pestic. Biochem. Physiol., 73: 27-36.
CrossRefDirect Link - Taskin, V., S. Baskurt, E. Dogac and B.G. Taskin, 2011. Frequencies of pyrethroid resistance-associated mutations of Vssc1 and CYP6D1 in field populations of Musca domestica L. in Turkey. J. Vector Ecol., 36: 239-247.
CrossRefDirect Link