
C. Prasanna Kumar
Centre for Advanced Study in Marine Biology, Faculty of Marine Biology, Annamalai University, Parangipettai, Tamil Nadu, India
B. Akbar John
Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
S. Ajmal Khan
Centre for Advanced Study in Marine Biology, Faculty of Marine Biology, Annamalai University, Parangipettai, Tamil Nadu, India
P.S. Lyla
Centre for Advanced Study in Marine Biology, Faculty of Marine Biology, Annamalai University, Parangipettai, Tamil Nadu, India
B.Y. Kamaruzzaman
Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
K.C.A. Jalal
Department of Biotechnology, Kulliyyah of Science, International Islamic University Malaysia, Jalan Sultan Ahmad Shah, Bandar Indera Mahkota, 25200 Kuantan, Pahang, Malaysia
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 1 | Page No.: 26-32







ABSTRACT
Cultivable marine bacteria associated with the sponge Hyattella cribriformis (Hyatt, 1877) was studied through 16S rRNA gene sequencing. These marine bacterial colonies produced on Zobell Marine agar plates were distinguished based on phenotypic characters viz., colony morphology and pigmentations of the individual colonies. The extracted DNA from the individual bacterial isolates was PCR amplified using universal 16S primers and subjected to DNA sequencing. BLAST (Basic Local Alignment Tool) analysis finds that Vibrio diazotrophicus, Bacillus subtilis, B. firmus, Thalassomonas agarivorans, Oleiphilus messinensis, Planococcus maritimus and Brevundimonas vesicularis are the culturable marine bacteria associated with the sponge Hyattella cribriformis. The phylogram constructed clearly delineated the bacterial isolates into its corresponding phylum. The cultivable bacterial density of Hyattella cribriformis was found to be 65x10-5 CFU mL-1. Submitted sequences in NCBI were assigned with accession numbers (FJ834327, FJ834328, FJ834329, FJ845392, FJ845393, FJ845394 and FJ845395). It could be concluded that, application of 16S rRNA sequencing approach would yield novel insights into the diversity of bacteria associated with sponges and hence would help in exploring the commercial importance and their industrial applications.
PDF Abstract XML References Citation
Received: October 01, 2012;
Accepted: October 05, 2012;
Published: February 09, 2013
How to cite this article
C. Prasanna Kumar, B. Akbar John, S. Ajmal Khan, P.S. Lyla, B.Y. Kamaruzzaman and K.C.A. Jalal, 2013. Cultivable Marine Bacterial Isolates from a Sponge Hyattella cribriformis. Journal of Biological Sciences, 13: 26-32.
DOI: 10.3923/jbs.2013.26.32
URL: https://scialert.net/abstract/?doi=jbs.2013.26.32
DOI: 10.3923/jbs.2013.26.32
URL: https://scialert.net/abstract/?doi=jbs.2013.26.32
INTRODUCTION
Sponges (Phylum Porifera) are evolutionarily ancient metazoans whose origin dates back about 600 million years to the Precambrian. With 7,000 formally described and an estimated 15,000 extant species, sponges are among the most diverse marine invertebrate groups and are important components of all aquatic habitats including freshwater environments, tropical reefs and even the deep sea. Numerous sponges live in permanent and close associations with microorganisms and many of them host phylogenetically diverse populations of microbes (Hentschel et al., 2006; Taylor et al., 2007). Sponge-bacteria associations are profound and evolutionary ancient (Wilkinson, 1987). Sponges harbor large amount of bacteria in their tissues that can amount to 40% of their total biomass exceeding that of sea water by two to three order of magnitude. Symbiotic functions that have been attributed to microbial symbionts include nutrient acquisition, stabilization of sponge skeleton, processing of metabolic waste and secondary production (Vacelet et al., 1995; Beer and Ilan, 1998; Schmid et al., 2000). In most cases sponges and bacteria can not be segregated even in cell lines (Mitova et al., 2004). The associated forms are normally multidrug resisting beside novel source of bioactive components (Thakur et al., 2004; Selvin and Lipton, 2004).
