
Safaa M. Halawa
Department of Plant Protection, Faculty of Agriculture, Behan University, Moshtohor, Egypt
Reinhold Hustert
JFB-Institut of Zoologie, Abt. Neurobiologie, Georg-August-Universitat Gottingen, Germany







ABSTRACT
Essential oil from lovage (Levisticum officinale Koch.) is a potential low-risk insecticide containing monoterpenes. It was tested with increasing concentrations in saline for neural effects on the autonomous ventilatory rhythm of an isolated abdominal ganglion (AG5) which supplies the fifth abdominal segment of Locusta migratoria (Linneus). The inhibition by lovage oil was compared with that of its main components, the enantiomers of limonene (D- and L-limonene). All substances depressed the persistent ventilatory motor output: the concentration-dependent reduction of the neural activity was measured with the average Action Potential (AP) frequency of the inspiratory motoneurons in the median nerve. Occasionally the motor output could switch from bursting activity with more or less gradual transients to continuous tonic firing. Recovery of the AG5 in pure saline after highest concentrations had been applied was gradual but the ventilatory output never returned completely to the pre-application level of motor activity. The strong short-term neurotoxic effects of the limonene enantiomeres in the insect CNS are followed by a moderate long term neural depression.
PDF Abstract XML References Citation
Received: September 13, 2013;
Accepted: January 24, 2014;
Published: March 08, 2014
How to cite this article
Safaa M. Halawa and Reinhold Hustert, 2014. Neural Activity of a Locust Ganglion is Impaired by Limonene and Lovage
(Levisticum officinale) Extracts. Journal of Entomology, 11: 1-13.
DOI: 10.3923/je.2014.1.13
URL: https://scialert.net/abstract/?doi=je.2014.1.13
DOI: 10.3923/je.2014.1.13
URL: https://scialert.net/abstract/?doi=je.2014.1.13
INTRODUCTION
Many plants are protected from being eaten by their chemical components: feeding deterrents repel while noxious compounds may not prevent ingestion of a plant but later they influence the vitality of the consumer. They inflict damage on one or several subsystems of the organism which can weaken the animal, cause lethal effects or reduce its vitality and fertility. Many studies of essential oil and plant extract compounds from specific plants showed toxic, deterrent activity or repellent effects on a range of insect species (Regnault-Roger, 1997; Prabuseenivasan et al., 2006; Akinwumi et al., 2007; Ikbal et al., 2007; Koul et al., 2008; Soummane et al., 2011; Pandey et al., 2012; El-Gendy and Shaalan, 2012). The typical structure-activity studies in individual insect species provided only limited insights into the mode of action of these compounds. Treatments of flies with essential oils containing monoterpenes or the application of purified constituents resulted in visible symptoms (Ryan and Byrne, 1988), apparently from neurotoxic influences. The overt symptoms were hyperactivity, convulsions and tremors followed by paralysis (‘knock-down’)-similar to those produced by such insecticides as organophosphates and carbamates (Grundy and Still, 1985; Coats et al., 1991; Kostyukovsky et al., 2002).
Many of the deterrent plants belong to the family Apiaceae. They have received much attention at least partly owing to the presence of different kind of terpenes. It was found that the water extracts of fennel (Foeniculum capillaceum Gilib.), garden angelica (Archangelica pfficinalis Hoffm.), common caraway (Carum cavi L.), garden lovage (Levisticum officinale Koch.) and coriander (Coyiandrum sativum L.) significantly affected the feeding effectiveness as well as developmental potential of the Colorado potato beetle (Leptinotarsa decemlineata Say) (Wawrzyniak and Lamparski, 2006).
The seeds of the lovage plant (L. officinale Koch.) are commonly used as a spice. The yield of oil (w/w%) in different stages was in the order: immature fruit (1.5%) > green mature fruit (1.0%) > ripened fruit (0.6%) > flower (0.1%) (Mirjalili et al., 2010) deterrent and insecticidal effects of essential oil of lovage, against Locusts Locusta migratoria (Linneus). The GC-MS analysis of lovage essential oil revealed that the major constituents in oil was limonene followed by beta pinene, pentylbenzene, beta myrcene, ocimene and the hydrocarbons were also detected in high quantity. Limonene was the most abundant headspace component in lovage constituents and revealed a mixture of its enantiomers D and L-limonene. Enantiomers differ from each other only in stereo-configuration but their odors can be perceived as different by insects and mammals (Friedman and Miller, 1971; Leitereg et al., 1971; Heth et al., 1992; Taniguchi et al., 1992; Laska and Teubner, 1999a, b; Laska et al., 1999a, b; Laska and Galizia, 2001; Rubin and Katz, 2001). The higher concentration of lovage oil induced the lowest eaten area percentage (44.22%) of L. migratoria, followed by the D and L-limonene (18.89 and 18.79%), respectively. Antifeedant and protection activity percentage were increased by increasing the concentration. The highest mortality percentage (100%) of L. migratoria nymphs was recorded at 5000 ppm lovage oil while it was 93.1, 86.7 and 82.8% in the case of D-limonene, L-limonene and the mix of D and L-limonene, respectively. Ngoh et al. (1998) stated that Limonene had previously been considered as a potential pesticide for insects since benzene derivatives such as limonene were more toxic and repellent than other terpenes to Periplaneta americana. Ozaki et al. (2003) stated that D-limonene has strong oral toxicity in flies.
