
Mandumbu Ronald
Department of Agronomy, University of Fort Hare, P.O. Box X1314, 5700 Alice, South Africa
LiveDNA: 263.12613
Mutengwa Charles
Department of Agronomy, University of Fort Hare, P.O. Box X1314, 5700 Alice, South Africa
Mabasa Stanford
Department of Crop Science, University of Zimbabwe, P.O. Box MP167, Mount Pleasant, Harare, Zimbabwe
Mwenje Eddie
Department of Crop Science, Bindura University of Science Education, P.O. Box 1020, Bindura, Zimbabwe
Journal of Biological Sciences
Year: 2017 | Volume: 17 | Issue: 5 | Page No.: 194-201







ABSTRACT
Striga is a major parasitic weedy species in Southern Africa and is an impediment to attainment of household food security for poorly resourced communal farmers. The objective of this study was to use future niche descriptions and the life stages of Striga to predict the Striga epidemic in the future. Climate change projections through time scale analysis, general circulation models (GCM) down scaling and dynamical down scaling were used to predict the likely scenario in relation to the Striga epidemic. Agricultural systems are expected to face an increasing risk of erosion, runoff and soil degradation. Alternating high temperatures and rainfall may assist breaking of dormancy in Striga whilst severe winds greatly aid dispersal of the weed seeds. Generally production of strigolactones, haustorial initiation factors, attachments, seed production and dispersal were expected to increase as temperature rises like other biological processes. From this study it can be concluded that the Striga epidemic is going to increase under the new climate. The parasitic weed is likely going to become a more serious threat to crop production.
PDF Abstract XML References Citation
Received: May 24, 2017;
Accepted: August 28, 2017;
Published: September 09, 2017
Copyright: © 2017. This is an open access article distributed under the terms of the creative commons attribution License, which permits unrestricted use, distribution and reproduction in any medium, provided the original author and source are credited.
How to cite this article
Mandumbu Ronald, Mutengwa Charles, Mabasa Stanford and Mwenje Eddie, 2017. Predictions of the Striga Scourge under New Climate in Southern Africa: A Perspective. Journal of Biological Sciences, 17: 194-201.
DOI: 10.3923/jbs.2017.194.201
URL: https://scialert.net/abstract/?doi=jbs.2017.194.201
DOI: 10.3923/jbs.2017.194.201
URL: https://scialert.net/abstract/?doi=jbs.2017.194.201
INTRODUCTION
Striga is the major biological constraint to increased cereal production in the smallholder sector of Southern Africa. Striga species are a major parasitic weedy pest throughout the semi-arid sub Saharan Africa and many parts of Asia1. Many African countries including Tanzania, Kenya, Malawi, Madagascar, Botswana, Zimbabwe, Gabon, Nigeria, Ethiopia, Niger, Togo, Benin and Burkina Faso are highly infested with Striga causing serious yield losses that are as high as 100% at some sites2. In particular, the major crops that supply the bulk of the energy and protein needs of the poor in the African savannah, namely maize, sorghum, millets, upland rice and cowpeas have been severely vulnerable3.
Losses from Striga are compounded because of the tendency of crops grown under severe moisture and poor fertility conditions to show significant predisposition to Striga. According to Scholes and Press4 and Ejeta5 over 50 million hectares of arable farmland under cultivation with cereals and legumes in sub Saharan Africa are infested with one or more Striga species. In many of these places the Striga has reached epidemic proportions presenting a desperate situation in subsistence agriculture6. The weed causes annual losses of yield estimated to be in excess of US$10 billion5. The Striga spp. affects the welfare and livelihoods of over 100 million people in Africa1. According to Parker7 the weed has impacted on the sub region’s economy.
Over the years, many promising Striga control methods have been suggested in various formats, some suggestions appearing in multiple incarnations8. Despite this valuable work, adoption and utility of these control methods are limited, yield loss attributable to Striga is acute, perhaps even exacerbated ranging from 35-72% in some studies9. It is becoming very apparent that there is no silver bullet for Striga control. Despite its status as a serious parasitic weed work on the status of Striga under climate change is limited. Prior weed predictions done have not included Striga as a threat to food security under climate change. The objective of this paper is to use future niche descriptions to predict the Striga epidemic under the future climate. The authors speculate that the Striga epidemic is going to increase under the new climate.