In the present study, an attempt was made to study the cultivable bacterial diversity of the sponge Hyattella cribriformis belonging to family Spongiidae. The body of this sponge is covered by unevenly punctured dermal membrane (Thomas, 1973). The culture independent methods like 16S rRNA-DGGE sequencing are commonly used to find out the precise diversity of sponge associated microbial community. In the present study, the sponge-associated microbial community via cultivation-based approach has been studied in addition to 16S rDNA sequencing. Phylogeny of the microbial community was studied by constructing phylogram using 16S rDNA sequences obtained.
MATERIALS AND METHODS
Sample collection and bacteria isolation: The sponges were collected by skin diving in the shallow waters of the Palk Bay (lat. 9°17’N long. 79°17’E). The specimens were stored immediately in sterile air tight bags and transported to laboratory as quickly as possible. The sponge was identified as Hyattella cribriformis (Hyatt 1877) using sponge identification manual (Thomas, 1973). About 1 g of wet sponge biomass was crushed with the aid of sterile pestle and mortar in the presence of 10 mL of sterile sea water in laminar air flow condition. One microliter of crushed soup was transferred through series of 9 mL autoclaved sea water in screw capped test tubes and subjected to serial dilution (up to 10-9 dilution). Triplicate plates of Zobell Marine Agar 2216 (Himedia) were made for each dilution and the plates were incubated at 37°C for 24 h. Following incubation, the culture was segregated based on phenotypic and pigmentation characteristics. Each individual colony was purified by quadrant streaking in Zobell Marine Agar 2216 (Himedia) plates.
DNA extraction and PCR: Standard phenol chloroform protocol was adopted to isolate the total genomic DNA of the marine bacteria (Chomczynski and Sacchi, 2006). Primers 16S1 (5’-GAGTTTGATCCTGGCTCA-3’) and 16S2 (5’-ACGGCTACCTTGTTACGACTT-3’) were used to amplify the 16S rRNA gene of the marine bacterial communities. Polymerase chain reactions were conducted at 95°C for 12 min, 40 cycles of 95°C for 30 sec, 52°C for 1 min and 68°C for 1 min 30 sec. About 10 μL PCR amplicons were gel checked using 0.8% agarose gel with 3 μL Ethidium Bromide as staining dye and 5 μL of Bromothymol blue as tracking dye.
DNA sequencing: Sequencing reaction was prepared in 0.2 mL thin-wall tubes. All the reagents were kept in ice during the preparations and added in order. QIAGEN-QIAquickTM was used to prepare DNA for sequencing reaction and protocol followed was based on manufacturer’s instruction. About 10 μL of dH2O, 10 μL of template DNA, 2 μL of 16S1 primer and 8 μL of DTCS quick start master mix (Provided with the QIAGEN-QIAquickTM kit) was taken in the tube and thoroughly mixed. The liquid was consolidated at the bottom of the tube by centrifuging before thermal cycling reaction. The single strand of amplified 16S rRNA gene was performed using the following thermo-cyclic conditions; 30 cycles of initiation at 96°C for 20 sec, annealing at 50°C for 20 sec and extension at 60°C for 4 min followed by holding at 4°C. The sequencing reaction was transferred to appropriately labeled 0.5 mL microcentrifuge tubes and mixed thoroughly. About 60 μL of cold 95% (v/v) ethanol/dH2O from -20°C freezer was added and mixed thoroughly. The tubes were immediately centrifuged at 14,000 rpm at 4°C for 15 min. The pellets were carefully removed with the micropipette and rinsed twice with 200 μL 70% (v/v) ethanol from -20°C freezer and immediately centrifuged at 14,000 rpm at 4°C for 2 min between each rinses. The pellet was vacuum dried for 10 min and the sample was re-suspended in 40 μL of sample loading solution (provided with the kit). The re-suspended sample was transferred into appropriate wells of the sequencing plates. Each re-suspended sample was over laid with a drop of mineral oil (provided with the kit). The sequencing plate was loaded into the Mega Bace 1000 automated high-throughput sequencer machine and the sequencing reactions were initiated.
Following sequencing reaction, the sequences were manually double checked using Chromas Pro software version 1.4. The sequence alignment and phylogram construction was carried out using MEGA (Molecular Evolutionary Genetic Analysis) version 4.0 (Tamura et al., 2007).