Major targets of many toxins, after their uptake in the body, are located in the Central Nervous System (CNS) and the peripheral nervous system (Ngoh et al., 1998) stated that the exact mode of action of the four contact toxicants (eugenol, methyl eugenol, safrole and isosafrole) on P. americana is unknown, the contact toxicity of these chemicals suggest that they can somehow penetrate the integument of the insects, to act presumably on the nervous or respiratory system to produce the killing effect. Ferber et al. (2001) reported that the different toxins can act at various levels of neuronal metabolism: on cell membranes and their ion channels or on synaptic mechanisms of the pathways of neural transmission, such as many pesticides do in insects. The effects can be over-excitation or depression of neural activity that is seen as abnormal motor output of the nervous system.
In larger insects such as locusts, specific physiological preparations or isolated parts of the CNS can be used to study the effects of neuro-active substances if they are super fused by different concentrations of plant extracts or fractions that may be noxious. We used a new preparation from the insect CNS which is very simple to prepare for testing neurotoxic effects: the isolated fifth abdominal ganglion (AG5) of adult specimens of Locusta migratoria. When this ganglion is separated from the neighboring ganglia (AG4 and AG6) by cutting the connectives, it releases a continuing ventilatory motor output in axons of peripheral nerves (Hustert and Mashaly, 2013). The isolated AG5 maintains this autonomous ventilatory motor output for hours if kept in physiological saline with sufficient oxygen supply via the “bath-just…” as shown previously for the more complex locust metathoracic ganglion (Bustami and Hustert, 2000).
In the current study we investigated the acute toxicities and effects of essential oil from lovage (Levisticum officinale Koch.) as insecticide containing monoterpenes. It was tested with increasing concentrations in saline for neural effects on the autonomous ventilatory rhythm of an isolated abdominal ganglion (AG5) which supplies the fifth abdominal segment of Locusta migratoria (Forskal). The inhibition by lovage oil was compared with that of its main components, the enantiomers of limonene (D-and L-limonene).
MATERIALS AND METHODS
Plant material: Seeds of lovage plants (Levisticum officinale Koch.) were harvested in an experimental farm (Department of Horticulture, in the Faculty of Agriculture, Moshtohor, Benha University, Egypt.
Extraction of the essential oil: Lovage essential oil was obtained by hydro-distillation with a Clevenger apparatus (Clevenger, 1928) for 2-3 h. First, 150 g of lovage seeds were powdered coarsely and then hydro-distilled on a sand bath. The collected oil was dried over anhydrous sodium sulfate, filtered, stored and later diluted for bioassays. The identification and quantitative determination of volatile components was performed by GC/MS analysis.
Experimental animals: A colony of gregarious Locusta migratoria (Forskal) was kept in the Institute of Zoology of the University of Göttingen (Germany), reared under crowded conditions and fed on wheat. Lighting had a 12:12 h light-dark cycle and temperature was kept at 32±2°C and 65±5% R.H.
Preparation of the test ganglion and nerve recording: Before isolating the abdominal ganglion (AG5) (Fig. 1) adult males of Locusta migratoria were chilled below 5°C for anesthesia. The head and the rear segments of the abdomen were cut off. The gut was removed and the dorsal part of the insect was cut away with lateral incisions. The ventral part containing the nerve cord was fixed with insect needles in a small petri-dish on Sylgard (Dow Corning) and flooded with saline. The second free abdominal ganglion (AG5) was carefully dissected out by cutting its connectives to the adjacent ganglion and its peripheral nerves at some distance from the ganglion. That severed also the tracheal supply entering at the ventral nerve2 (N2) but the ganglion was kept in a shallow bath of locust saline (pH 6.8, with sucrose (Clements and May, 1974) providing a sufficient oxygen supply.