Genus Striga: Striga is a latin word for ‘witch’ presumably because plants diseased by Striga display stunted growth and an overall drought like phenotype long before the weed appears. The genus was previously grouped within the family Scrophulariaceae but more recent analysis have placed the Striga under the family Orobanchaceae5,10. Striga possibly originates from a region between Semien Mountains of Ethiopia and the Nubian Hills of Sudan11. The same is the birth place of cultivated sorghum which is the major host species for several Striga spp.10. The main agriculturally important Striga spp. are Striga hermonthica (Del) benth and S. asiatica (L.) Kuntze in cereal crops and Striga gesnerioides (Willd) Vatke in cow peas. Striga aspera (Willd) Benth and Striga forbesii are also significant problems in cereals in limited locations7. The spread of various Striga spp. across the African continent is shown on Table 1.
Extent of the Striga problem in Southern Africa: Parasitic weeds are fast becoming a major constraint to many crops in Southern Africa and yet the efficacy of available means to control them are minimal. It has become one of the greatest biological constraints to food production in the drier parts of Africa, probably a more serious problem than insects, birds or plant diseases. The C4 cereal maize, sorghum, rice and millet are the preferred hosts and the infection of these plants by Striga spp. can result in severe grain losses. According to Rubiales et al.1, typical yield losses vary from 15-20% at a regional level but can be much more severe at local scales, sometimes resulting in total crop failure. The losses largely depend on the level of infection, crop variety, soil fertility and rainfall12.
It has been estimated that yield losses attributed to the weed exceed US$7 billion in value without accounting for the adverse effect on the welfare and livelihoods of over 100 million people in Africa1. The most affected are the resource poor, small scale subsistence farmers and severe infestations cause serious food shortages in Southern Africa. There are variable statistics on the extent of the Striga infestations by individual species but generally, 40% of arable land in sub Saharan Africa and 67% of the 73 million hectares in cereal zones is infested by Striga. Harsh conditions mean that few alternative crops are available and the use of high cost inputs such as herbicides are generally not feasible for resource poor farmers. The small scale famers’ cash investments in crop production are low. According to Ronald et al.13 inputs are low, rainfall is erratic and soils are poor and these conditions are the most suitable for Striga spp. to thrive.
In other areas the weed has reached epidemic proportions presenting a desperate scenario to small scale farmers. Where the scenario has worsened to these proportions the farmers are left with one option which is to abandon the land. According to Atera et al.14, demographic pressure has led to monocropping, thus increasing the frequency of Striga spp host crops in the cropping system, an ideal condition for Striga to thrive.
| Table 1: | Distribution and occurrence of Striga spp. in sub Saharan Africa |
 | |
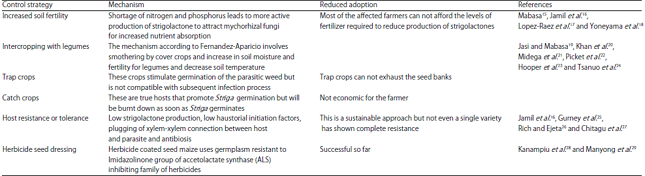
Managing Striga in Southern Africa: Despite the concerted efforts to come up with a sustainable method of Striga asiatica control, there is no sustainable control method available for managing the parasite. It remains as the number one biological constraint limiting the production of cereal grains in Southern Africa. Several methods have been studied but accompanied by minimum success. This is partly due to the complex life cycle of Striga, which is intimately linked to its host and depends on the response to chemical and tactile cues posing a challenge to control of the weed both prior and after attachment to the host. The numerous methods that have been suggested or developed together with their technical limitation are shown in Table 2.
Prediction method: Niche based model: The niche based model is based on the knowledge for suitable habitat beyond the species’ current distribution or severity. It is a pattern based indicator30. The first task is to use the current data on distribution to predict the severity of spread under current conditions then use niche descriptions to predict the level of spread under future climate.
The second task according to Crossman et al.30 is to use expert opinion to quantify dispersibility and identify the level of the epidemic expected. For Striga spp. the life cycle is a multistage ranging from dormancy breaking, strigolactones release, stimulation of germination, haustorial formation and attachment, its life under ground and life outside after emergence then seed production and dispersal. The objective is to try and predict the level of occurrence of each stage under the current climate and under future climate. Whilst Striga is a threat now predictions have to be made to check the level of spread under future climate. The current climate may be suboptimal preventing the weed from reaching epidemic levels or they may be optimal conditions and when climate changes the weed may deteriorate in status or infections.