RESULTS
Bacterial pure culture, DNA isolation and PCR: The triplicate culture plates of 10-5 dilution had optimal number (countable numbers) of isolated colonies (N = 65) were selected. About 7 different colonies were differentiated and isolated based on its phenotypic variations (colony morphology and pigmentations). The isolated colonies were pure cultured in Zobell Marine agar slants and Gram stained using Hi-Media Gram stain Kit. Phenol-Chloroform methodology for DNA isolation gave good success as few loops of culture gave good quantity of DNA for PCR analysis. The universal primers 16S1 and 16S2 successfully gave amplicons (Fig. 1) at the size of 1500 bp measured in the gel with the aid of 100 bp DNA ladder as standard marker.
 | |
| Fig. 1: | PCR amplicons of 16S rDNA gel checked using 1% agarose gel, Lane 1-7: The sample lanes, Lane M: 100 bp ladder |
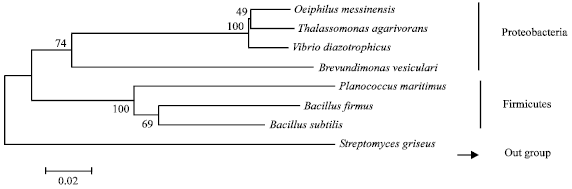 | |
| Fig. 2: | Phylogram showing the relationship among the culturable marine bacterial strains associated with Hyattella cribriformis, The numbers at the nodes of the tree indicate the bootstrap vales of the corresponding nodes, The tree is drawn with the minimum evolutionary rate of 0.1% |
| Table 1: | BLAST analysis along with Grams reaction and accession numbers assigned by NCBI |
 | |
BLAST analysis and phylogeny: The species of the bacteria were defined after the BLAST (http://blast.ncbi.nlm.nih.gov/blast.cgi) analysis. 16S rDNA queried via BLAST revealed high success rate as the minimum percentage of similarity was 98% and the maximum was cent percent. The results of BLAST analysis, closest match and corresponding accession numbers assigned for submitted sequences are given in Table 1. Vibrio diazotrophicus, Bacillus subtilis, B. firmus, Thalassomonas agarivorans, Oleiphilus messinensis, Planococcus maritimus and Brevundimonas vesicularis are the culturable marine bacteria found associated with the sponge Hyattella cribriformis.
For phylogenetic studies, 16S rDNA sequence of Streptomyces griseus (FJ796405) was chosen as an out group to test the reliability of the phylogram. The Phylogram were clearly distinguished in to two main clades, one consisting of Hyattella cribriformis associated marine bacteria and the second consisting of chosen Actinobacterial out group (Fig. 2).
The constructed phylogram showed two distinct clades, correspondingly distinguishing two phyla; Proteobacteria and Firmicutes. Among the 7 bacteria isolated from Hyattella cribriformis, majority of culturable strains was Proteobacteria (Thalassomonas agarivorans, Oleiphilus messinensis, Vibrio diazotrophicus and Brevundimonas vesicularis) as they were known to dominate sponge associated bacterial communities (Hill et al., 2006). The other 3 strains (Bacillus subtilis, B. firmus and Planococcus maritimus) belonged to the phylum Firmicutes. Among the Proteobacterial communities were Gammaproteobacteria except Brevundimonas vesicularis which belonged to Alphaproteobacterial group. Firmicutes community consisted of two members from Bacillaceae and one from Planococcaceae. The members of Bacillaceae branched together leaving the other member of Planococcaceae in different clad.
DISCUSSION
The implementation of the 16S rDNA approach has revolutionized the field of microbial Ecology. Through the use of 16S rDNA markers as a phylogenetic marker, it has been made possible to determine the precise phylogenetic position of marine environmental bacterial populations independent of their complex niches (symbiotic or free living). It was noted that Proteobacteria are predominant in the marine environment and are usually associated with marine plants and animals (Sekiguchi et al., 2002; Weidner et al., 2000; Lau et al., 2002). From the present study 4 culturable marine Proteobacteria (Thalassomonas agarivorans, Oleiphilus messinensis, Brevundimonas vesicularis and Vibrio diazotrophicus) have been found to be associated with the sponge Hyattella cribriformis.