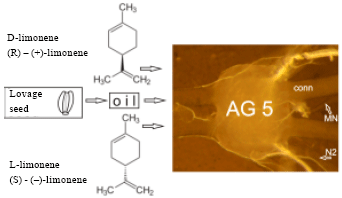 | |
| Fig. 1: | An isolated locust abdominal ganglion provides a simple model for testing neurotoxicity. The regular ventilatory motor output activity is the monitor. Extracts from lovage and its major components D- and L-limonene (enantiomers) are neurotoxic. A short-term suppression and a long-term toxicity can be distinguished |
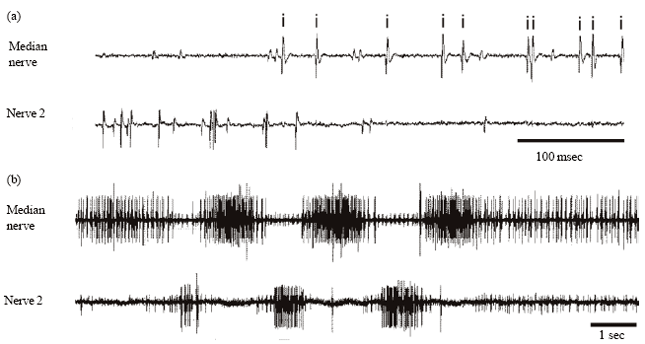 | |
| Fig. 2(a-b): | Efferent patterns of the isolated fifth abdominal ganglion. Recordings from the median nerve and from one ventral nerve (nerve 2) of the isolated fifth abdominal ganglion (at room temperature of 23°C). (a) There are four motor units recorded in the median nerve (upper trace): two larger units with similar amplitude (marked with “i”) innervate the inspiratory muscles (M225) and two small units which innervate spiracular muscles in the sixth abdominal segment. In nerve 2 (lower trace) various slow and fast motor units supply dorso-ventral expiratory muscles of the fifth abdominal segment. (b) Simultaneous recording of tonic activity and alternating bursts in the median nerve and nerve 2 during a period of discontinuous ventilation |
The typical output of the motoneurons for the ventilatory muscles in this segment alternates between inspiration and expiration (Lewis et al., 1973; Hustert, 1975). That rhythm was recorded routinely (monopolar suction electrodes with tapering tips drawn from thin plastic tubes) from the stumps of the paired ventral nerves 2 (N2; Fig. 2b) and from the unpaired posterior median nerve (MN; Fig. 2a). The main indicator of ventilation was the activity of the two large motoneurons with very similar amplitudes and frequency pattern in the posterior median nerve. These supply the pair of inspiratory muscles M225 (Snodgrass, 1935) of the next (6th abdominal) segment (Miller, 1960). In the same (median) nerve the only other two motoneurones (with very small amplitudes) supply the spiracle closer muscles in the 6th abdominal segment. Their irregular activity only rarely joins in the ventilatory rhythm.
All action potentials were pre-amplified (1000x) and the larger units selected after they had been recorded on magnetic tape or by digital computer recording (Datapac 2K2, Run Technologies). The average frequency of their firing rates (combined from both units) was recorded (1 min standard recording time). That inspiratory activity monitored the neural effects of substances applied at different concentrations on the locust ganglion. The inspiratory motor output of the ganglion displayed occasional changes from bursting to more tonic firing. Therefore, burst intervals were neglected and only the average frequencies over the one minute periods were taken for the analysis.
The saline in the bath could be replaced routinely and substances were added to saline to test their effect on the motor output of the ganglion.
Bath applications: For testing the effects of potentially neurotoxic substances contained in lovage (L. officinale Koch.), the essential oil was used or the pure allochemicals the enantiomers of limonene (D- and L-limonene, obtained from Sigma-Aldrich). These were diluted in locust saline (Clements and May, 1974) with the aid of small amounts of detergent (Tween 20 at 10 μg L-1 in the stock concentration) and applied on the isolated ganglion in the bath (six concentrations: 0.25, 0.5, 1.25, 2.5, 5 and 10 μg L-1) for the different test substances: (1) Lovage essential oil, (2) Saline with L-limonene, (3) Saline with D-limonene, (4) Saline with a 50% of D and L-limonene mixture and (5) As a control saline and detergent only at concentrations used for the extracts and substances.
The basic protocol was to record neuronal activity for at least 5 minutes (recording for analysis during the last minute), then to change the bathing fluid with the next and higher concentration of substance in saline. After a recovery time for five minutes, the effects of the new concentration were recorded. After each session of recording from a ganglion, going through all (six) different concentrations for one tested substance, the ganglion was washed in pure saline three times.
Statistical analysis: The inspiratory motoneuron activity (large spikes) was analyzed by counting the total number of spikes occurring within a 60 sec window for every concentration. From the spike intervals (t) the instantaneous frequencies (t-1) were taken and sorted in 10 Hz intervals from 0 to 300 Hz in order to establish a frequency distribution for each record. For the sake of comparability, a normal distribution was assumed and then the standard deviation of the frequency distribution and the mean frequency were calculated.
Data from concentration-dependent responses (spikes in Hz) of the tested substances were transformed to square roots (Square root values were calculated (N sqrt = N/N max) and subjected to analysis using Wilcoxon Rank-comparison (Wilcoxon, 1945) of lower to next higher within each previous concentration. The values of the total number of spikes (N) the standard deviation (SD) and mean frequency (Mean F) are used to characterize the breathing activity during each record. Also, analysis of variance and “F” tests were used to compare between substances and using a computer software package, “costat” a product of Cohort software Inc., Berkeley. California. USA. Duncan's multiple range test (Duncan, 1955) was applied subsequently to differentiate between means.
The Square root characteristic values of each concentration were averaged between the five ganglia. Means of results for all substances were graphically displayed under “overview”, of the data for each individual ganglion under the respective substance category.