Current climate and expected future climate in Southern Africa: Southern Africa is predominantly a semi-arid region with high rainfall variability, characterized by frequent drought and floods. According to Davis31 there is a high degree of spatial variation in rainfall across southern Africa and the average for the region is just less than 1000 mm/year. The highest amount is 3100 mm and the lowest is less than 100 mm/year. The majority of the region is between 500-1500 mm/year. Southern African rainfall shows a clear seasonal characteristics with a large part of the subcontinent experiencing a summer rainfall season, usually commencing in October/November and tapering off in February and march32.
| Table 2: | Striga control strategies, mechanisms and causes of reduced adoption |
 | |
The Southern African region exhibits a largely warm climate, with warm average temperature mostly above 17°C with exceptions of high altitudes and coastal areas. Given bellow is a summary of the climate change projections using various techniques for predictions according to Tedross et al.33 (Table 3).
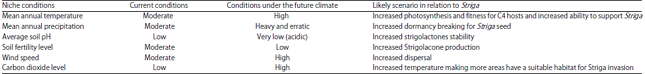
The future conditions may make niches for the Striga spp. When temperatures rise, they may provide a suitable habitat for the Striga parasite to increase or decrease its abundance. The implications of the niche to changes in climate are given in Table 4. When temperature increases other crops like wheat which are currently not under attack by the Striga because it is planted in winter may be susceptible. Vasey et al.34 found wheat to be susceptible to Striga hermonthica and questions the implications of this to wheat production areas under climate change in Africa. A summary of the niche descriptions for the future are shown in Table 4.
Striga epidemic under climate changeSeed dormancy and after ripening: Striga seeds have an after-ripening requirement and do not germinate in the season in which they are produced. Rich and Ejeta26 asserts that this requirement prevents newly matured seed from germinating too late in the season when host plants capable of supporting the parasitic plants to maturity are scarce. Striga seeds must go through a phase of conditioning10. Peak germination of Striga seeds occur in vitro after 10-15 days of soaking in water at a temperature of 28°C. Sun et al.35 reported that pre-conditioning at a suitable temperature releases the dormancy within 2-3 weeks and increases the sensitivity of Striga seed to strigolactones by several orders of magnitude. If no strigolactone is received during this time, the Striga seeds will eventually fall into secondary dormancy.
Germination is linked to the presence of the host that is nearby as the endosperm of Striga can not sustain growth for only 3-7 days. Striga seeds generally are dormant and the dormancy is broken down by alternating wet periods and high temperature exposure. Under climate change the alternating wet and hot conditions are predicted to be a norm hence dormancy will easily be broken and germination rates will be increased.
Strigolactone biosynthesis and exudation: Nutrient deficiencies have profound effects on strigolactones biosynthesis and exudation. According to Yoneyama et al.36, roots of host plants grown in phosphorus deficient soils were found to produce more strigolactones.
| Table 3: | Climate change projections using various techniques for Southern Africa |
 | |
| Table 4: | Niche conditions under current and future climate |
 | |
More root parasitic plants generally prevail on nutrient deficient soils and thus fertilizer application suppresses their emergence. Yoneyama et al.36 reports of a 100 fold increase in germination stimulation activity of root exudates from plants grown under phosphate starvation. Yoneyama et al.36 further confirms that in Sorghum bicolor suffering from phosphate deficiency enhances strigolactones production particularly the case of 5-deoxystrigol and sorghumol. The same was confirmed by several authors that nitrogen and phosphorus shortage enhances strigolactones biosynthesis26.
With climate change serious land degradation is expected which can result in heavy leaching and degradation of soils. Plant growing under that environment would require strong cooperation with AM fungi for increased adsorption of these nutrients. Therefore, cereal crops growing under degraded soils are likely to produce more strigolactones. Soil degradation is expected to alter soil pH to more acidic levels. Although the effects of soil pH on Striga germination have not been studied adequately. The projected increases in soil degradation in Africa may therefore indirectly four Striga spp.37,38. Germination of Striga seeds in agar was obtained at a pH of 4. Loss of activity of synthetic germination stimulant (GR 7) was lost in alkaline soils. This may point to more strigolactones production under acidic conditions and the increased stability of strigolactones under the same environment which is a common feature under degraded soils.
Zwaneburg et al.39 and Bouwmeester et al.40 found that strigolactones are unstable in watery environments. With the alternating periods of very wet and dry conditions under climate change, germination may occur under dry conditions phase. Once germinated the Striga grows under ground for between 4-8 weeks and so the Striga may continue to grow once germinated.