Thalassomonas agarivorans is a non-fermentative, gram-negative bacterium which mandatorily requires 3% NaCl for its growth. It grows aerobically and is not capable of anaerobic growth by fermenting glucose or other carbohydrates (Jean et al., 2006). This bacterium is also known for its agarolytic activity in the absence of alternative carbon source. Recently a novel β agarase enzyme has been patented from T. agarivorans but no such activity was seen in the present study, as Zobell Marine agar contains multiple carbon sources. O. messinensis is a gram-negative, aerobic, motile, rod-shaped bacterium which uses a narrow spectrum of organic compounds, including aliphatic hydrocarbons, alkanoates and alkanoles, as carbon and energy sources (Golyshin et al., 2002). This bacterium is considered to be the most enigmatic marine bacteria, initially isolated from harbor sediments seem also to thrive elsewhere as a sponge symbiont (Golyshin et al., 2002). Oleiphilus-like 16S rRNA gene fragments have been recovered from bryozoan and dictyoceratida sponges sampled in North Atlantic and equatorial Pacific oceans (Ridley et al., 2005). The bio-flim forming ability of O. messinensis during the cultivation in n-alkane media was also reported and hence it could be a member of bio-flim forming community (Golyshin et al., 2002).
Proteobacteria associated with sponges are believed to communicate with each other via the phenomenon of quorum signaling pathways (Mohamed et al., 2008). Vibrio diazotrophicus and Thalassomonas agarivorans are known to involve in quorum signaling pathways by producing N-acyl homoserine lactones. Vibrio diazotrophicus is a gram-negative, short rod bacterium motile by means of a single polar flagellum. It is also known for its ability to fix nitrogen (Guerinot et al., 1982). Occurrence of V. diazotrophicus in clams, fresh and frozen sea foods were also reported (Hidalgo et al., 2008; Ottaviani et al., 2001). Some sponges like Ircinia felix and Aplysina cauliformis are known to utilize the nitrogen fixed by its associated microbial communities. However, the ability of Hyattella cribriformis in establishing such relationships with V. diazotrophicus needs to be proved.
Brevundimonas vesicularis, isolated in the present study is a gram-negative, short rod bacterium known for its infrequent human infection. The genus Brevundimonas corresponds to the least nutritionally versatile bacteria studied (Segers et al., 1994). This species occur typically in freshwater and marine habitats with low nutrient levels. The results of DNA-rRNA hybridization studies, 16S rRNA cataloging and 16S rRNA sequencing indicated that this species belong to a separate genus in the alpha subclass (rRNA superfamily IV) of the Proteobacteria, for which the name Brevundimonas was proposed (Segers et al., 1994). Bacillus vesicularis has been implicated in rare cases of human infections (Han and Andrade, 2005). It rarely causes meningitis in humans due to resistance to piperacillin, gentamicin and amikacin as well as to cephalosporins, aztreonam, imipenem and meropenem (Mondello et al., 2006). But it is not yet clear whether they are true pathogens (Chi et al., 2004). Hence, presence of B. vesicularis among the population of H. cribriformis associated bacteria is surprising. Brevundimonas spp., possess features that make them amenable to air dispersal. The common airborne microbes included several species of Brevundimonas spp. (Tringe et al., 2008). Brevundimonas sp. has also been cultivated from nominally sterile environments such as the space station Mir and in clinical settings where they have been implicated in opportunistic infection (Han and Andrade, 2005; Kawamura et al., 2001).
Bacillus vesicularis has also been isolated from the biofilms on the polymer granules from a fixed-bed reactor for the denitrification of drinking water, a shower hose, water from the hydrotherapy pool of a large hospital at high counts, in hand bags and coats of hospital workers, in dental water reservoirs, gastric mucosa of dogs, bottled mineral water stored in returnable glass bottles and in polyvinyl chloride bottles, in fresh water habitats, a fish farm, conjunctival swabs, cervical cultures and also from soil (Aspinall and Graham, 1989). Hence, the source of B. vesicularis in marine system may be from fresh water run off. Occurrence of B. vesicularis in marine environment and its ability to degrade nylon was also reported (Sudhakar et al., 2007). Bacillus vesicularis has shown its ability to produce Poly hydroxy alkanoates, whose physical and chemical properties are similar to those of petroleum-derived plastics (Silva et al., 2007). The occurrence of B. vesicularis in sponges Craniella australiensis, Halichondria rugosa and Dysidea avara has been reported in China sea (Li et al., 2007). However, the occurrence of Bacillus vesicularis in the sponge Hyattella cribriformis has not been reported earlier and it may be the first research report.