RESULTS
Effects of the lovage essential oil on the ventilatory output of the isolated AG5 of locusts: The application of different concentrations of the lovage essential oil (L. officinale Koch.) in saline changed the activity regularly. It induced strong depression effects (p<0.02) on the ganglionic motor output (Fig. 3).
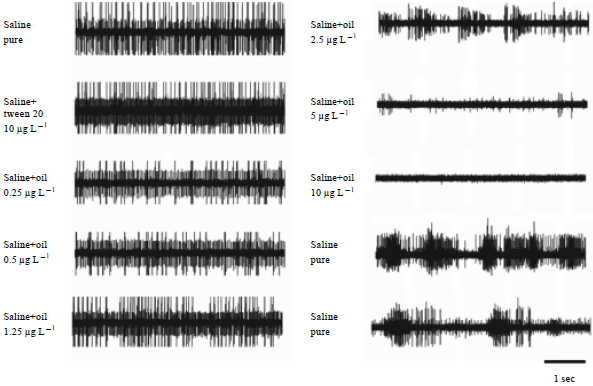 | |
| Fig. 3(a-b): | Depression of neuronal activity in the isolated 5th abdominal ganglion by the potentially neurotoxic lovage oil. Each trace is a sample of spiking activity of the large inspiratory motor units during exposure to different oil concentrations. The sequence is from pure saline (track 1) over saline with just the detergent Tween 20 (track 2) to ascending oil in concentrations (tracks 3 to 8) and finally bathing in pure saline again (tracks 9/10) |
The mean frequency of spikes fell to 44 to around 35 Hz (spikes sec-1) at 0.25 μg of oil concentration (Fig. 4a, e). Higher concentrations resulted not only in significant decreases of the mean frequency of spikes but also in decrease of spike amplitudes (Fig. 3). At the maximum oil concentration (10 μg L-1) the motoneurones of the isolated ganglion discharged at an average of 6 Hz (p<0.05) or less (Fig. 4a). Increasing the concentrations always resulted in step by step reduction of the ventilatory motor output as measured by the average AP frequency of the inspiratory motoneurons in the median nerve (Fig. 4a, e). In some preparations the motor output also switched from bursting activity to continuous tonic firing. Washing the ganglion with pure saline after the highest concentration had been applied resulted in gradual recovery of the average output from the ganglion (14.1 Hz after 5 min, 19.2 Hz after 30 min and 22.4 Hz after 1 h); (Fig. 4a, e). The spiking activity of the motor units never reached the pre-application level of excitation of 44.0 Hz. Control experiments with concentrations of only the detergent in saline (Tween 20 at 10 μg L-1), which had been necessary to dilute the oil in saline, showed no significant changes (39.1 Hz in saline+tween20 vs. 44.0 Hz in saline only) of the motor output from the ganglion (Fig. 3).
Effects of the pure enantiomers D- and L-limonene on the ventilatory output of the isolated (AG5) of locusts: Limonene, a monoterpenoid, is the major component of the allochemicals in lovage seed (L. officinale Koch.) essential oil, as identified by GC/MS analysis.
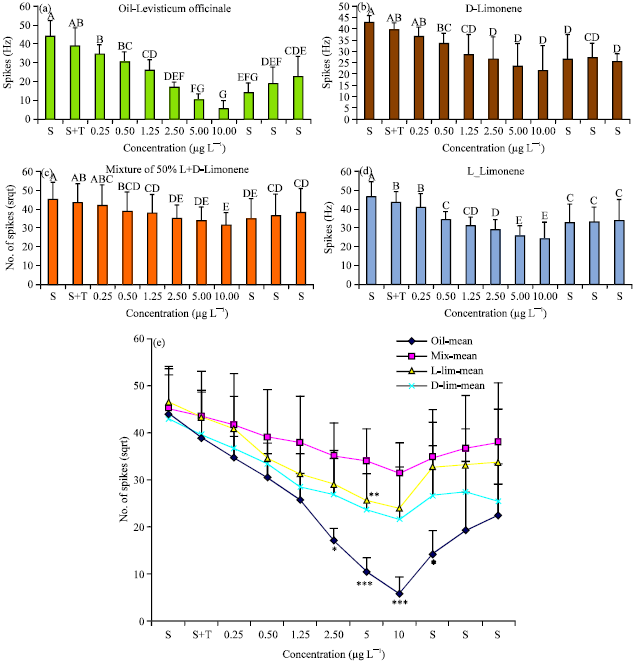 | |
| Fig. 4(a-e): | Concentration-dependent decrease of inspiratory spike frequency. Motor output from the isolated fifth abdominal ganglion when treated with different concentrations (each experiment: n = 5) of extracts, (a) Essential oil of lovage, (b) Pure D-limonene, (c) Mixture of 50% D- and L-limonene, (d) Pure L-limonene. For A-D: Results of Wilcoxon-comparison (comparison of lower to next higher within each concentration) as tests of difference between the adjacent results in each series of tests. The letters above the standard deviation indicators are different when they are significantly different between neighboring columns and (e) Comparative graph of the effect of different substances on the inspiratory motor output. Differences between the effects of the substances were tested statistically at each concentration value for the output under saline and the different concentration in saline. concentration in saline. S: Saline only, S+T: Saline with the detergent tween, numerals: Concentration of the respective substances |
It is a mix of the limonene enantiomers D- and L-limonene. The effects of pure D-limonene and L-limonene alone and the effect of their mixture (50% of D- and L-limonene) were recorded from the Median Nerve (MN) of the ganglion at different bath concentrations (0.25, 0.5, 1.25, 2.5, 5 and 10 μg L-1) and after subsequent washing out with pure saline following the highest concentrations (Fig. 4b-e).