Various authors have reported that maximum germination occurs at temperatures between 30-35°C. Currently the average temperature is bellow 28°C in Southern Africa and thus with increased carbon dioxide enrichment in the atmosphere, temperature changes are likely to lead to increased germination of Striga as long as they bellow 35°C. Hence the 2-3°C increase expected is likely going to increase germination of Striga.
Haustorial inducing factors: Haustoria are invasive structures that develop at the tip of Striga radicles in response to host root contact. The formation and penetration of host root by Striga is stimulated by haustorial inducing factors. These are compounds such as quinines, flavonoids and phenolics. The only haustorial inducing factor isolated in plants is 2-4 dimethoxybenzoquinone and it comes from the oxidative degradation of lignin and decarboxylation of phenolic acids.
The quinines, flavonoids and phenolics are called allelochemicals and are produced under extreme conditions such as higher temperature and exposure to extreme radiation. With these expected to increase under climate change the production of haustorial inducing factors is expected to increase. This may lead to an increase in number of Striga plants successfully attaching to the host.
Seed production and dispersal: Hearne8 reported that Striga has a high reproductive capacity producing between 10 000-200 000 seeds/plant. These seeds are small with dimensions of 0.3 and 0.15 mm. the seeds are very light weighing 4-7 μg and are easily dispersed by winds, water and animals. With strong winds expected under climate change long distance dispersal is expected to be more. The seed is likely going to be spread to new areas as a result of wing dispersal.
With increased temperatures and carbon dioxide expected most C4 host plants are expected to increase in productivity and Striga productivity depends on food availability. The large quantities of long lived seeds will ensure that the parasite adapt to changes in host availability, consequently making them more difficult to control34.
Perspective: The current ecological drivers for Striga presently are temperature, degraded soils with acidic pH, winds that are prevalent for dispersal and precipitation. All these ecological attributes are predicted to change under the future climate therefore the Striga scourge may also change. Laboratory experiments done found that more acidic pH increases the stability of strigolactones, more prevalent winds increase dispersal. Extremes of temperature may limit the growing of temperature sensitive plants, farmers may concentrate on C4 plants (sorghum, millets and maize) which are hosts.
Although it is difficult to predict with certainty the level of Striga epidemic under the future climate, life stages of Striga may be promoted under the future climate. Whilst Striga is a problem in particular areas at the present moment it is likely that the width of its niche is likely to increase under the changed climate. For Southern Africa, a 4°C increase in the average temperature may move the average to 34°C for most areas. Most of the biological processes involved in Striga infection occur at their optimum at that temperature. Soil degradation that is predicted under the future climate may move soil pH to more acidic levels. Mohamed et al.41 have predicted that Striga distribution might in the long run expand to moderate climate zones at the expense of tropical and sub-tropical areas. Due to the fact that the life cycle of the weed is multistage, it is very difficult to assume that all stages are going to be enhanced under the new climate. Detailed laboratory experiments may need to be done to determine the conditions with respect to water availability, temperature and wind speed that give optimum responses to particular life stages of the Striga life cycle.
CONCLUSION
It was concluded that under the new climate, the Striga epidemic may worsen. The various conditions under the new climate may worsen the Striga scourge as they seem to favor the various life stages of the parasite. Striga spp. may start to be a weed of economic importance in areas where it was not found.
SIGNIFICANCE STATEMENTS
This study predicts that Striga parasitism is going to worsen under the new climate. Such studies are limited in literature. That knowledge is beneficial in that the weed managers and the farming community have to be prepared to deal with the epidemic. This study will help the researcher to link every life stage of Striga to the expected future conditions that many researchers did not explore.
ACKNOWLEDGMENTS
The Research Council of Zimbabwe (grant number 2015/4) and the Bindura University of Science Education Post Graduate and Research Centre are acknowledged for funding the research.
REFERENCES
- Rubiales, D., J. Verkleij, M. Vurro, A.J. Murdoch and D.M. Joel, 2009. Parasitic plant management in sustainable agriculture. Weed Res., 49: 1-5.
CrossRefDirect Link - Scholes, J.D. and M.C. Press, 2008. Striga infestation of cereal crops-an unsolved problem in resource limited agriculture. Curr. Opin. Plant Biol., 11: 180-186.