Bacillus subtilis, B. firmus and Planococcus maritimus represent the phylum Firmicutes in the present study. P. maritimus was for the first time reported from the tidal flat of Korea by Yoon et al. (2003). Planococcus maritimus is gram-negative, motile cocci requiring 2% of NaCl for its growth (Yoon et al., 2003). This bacterium was explored commercially for its anti-oxidative, anti-cancer and anti-inflammatory bioactives (Krishnaveni and Jayachandran, 2009). The bacterium was also reported to occur in marine sediments at the depth of 1000 m in Andaman Sea, India (Krishnaveni and Jayachandran, 2009). In the present study, this bacterium was retrieved from the sponge collected at the depth of 5-10 m. This shows its facultative barophilic nature. Bacillus subtilis, known as the hay bacillus or grass bacillus, is a gram-positive, catalase-positive, rod-shaped bacterium commonly found in soil. Ability to form a tough, protective endospore, allowing the organism to tolerate extreme environmental conditions is a prime characteristic feature of this bacterium (Nakano and Zuber, 1998). The strain is considered to be commercially sound. With the development of genetic engineering, Bacillus subtilis is becoming a more attractive host to molecular biologists. The advantages of B. subtilis such are fast growth, ease in culturing, high secretion capacity and non-pathogenic GRAS (Generally Regarded As Safe) status for non-antibiotic producing strains. These traits have made it an attractive host for the production of heterologous enzymes (Deboer and Diderichsen, 1991). Preparing competent cells in B. subtilis was easier and effective.
About 58% (2379 genes) of B. subtilis contains protein-coding genes. Many hydrolytic enzymes such as amylase and protease, thermostable lipases have been commercially extracted from B. subtilis (Ahmed et al., 2009). Productions of lipopeptide-type biosurfactant, surfactin and rapid degradation of alkanes have also reported (Kim et al., 2000). The absence of an outer membrane in B. subtilis can simplify the protein secretion pathways and allow the organism to secrete high levels of extracellular proteins. Bacillus subtilis is also known to produce subtilin, subtilosin, rhizocticin two lipopeptides, surfactin and mycosubtilin and biopesticide (Kinsella et al., 2009).
Bacillus subtilis were shown to accumulate biologically active phenolic substances from culture liquid. These cells grown under micro-aerobic conditions accumulate up to 80-fold more copper than aerobically grown cells and maintain partial pH homeostasis during growth and withstand high concentrations of permanent acid stress (Dura et al., 2008; Kitko et al., 2009). B. subtilis possess features such as ability to survive in human gastrointestinal tract which could be advantageously used. This included the ability to form biofilms, to sporulate anaerobically and secretion of antimicrobials. B. subtilis probably got adapted for life within the gastrointestinal tract and should be considered as gut commensals rather than solely soil microorganism (Hong et al., 2009). It is evident from the present study that B. subtilis occurs both as symbiotic and free living form in the Ocean. This may be due to anthropogenic contamination via river run off.
Bacillus firmus is a facultative aerobic, alkaliphilic gram positive bacterium. It is commonly found in soil and known to fix nitrogen. B. firmus produces a strong acidic polysaccharide which could biosorbes Pb, Cu and Zn and could also act as bioflocculants (Salehizadeh and Shojaosadati, 2002). The strain was commercially exploited for its thermostable xylanase enzyme (Chang et al., 2004). It is also a nonpathogenic bacterium. Coupled with ovalbumin it was used as an antigen carrier in immunization experiments. B. firmus is a potential mucosal adjuvant because of its remarkable immunomodulatory properties. It has an ability to activate macrophages in vivo and in vitro accompanied by stimulation of NO synthase (Zidek et al., 1998). Cyclodextrin- glycosyltransferase recovered from this species is used in food, pharmaceutical and chemical industries as well as agriculture and environmental engineering (Matiol et al., 2000).