When D- and L-limonene were applied separately each produced strong depression effects on the ganglion motor output but D-limonene was more effective than L-limonene (p<0.05). Surprisingly, mixtures of 50% D- and L-limonene with the same final concentrations were less effective than application of the single enantiomers (Fig. 4b-e). All applied chemicals caused at increased concentrations in saline a significant decrease in the ganglion motor output (p<0.02). When after application of the highest limonene concentrations the lowest level of output had been reached, the subsequent washing out with pure saline reversed the effects but spiking activity and the amplitudes of spikes never gained the initial level completely.
DISCUSSION
The isolated AG5 in air-saturated saline sends through its peripheral nerves efferent Action Potentials (AP) of ventilatory motoneurons. Typical patterns of APs are periods of ventilatory bursting activity that alternate between the bilateral ventral nerves (N2) and the unpaired Median Nerve (MN) (Fig. 2). The two inspiratory motoneurons have their axons in the median nerve which branch more posterior bilaterally (left and right transversal nerve). The axons inside split as well and innervate the left and the right inspiratory muscles M225 (Snodgrass, 1935; Miller, 1960) which distend the sixth abdominal segment dorso-ventrally. Two smaller amplitude APs appear regularly but not in strict correlation to ventilation. They represent two motor axons to the spiracle closer muscles but spiracles of middle abdominal segments often remain closed during natural ventilation (Miller, 1960; Burrows, 1996). Motor activity in N2 supplies mainly three dorso-ventral expiratory muscles which in the fifth segment compress the fifth abdominal segment Ms 205,206,209 (Snodgrass, 1935) by pulling the sternite closer to and into the tergal lumen.
At laboratory temperatures between 20 and 25°C the mean frequency of the regular ventilatory bursting patterns from the isolated AG5 range between 1.4-0.27 Hz. Interruptions of bursting by up to 15 min of inactivity or long lasting tonic AP activity are possible in both nerves (Fig. 2b). That indicates that the ventilatory rhythm in the isolated fifth abdominal ganglion can be continuous or discontinuous (see: discontinuous ventilation) (Kestler, 1984) similar to the patterns found in intact locusts (Watts, 1951) and in the isolated metathoracic ganglia (Bustami and Hustert, 2000). At temperatures below 15°C bursting is interrupted regularly by long periods of inactivity (discontinuous ventilation) or by tonic discharges of motoneurons.
This study has two major aspects: (1) The suitability of this isolated ganglion (AG5) as a model system for tests of potential neurotoxic effects and (2) The insecticidal potential of essential oils containing monoterpenes from a common cultural plant.
The first seems to be proven unequivocally to us, although questions remain: (1) What is the primary drive for spontaneous ventilatory pattern generation in a single abdominal ganglion, (2) Where do the noxious effects find their target and (3) Why does the motor output change from bursting to tonic occasionally?
The second aspect supports ideas that biological terpenes may serve as an environmental friendly alternative to synthetic insecticides (Regnault-Roger, 1997; Isman, 2000; Isman and Machial, 2006; Liu et al., 2011; Regnault-Roger et al., 2012).
Previous insights into the physiological actions of potential biological insecticides were gathered by treatment of model insects (flies, cockroaches, beetles, bees) with essential oils of plants or the chemically pure form of their allochemicals. These partly caused symptoms suggesting a neurotoxic mode of action (Coats et al., 1991; Kostyukovsky et al., 2002). For detailed testing, mostly larger and easy to rear insects were used to develop specific organ preparations or isolated subsystems of the CNS where specific neuronal activities could be used to study the effects of neuroactive substances. Exposure or superfusion by different concentrations of plant extracts or fractions excites or inhibits the neurons, showing that there are noxious components (Enan, 2001, 2005; Kostyukovsky et al., 2002; Price and Berry, 2006).
In intact animals the CNS, including the segmental ganglia, controls most basic physiological functions such as respiration, circulation, heartbeat, gut motility, reproduction and control of posture (Lewis et al., 1973; Hustert, 1975; Weevers, 1985; Raabe, 1989; Burrows, 1996). Our specific neuronal testing system, the isolated locust abdominal ganglion (AG5), relies on one main parameter: changes of the persistent ventilatory output as recorded from peripheral nerves. The advantage of testing with the isolated CNS over testing whole animals or other organ systems (e.g., gut motility) is the absence of modulating sensory inputs and of coordinative influences by interneurons from other ganglia or the brain, which may be influenced by the test substance differently.