CrossRefPubMedDirect Link - Ronald, M., M. Charles, M. Stanford and M. Eddie, 2016. Existence of different physiological ‘strains’ of Striga asiatica (L.) Kuntze on sorghum species [Sorghum bicolor (L.) Moench and Sorghum arundinaceum (Desv.) Stapf] in Zimbabwe. Res. Crops, 17: 468-478.
CrossRef - Parker, C., 2009. Observations on the current status of Orobanche and Striga problems worldwide. Pest Manage. Sci., 65: 453-459.
CrossRefDirect Link - Hearne, S.J., 2009. Control-the Striga conundrum. Pest Manage. Sci., 65: 603-614.
CrossRefDirect Link - De Groote, H., L. Wangare, F. Kanampiu, M. Odendo, A. Diallo, H. Karaya and D. Friesen, 2008. The potential of a herbicide resistant maize technology for Striga control in Africa. Agric. Syst., 97: 83-94.
CrossRefDirect Link - Spallek, T., M. Mutuku and K. Shirasu, 2013. The genus Striga: A witch profile. Mol. Plant Pathol., 14: 861-869.
CrossRefDirect Link - Atera, E.A., K. Itoh and J.C. Onyango, 2011. Evaluation of ecologies and severity of Striga weed on rice in sub-Saharan Africa. Agric. Biol. J. North Am., 2: 752-760.
Direct Link - Menkir, A. and J.G. Kling, 2007. Response to recurrent selection for resistance to Striga hermonthica (Del.) Benth in a tropical maize population. Crop Sci., 47: 674-682.
CrossRefDirect Link - Ronald, M., M. Charles, M. Stanford and M. Eddie, 2017. Effect of witchweed (Striga asiatica L. Kuntze) infestation and moisture stress on selected morpho-physiological traits of sorghum (Sorghum bicolor L. Moench) genotypes in Zimbabwe. J. Agron., 16: 65-75.
CrossRefDirect Link - Atera, E.A., K. Itoh, T. Azuma and T. Ishii, 2012. Farmer's perspectives on the biotic constraint of Striga hermonthica and its control in Western Kenya. Weed Biol. Manage., 12: 53-62.
CrossRefDirect Link - Jamil, M., J. Rodenburg, T. Charnikhova and H.J. Bouwmeester, 2011. Pre-attachment Striga hermonthica resistance of New Rice for Africa (NERICA) cultivars based on low strigolactone production. New Phytol., 192: 964-975.
CrossRefDirect Link - Lopez-Raez, J.A., R. Matusova, C. Cardoso, M. Jamil and T. Charnikhova et al., 2009. Strigolactones: Ecological significance and use as a target for parasitic plant control. Pest Manage. Sci., 65: 471-477.
CrossRefDirect Link - Yoneyama, K., K. Yoneyama, Y. Takeuchi and H. Sekimoto, 2007. Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta, 225: 1031-1038.
CrossRefDirect Link - Jasi, L. and S. Mabasa, 2001. The Status of Striga (Witchweed Research and Extension in Zimbabwe). In: Striga Research in Southern Africa and Strategies for Regionalized Control Options, Mgonja, M.A., O.A. Chivinge and E.S. Monyo (Eds.). International Crop research Institute for the Semi-Arid Tropics, India, pp: 35-47.
- Khan, Z.R., C.A.O. Midega, A. Hassanali, J.A. Pickett, L.J. Wadhams and A. Wanjoya, 2006. Management of witchweed, Striga hermonthica and stemborers in sorghum, Sorghum bicolor, through intercropping with greenleaf desmodium, Desmodium intortum. Int. J. Pest Manage., 52: 297-302.
CrossRefDirect Link - Midega, C.A.O., Z.R. Khan, D.M. Amudavi, J. Pittchar and J.A. Pickett, 2010. Integrated management of Striga hermonthica and cereal stemborers in finger millet (Eleusine coracana (L.) Gaertn.) through intercropping with Desmodium intortum. Int. J. Pest Manage., 56: 145-151.
CrossRefDirect Link - Pickett, J.A., M.L. Hamilton, A.M. Hooper, A. Khan and C.A.O. Midega, 2010. Companion cropping to manage parasitic plants. Annu. Rev. Phytopathol., 48: 161-177.
CrossRefDirect Link - Hooper, A.M., M.K. Tsanuo, K. Chamberlain, K. Tittcomb and J. Scholes et al., 2010. Isoschaftoside, a C-glycosylflavonoid from Desmodium uncinatum root exudate, is an allelochemical against the development of Striga. Phytochemistry, 71: 904-908.