CONCLUSION
In the present study cultivable bacterial species associated with sponge Hyattella cribriformis have been isolated and precisely identified. The cultivable bacterial density of Hyattella cribriformis was found to be 65x10-5 CFU mL-1. The dominant Proteobacterial members in the community, ubiquitousness of Brevundimonas vesicularis and commercial probabilities of Firmicutes have been reviewed. The 16S rRNA sequences of the bacterial isolates can be accessed in NCBI’s accession numbers FJ834327, FJ834328, FJ834329, FJ845392, FJ845393, FJ845394 and FJ845395. Exploring the other commercial possibilities of the isolated strains will be the next priority.
ACKNOWLEDGMENTS
Authors are thankful to Prof. T. Balasubramanian, Dean, Faculty of Marine Science for the encouragement and the university authorities for the facilities. Authors thank Bioserve Biotechnologies, Pvt. Ltd. for technical support provided. The first author thanks UGC for financial support to carry out this work.
REFERENCES
- Beer, S. and M. Ilan, 1998. In Situ measurement of photosynthetic irradiance responses of two red sea sponges growing under dim light condition. Mar. Biol., 131: 613-617.
Direct Link - Chang, P., W.S. Tsai, C.L. Tsai and M.J. Tseng, 2004. Cloning and characterization of two thermostable xylanases from an alkaliphilic Bacillus firmus. Biochem. Biophys. Res. Commun., 319: 1017-1025.
PubMed - Chi, C.Y., C.P. Fung, W.W. Wong and C.Y. Liu, 2004. Brevundimonas bacteremia: Two case reports and literature review. Scand. J. Infect. Dis., 36: 59-61.
PubMed - Chomczynski, P. and N. Sacchi, 2006. The single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction: Twenty-something years on. Nat. Protocols, 1: 581-585.
CrossRefPubMedDirect Link - De Boer, A.S. and B. Diderichsen, 1991. On the safety of Bacillus subtilis and B. amyloliquefaciens: A review. Applied Microbiol. Biotechnol., 36: 1-4.
CrossRefPubMedDirect Link - Golyshin, P.N., T.N. Chernikova, W.R. Abraham, H. Lunsdorf, K.N. Timmis and M.M. Yakimov, 2002. Oleiphilaceae fam. nov., to include Oleiphilus messinensis gen. nov., sp. nov., a novel marine bacterium that obligately utilizes hydrocarbons. Int. J. Syst. Evol. Microbiol., 52: 901-911.
Direct Link - Guerinot, M.L., P.A. West, J.V. Lee and R.R. Colwell, 1982. Vibrio diazotrophicus sp. nov., a marine nitrogen-fixing bacterium. Int. J. Syst. Bacteriol. 32: 350-357.
Direct Link - Han, X.Y. and R.A. Andrade, 2005. Brevundimonas diminuta infections and its resistance to fluoroquinolones. J. Antimicrob. Chemother., 55: 853-859.
CrossRefDirect Link - Hill, M., A. Hill, N. Lopez and O. Harriott, 2006. Sponge-specific bacterial symbiont in the Caribbean sponge, Chondrilla nucula (Desmospongiae, Chondrosida). Mar. Biol., 148: 1221-1230.
Direct Link - Hong, H.A., R. Khaneja, N.M.K. Tam, A. Cazzato and S. Tan et al., 2009. Bacillus subtilis isolated from the human gastrointestinal tract. Res. Microbiol., 160: 134-143.
CrossRefPubMedDirect Link - Jean, W.D., W.Y. Shieh and T.Y. Liu, 2006. Thalassomonas agarivorans sp. nov., a marine agarolytic bacterium isolated from shallow coastal water of An-Ping Harbour, Taiwan and emended description of the genus Thalassomonas. Int. J. Syst. Evol. Microbiol., 56: 1245-1250.
CrossRef - Kawamura, Y., Y. Li, H. Liu, X. Huang and Z. Li, 2001. Bacterial population in Russian space station Mir. Microbiol. Immunol., 45: 819-828.
PubMed - Kim, H.S., S.B. Kim, S.H. Park, H.M. Oh and Y. Park et al., 2000. Expression of sfp gene and hydrocarbon degradation by Bacillus subtilis. Biotechnol. Lett., 22: 1431-1436.