Ventilatory rhythms are generated by neuronal networks of ganglia in the CNS of insects. In intact locusts ventilatory pattern generation is distributed between different ganglia, with one leading pacemaker in the metathoracic ganglion (Miller, 1960). That ganglion coordinates by means of by command interneurons the ventilatory output of the other abdominal ganglia (Pearson, 1980), specifically in moderate to strong ventilation. No specific sensory receptors for respiratory gases that could control the ventilatory patterns and the motor output were ever found outside the insect CNS. Our results from isolated AG5 indicate that an intraganglionic sensitivity of neurons drives the network for ventilatory pattern generation and that each ganglion represents an autonomous system which is coupled to other pattern generators in the intact CNS. For the isolated AG5 any neurotoxic substances applied in the bath can influence directly or indirectly neurons of its intrinsic rhythm generating system. That was demonstrated here for essential oil from lovage plants (Levisticum officinale Koch.) and separately for their main allochemicals D-limonene and L-limonene.
Acute effects on neural activities: Essential oils are lipophilic and therefore should gain access through cell membranes to the neurons of a ganglion and produce direct intracellular effects. Basically, lovage oil caused decreased respiratory activity and this inhibitory effect can be washed out in pure saline but without complete return to the initial activity level. That indicates a residual permanent neurotoxicity, probably due to long-lasting lipophilic binding in cell membranes. In the insect ganglia, possible sites of influences after bath applications are also non-neuronal cells and their membranes such as glia of the ganglionic sheath serving partly as a blood-brain barrier (Treherne and Schofield, 1981) and internal glia cells all of which contribute to the ionic balance in the neuropile.
The direct effects that reduce AP amplitudes of axons must involve the membrane properties, which in turn influence the sodium-potassium-chloride balance across the membrane or the rate of transport through the fast sodium channels during action potentials. Ferber et al. (2001) reported that neurons in the insect Central Nervous System (CNS) and in peripheral nervous system are a major target of many toxins after their injection into the body. In general, the different toxins can act at various levels of neuronal metabolism, on cell membranes and their ion channels or on synaptic mechanisms of the pathways of neural transmission, such as many pesticides act in insects. The outcome can be over-excitation or depression of neural activity which is seen as abnormal motor output of the nervous system.
A decrease of ventilatory cycle periods could also be achieved by a different mechanism (Hou, 2011). In heatshock-preconditioned (HS) locusts the ventilatory muscle burst duration was increased after recovery from anoxic coma. Anoxia alone did not affect the activity or the localization of the Na+/K+-ATPase within the neuronal membranes but HS-preconditioning increased the total activity of the Na+/K+-ATPase and the localization of the Na+/K+-ATPase.
Persistent effects: Recovery of respiratory activity in the ganglion after washing out was only partial. The oils, if ingested by an insect, can be stored in fatty tissue due to their lipophilic properties, from where their components could spread gradually to the CNS and have long lasting effects and reduce also the ventilatory rhythm (Price and Berry, 2006). That may be, among others, a component that increases lethality after the uptake of lovage oil in locust hoppers. As a specific inhibitory mechanism of limonenes, the ventilation network could also be blocked reversibly (partially) from gas exchange or ionic diffusion through the ganglionic sheath (Treherne and Schofield, 1981). But the same may be true for other inhibited or even destroyed metabolic mechanisms under the influence of limonene inside the ganglion. Mills et al. (2004) refer to other monoterpenes rather than to D-and L-limonene as anti-cholinesterase inhibitors. On the contrary, Kostyukovsky et al. (2002) reported that limonene is ineffective in inhibiting acetylcholinesterase activity. Price and Berry (2006) indicated that in cockroaches the monoterpenoid eugenol acted as an octopamine mimetic or as an antagonist in its effects on dorsal unpaired median neurons (DUM), producing inhibition rather than excitation and spiking. Since other workers have found agonistic and antagonistic effects of eugenol (Enan, 2001, 2005) suggested that the oil affects specific sub-types of octopamine receptors. Octopamine is found throughout the ventral nerve cord, concentrated mainly within the ganglia but it also associated with a broad range of tissues, including muscular, secretory and reproductive tissues. Octopamine functions as a neurotransmitter, neurohormone and neuromodulator in insects, mediating also a sympathomimetic responses (Orchard, 1982). Neurotoxic and other less closely related mechanisms of essential oils were reviewed recently by Regnault-Roger et al. (2012).
Different efficiency of substances: The differences in neurotoxic effects of lovage oil, pure D-limonene, pure L-limonene and the 50/50% mixture of D- and L-limonene remain without a clear-cut explanation. The highest toxicity of the essential oil might be caused by additional components other than the main componenets D- and L-limonene. The difference between effects of the pure L- and D-forms of limonene is not surprising since differences in stereochemistry affect binding to other molecules could be the cause. More surprising is the lowered effect of their mix compared to the efficiency of the pure D- or L-limonene. Competitive binding effects may be involved but it is not clear how that may influence the neural activity of ventilatory motoneurones in different ways.