CrossRefDirect Link - Tsanuo, M.K., A. Hassanali, A.M. Hooper, Z. Khan, F. Kaberia, J.A. Pickett and L.J. Wadhams, 2003. Isoflavanones from the allelopathic aqueous root exudate of Desmodium uncinatum. Phytochemistry, 64: 265-273.
CrossRefDirect Link - Gurney, A.L., D. Grimanelli, F. Kanampiu, D. Hoisington, J.D. Scholes and M.C. Press, 2003. Novel sources of resistance to Striga hermonthica in Tripsacum dactyloides, a wild relative of maize. New Phytol., 160: 557-568.
CrossRefDirect Link - Rich, P.J. and G. Ejeta, 2008. Towards effective resistance to Striga in African maize. Plant Signal. Behav., 3: 618-621.
CrossRefDirect Link - Chitagu, M., J.T. Rugare and S. Mabasa, 2014. Screening maize (Zea mays) genotypes for tolerance to witchweed (Striga asiatica L. Kuntze) infection. J. Agric. Sci., 6: 160-169.
CrossRefDirect Link - Kanampiu, F.K., J.K. Ransom, D. Friesen and J. Gressel, 2002. Imazapyr and pyrithiobac movement in soil and from maize seed coats to control Striga in legume intercropping. Crop Prot., 21: 611-619.
CrossRefDirect Link - Crossman, N.D., B.A. Bryan and D.A. Cooke, 2011. An invasive plant and climate change threat index for weed risk management: Integrating habitat distribution pattern and dispersal process. Ecol. Indicators, 11: 183-198.
CrossRefDirect Link - Tedross, M., C. Davis, F. Engelbretchm, A. Joubert and E.A. van Gareren, 2011. Regional Scenarios of Future Climate Change over Southern Africa. In: Climate Risk Vulnerability: A Handbook for Southern Africa, Davis, C. (Ed.). Council for Scientific and Industrial Research, Pretoria, South Africa, ISBN: 978-0-620-50627-4, pp: 28-50.
- Vasey, R.A., J.D. Scholes and M.C. Press, 2005. Wheat (Triticum aestivum) is susceptible to the parasitic angiosperm Striga hermonthica, a major cereal pathogen in Africa. Phytopathology, 95: 1294-1300.
CrossRefDirect Link - Sun, Z., J. Hans, M.H. Walter, R. Matusova and J. Beekwilder et al., 2008. Cloning and characterisation of a maize carotenoid cleavage dioxygenase (ZmCCD1) and its involvement in the biosynthesis of apocarotenoids with various roles in mutualistic and parasitic interactions. Planta, 228: 789-801.
CrossRefDirect Link - Yoneyama, K., X. Xie, D. Kusumoto, H. Sekimoto, Y. Sugimoto, Y. Takeuchi and K. Yoneyama, 2007. Nitrogen deficiency as well as phosphorus deficiency in sorghum promotes the production and exudation of 5-deoxystrigol, the host recognition signal for arbuscular mycorrhizal fungi and root parasites. Planta, 227: 125-132.
CrossRefDirect Link - Mohamed, K.I., M. Papes, R. Williams, B.W. Benz and A.T. Peterson, 2006. Global invasive potential of 10 parasitic witchweeds and related orobanchaceae. AMBIO: J. Hum. Environ., 35: 281-288.
CrossRefDirect Link - Rodenburg, J., C.R. Riches and J.M. Kayeke, 2010. Addressing current and future problems of parasitic weeds in rice. Crop Prot., 29: 210-221.
CrossRefDirect Link - Zwanenburg, B., A.S. Mwakaboko, A. Reizelman, G. Anilkumar and D. Sethumadhavan, 2009. Structure and function of natural and synthetic signalling molecules in parasitic weed germination. Pest Manage. Sci., 65: 478-491.
CrossRefPubMedDirect Link - Bouwmeester, H.J., C. Roux, J.A. Lopez-Raez and G. Becard, 2007. Rhizosphere communication of plants, parasitic plants and AM fungi. Trends Plant Sci., 12: 224-230.
CrossRefDirect Link - Mohamed, A., A. Ellicot, T.C. Housley and G. Ejeta, 2003. Hypersensitive response to Striga infection in Sorghum. Crop Sci., 43: 1320-1324.
CrossRefDirect Link