CrossRef - Kinsella, K., C.P. Schulthess, T.F. Morris and J.D. Stuart, 2009. Rapid quantification of Bacillus subtilis antibiotics in the rhizosphere. Soil Biol. Biochem., 41: 374-379.
CrossRefDirect Link - Lau, W.W.Y., P.A. Jumars and E.V. Armburst, 2002. Genetic diversity of attached bacteria in the hindgut of the deposit-feeding shrimp neotrypaea (formerly Callianassa) californiensis (Decapoda: Thalassinidae). Microb. Ecol., 43: 455-466.
CrossRef - Li, Z., Y. Hu, Y.Q. Huang and Y. Huang, 2007. Isolation and phylogenetic analysis of the biologically active bacteria associated with three south China sea sponges. Microbiology, 76: 494-499.
CrossRef - Matiol, G., G. Zanin and F. Moraes, 2000. Enhancement of selectivity for producing γ-cyclodextrin. Applied Biochem. Biotech., 84-86: 955-962.
PubMed - Mondello, P., L. Ferrari and G. Carnevale, 2006. Nosocomial Brevundimonas vesicularis. Meningitis. Infez. Med., 14: 235-237.
PubMed - Nakano, M.M. and P. Zuber, 1998. Anaerobic growth of a strict aerobe (Bacillus subtilis). Annu. Rev. Microbiol., 52: 165-190.
PubMed - Ottaviani, D., I. Bacchiocchi, L. Masini, F. Leoni, A. Carraturo, M. Giammarioli and G. Sbaraglia, 2001. Antimicrobial susceptibility of potentially pathogenic halophilic vibrios isolated from seafood. Int. J. Antimicrobial Agents, 18: 135-140.
CrossRef - Salehizadeh, H. and S.A. Shojaosadati, 2002. Isolation and characterisation of a bioflocculant produced by Bacillus firmus. Biotech. Lett., 24: 35-40.
CrossRef - Segers, P., M. Vancanneyt, B. Pot, U. Torck and B. Hoste et al., 1994. Classification of Pseudomonas diminuta leifson and hugh 1954 and Pseudomonas vesicularis busing, doll and freytag 1953 in Brevundimonas gen. nov. as Brevundimonas diminuta comb. nov. and Brevundimonas vesicularis comb. nov., Respectively. Int. J. Syst. Bacteriol., 44: 499-510.
Direct Link - Selvin, J. and A.P. Lipton, 2004. Biopotentials of secondary metabolites isolated from marine spoges. Hydrobiologia, 513: 231-238.
Direct Link - Silva, J.A., L.M. Tobella, J. Becerra, F Godoy and M.A. Martinez, 2007. Biosynthesis of poly-β-hydroxyalkanoate by Brevundimonas vesicularis LMG P-23615 and Sphingopyxis macrogoltabida LMG 17324 using acid-hydrolyzed sawdust as carbon source. J. Biosci. Bioeng., 103: 542-546.
CrossRefDirect Link - Sudhakar, M., C. Priyadarshini, M. Doble, P.S. Murthy and R. Venkatesan, 2007. Marine bacteria mediated degradation of nylon 66 and 6. Int. Biodeter. Biodegra., 60: 144-151.
CrossRef - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Taylor, M.W., R. Radax, D. Steger and M. Wagner, 2007. Sponge-associated microorganisms: Evolution, ecology and biotechnological potential. Microbiol. Mol. Biol. Rev., 71: 295-347.
CrossRef - Tringe, S.G., T. Zhang, X. Liu, Y. Yu and W.H. Lee et al., 2008. The airborne metagenome in an indoor urban environment. PLoS One, Vol. 3.
CrossRefDirect Link - Vacelet, J., N. Boury-Esnault, A. Fial-medioni and C.R. Fisher, 1995. A methonotrophic carnivorous sponge. Nature, 377: 296-296.
CrossRef - Zidek, Z., L. Tuckova, M. Mara, R. Barot-Ciorbaru, L. Prokesova and H. Tlaskalova-Hogenova, 1998. Stimulation of macrophages by Bacillus firmus: Production of nitric oxide and cytokines. Int. J. Immunopharmacol., 20: 359-368.
PubMedDirect Link