CONCLUSION
The study could conclude that lovage oil and limonene can be used as an effective insecticide where he has the ability to permeability and causing the toxic effect of insects. The effects of food with lovage can induce a reduced vitality of locusts, recognized here by the altered neuronal output from the respiratory rhythm generation in the CNS. We have shown that a ventilatory active ganglion of the CNS is influenced by super fusion with low concentrations of limonenes and lovage oil. So other subsystems of the nervous system which were not tested here could be affected similarly: neurons for sensory integration, for locomotion control or for neurohormonal functions. These all can summate to a vitality-reducing effect for the whole insect by limonenes-as pure substances or as major component of the essential oil from L. officinale.
AKNOWLEDGMENTS
The authors thanks and grateful to Peter Gumrich for technical support in this manuscript. This work was supported by the Ministry Higher Education, Missions Program for a Postdoctoral research scholarship (S.H.) from the Benha University, Egypt. http://www.bu.edu.eg/.
REFERENCES
- Akinwumi, F.O., E.A. Fasakin and C.O. Adedire, 2007. Toxic and repellence activities of four plant extracts to Dermestes maculatus degeer on smoked african mud catfish, Clarias gariepinus Burchell. J. Entomol., 4: 149-154.
CrossRefDirect Link - Bustami, H.P. and R. Hustert, 2000. Typical ventilatory pattern of the intact locust is produced by the isolated CNS. J. Insect Physiol., 46: 1285-1293.
CrossRefDirect Link - Ikbal, C., B.H. Mounia and B.H. Habib, 2007. Toxicity experiments of the saponic extract of Cestrum Parqui (Solanaceae) on some insect spices. J. Entomol., 4: 113-120.
CrossRefDirect Link - Clements, A.N. and T.E. May, 1974. Studies on locust neuromuscular physiology in relation to glutamic acid. J. Exp. Biol., 60: 673-705.
Direct Link - Clevenger, J.F., 1928. Apparatus for the determination of volatile oil. J. Am. Pharm. Assoc., 17: 345-349.
CrossRefDirect Link - El-Gendy, N.A. and E.A. Shaalan, 2012. Oviposition deterrent activity of some volatile oils against the filaria mosquito vector Culex pipiens. J. Entomol., 9: 435-441.
CrossRefDirect Link - Enan, E., 2001. Insecticidal activity of essential oils: Octopaminergic sites of action. Comp. Biochem. Physiol. Part C: Toxicol. Pharmacol., 130: 325-337.
CrossRefPubMedDirect Link - Enan, E.E., 2005. Molecular and pharmacological analysis of an octopamine receptor from American cockroach and fruit fly in response to plant essential oils. Arch. Insect Biochem. Physiol., 59: 161-171.
CrossRefPubMedDirect Link - Ferber, M., M. Horner, S. Cepok and W. Gnatzy, 2001. Digger wasp versus cricket: Mechanisms underlying the total paralysis caused by the predator's venom. J. Neurobiol., 47: 207-222.
CrossRef - Friedman, L. and J.G. Miller, 1971. Odor incongruity and chirality. Science, 172: 1044-1046.
CrossRef - Grundy, D.L. and C.C. Still, 1985. Inhibition of acetylcholinesterases by pulegone-1,2-epoxide. Pesticide Biochem. Physiol., 23: 383-388.
CrossRefDirect Link - Heth, G., E. Nevo, R. Ikan, V. Weinstein, U. Ravid and H. Duncan, 1992. Differential olfactory perception of enantiomeric compounds by blind subterranean mole rats (Spalax ehrenbergi). Experientia, 48: 897-902.
CrossRef - Hustert, R., 1975. Neuromuscular coordination and proprioceptive control of rhythmical abdominal ventilation in intactLocusta migratoria migratorioides. J. Comp. Physiol., 97: 159-179.
CrossRefDirect Link - Hustert, R. and A.M. Mashaly, 2013. Spontaneous behavioral rhythms in the isolated CNS of insects-Presenting new model systems. J. Physiol. Paris, 107: 147-151.
CrossRef - Isman, M.B., 2000. Plant essential oils for pest and disease management. Crop Protect., 19: 603-608.
CrossRefDirect Link - Kostyukovsky, M., A. Rafaeli, C. Gileadi, N. Demchenko and E. Shaaya, 2002. Activation of octopaminergic receptors by essential oil constituents isolated from aromatic plants: Possible mode of action against insect pests. Pest Manage. Sci., 58: 1101-1106.
CrossRefPubMedDirect Link - Koul, O., S. Walia and G.S. Dhaliwal, 2008. Essential oils as green pesticides: Potential and constraints. Biopestic. Int., 4: 63-84.
Direct Link - Laska, M. and C.G. Galizia, 2001. Enantioselectivity of odor perception in honeybees (Apis mellifera carnica). Behav. Neurosci., 115: 632-639.
CrossRef - Laska, M., C.G. Galizia, M. Giurfa and R. Menzel, 1999. Olfactory discrimination ability and odor structure-activity relationships in honeybees. Chem. Senses, 24: 429-438.
CrossRef - Laska, M., A. Liesen and P. Teubner, 1999. Enantioselectivity of odor perception in squirrel monkeys and humans. Am. J. Physiol., 277: R1098-R1103.
PubMed - Laska, M. and P. Teubner, 1999. Olfactory discrimination ability of human subjects for ten pairs of enantiomers. Chem. Senses, 24: 161-170.
CrossRef - Laska, M. and P. Teubner, 1999. Olfactory discrimination ability for homologous series of aliphatic alcohols and aldehydes. Chem. Senses, 24: 263-270.
CrossRef - Leitereg, T.J., D.G. Guadagni, J. Harris, T.R. Mon and R. Teranishi, 1971. Evidence for the difference between the odours of the optical isomers (+)-and (-)-carvone. Nature, 230: 455-456.
CrossRef - Lewis, G.W., P.L. Miller and P.S. Mills, 1973. Neuro-muscular mechanisms of abdominal pumping in the locust. J. Exp. Biol., 59: 149-168.
Direct Link - Liu, Z.L., S.S. Chu and G.H. Jiang, 2011. Insecticidal activity and composition of essential oil of Ostericum sieboldii (Apiaceae) against Sitophilus zeamais and Tribolium castaneum. Rec. Nat. Prod., 5: 74-81.
Direct Link - Miller, P.L., 1960. Respiration in the desert locust I. The control of ventilation. J. Exp. Biol., 37: 224-236.
Direct Link - Mills, C., B.V. Cleary, J.J. Walsh and J.F. Gilmer, 2004. Inhibition of acetylcholinesterase by tea tree oil. J. Pharm. Pharmacol., 56: 375-379.
CrossRef - Ngoh, S.P., L.E.W. Choo, F.Y. Pang, Y. Huang, M.R. Kini and S.H. Ho, 1998. Insecticidal and repellent properties of nine volatile constituents of essential oils against the American cockroach, Periplaneta americana (L.). Pesticide Sci., 54: 261-268.
CrossRefDirect Link - Orchard, I., 1982. Octopamine in insects: Neurotransmitter, neurohormone and neuromodulator. Can. J. Zool., 60: 659-669.
CrossRef - Ozaki, M., T. Takahara, Y. Kawahara, A. Wada-Katsumata and K. Seno et al., 2003. Perception of noxious compounds by contact chemoreceptors of the blowfly, Phormia regina: Putative role of an odorant-binding protein. Chem. Senses, 28: 349-359.
CrossRef - Pearson, K.G., 1980. Burst generation in coordinating interneurons of the ventilatory system of the locust. J. Comp. Physiol., 137: 305-313.
CrossRef - Prabuseenivasan, S., M. Jayakumar and S. Ignacimuthu, 2006. In vitro antibacterial activity of some plant essential oil. BMC Complementary Altern. Med., Vol. 6.
CrossRefDirect Link - Price, D.N. and M.S. Berry, 2006. Comparison of effects of octopamine and insecticidal essential oils on activity in the nerve cord, foregut and dorsal unpaired median neurons of cockroaches. J. Insect Physiol., 52: 309-319.
CrossRef - Regnault-Roger, C., 1997. The potential of botanical essential oils for insect pest control. Integ. Pest. Manage. Rev., 2: 25-34.
CrossRefDirect Link - Regnault-Roger, C., C. Vincent and J.T. Arnason, 2012. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol., 57: 405-424.
CrossRefDirect Link - Rubin, B.D. and L.C. Katz, 2001. Spatial coding of enantiomers in the rat olfactory bulb. Nat. Neurosci., 4: 355-356.
CrossRef - Ryan, M.F. and O. Byrne, 1988. Plant-insect coevolution and inhibition of acetylcholinesterase. J. Chem. Ecol., 14: 1965-1975.
CrossRefDirect Link - Soummane, H., M. Larhsini, K. Naamani and J. Coll, 2011. Studies of larvicidal and adulticidal activities of some Halophyte plant extracts against Ceratits capitata (Wiedemann). J. Entomol., 8: 548-556.
CrossRefDirect Link - Pandey, S., J.P. Pandey and R.K. Tiwari, 2012. Effect of some botanicals on hemocytes and molting of Papilio demoleus Larvae. J. Entomol., 9: 23-31.
CrossRef - Taniguchi, M., M. Kashiwayanagi and K. Kurihara, 1992. Quantitative analysis on odor intensity and quality of optical isomers in turtle olfactory system. Am. J. Physiol. Regul. Integr. Comp. Physiol., 262: R99-R104.
Direct Link - Treherne, J.E. and P.K. Schofield, 1981. Mechanisms of ionic homeostasis in the central nervous system of an insect. J. Exp. Biol., 95: 61-73.
Direct Link - Wawrzyniak, M. and R. Lamparski, 2006. Effect of Umbelliferae (Apiaceae) plant water extracts on colorado potato beetle (Leptinotarsa decemlineata Say) feeding and development. Electron. J. Polish Agric. Univ.
Direct Link - Watts, D.T., 1951. Intratracheal pressure in insect respiration. Ann. Entomol. Soc. Am., 44: 527-538.
Direct Link