
H.H. Azzaz
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
H.A. Murad
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
A.M. Kholif
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
T.A. Morsy
Department of Dairy Science, National Research Center, Dokki, Giza, Egypt
A.M. Mansour
Department of Animal Production, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
H.M. El-Sayed
Department of Animal Production, Faculty of Agriculture, Ain Shams University, Cairo, Egypt
Journal of Biological Sciences
Year: 2013 | Volume: 13 | Issue: 4 | Page No.: 234-241







ABSTRACT
Two experiments were carried out to evaluate the effects of fibrolytic enzymes supplementation on in vitro degradation of sugar beet pulp and in vivo nutrients digestibility, milk yield and its composition by mild-lactating buffaloes. In the in vitro experiment, dry matter and organic matter disappearance (IVDMD and IVOMD) were determined for sugar beet pulp supplemented separately with laboratory produced fibrolytic enzymes (Asperozym) and commercial fibrolytic enzymes source (Tomoko®) at 3 levels (0, 1, 1.5 and 2 g kg-1 DM). Increasing the Asperozym and Tomoko® supplementation levels up to 2 g kg-1 DM exhibited the highest (p<0.05) values of IVDMD and IVOMD. In the in vivo experiment, fifteen mild-lactating buffaloes after 3 months of parturition were divided into three groups, five animals each, using complete random design. The first group was fed on 45% Concentrate Feed Mixture (CFM), 30% corn silage ,15% dried sugar beet pulp and 10% rice straw (control ration). The second group was fed control ration supplemented with Asperozym at 2 g kg-1 DM (R1), while the third group was fed control ration supplemented with Tomoko® at 2 g kg-1 DM. (R2). Asperozym and Tomoko® supplementation significantly (p<0.05) increased DM, OM, CF, NFE, NDF digestibility for treated groups compared with the control group, while blood plasma parameter , milk yield and its composition did not significantly (p<0.05) change among all groups.
PDF Abstract XML References Citation
Received: March 13, 2013;
Accepted: April 10, 2013;
Published: July 10, 2013
How to cite this article
H.H. Azzaz, H.A. Murad, A.M. Kholif, T.A. Morsy, A.M. Mansour and H.M. El-Sayed, 2013. Increasing Nutrients Bioavailability by Using Fibrolytic Enzymes in Dairy Buffaloes Feeding. Journal of Biological Sciences, 13: 234-241.
DOI: 10.3923/jbs.2013.234.241
URL: https://scialert.net/abstract/?doi=jbs.2013.234.241
DOI: 10.3923/jbs.2013.234.241
URL: https://scialert.net/abstract/?doi=jbs.2013.234.241
INTRODUCTION
The increasing demand for milk and input costs in the dairy industry requires integrated strategies to increase production efficiency. One way of increasing production efficiency would be to increase nutrients bioavailability of feedstuffs. Yang et al. (2000) reported that, digestion of lingo-cellulosic materials in the rumen is insufficient and can negatively influence animal performance and increase dairy production cost. As a result, many strategies have been developed to stimulate the digestion of these lingo-cellulosic materials in ruminant feeds. The soundest strategy nowadays is using fibrolytic enzymes preparations as a driver for specific digestive and metabolic processes in the rumen which may be maximize natural digestive processes and increase nutrient bioavailability.
Many researchers have demonstrated that supplementing diets of dairy animals with fibrolytic enzymes can improve feed utilization and animal performance by enhancing fiber degradation in vitro (Gado et al., 2007; Rodrigues et al., 2008; Azzaz et al., 2012) and in situ (Tricarico et al., 2005; Krueger et al., 2008). A number of studies (Gado et al., 2007, 2009; Kholif et al., 2012) have shown that using fibrolytic enzyme supplements in dairy animal’s diets can increase milk production, while others (Bowman et al., 2002; Reddish and Kung, 2007) reported no effects on milk production. The discrepancies between the studies cited above are due to factors such as lactation period, dietary components, forage to concentrate ratio, enzyme composition, enzyme level and method of application.
This study was carried out to:
| • | Investigate the effects of laboratory produced fibrolytic enzymes (Asperozym) on banana fiber degradation by using electronic microscope |
| • | Evaluate the potential using of Asperozym for improving digestibility of sugar beet pulp compared (in- vitro) with commercial fibrolytic enzymes source (Tomoko®) |
| • | Investigate the impact of adding Asperozym and Tomoko® to rations of mid-lactating buffaloes on nutrients digestibility, blood parameters, milk yield and its composition |
MATERIALS AND METHODS
Sugar beet pulp: Beet pulp is the solid residue after extracting sugar from sugar beets; it was dried and pelleted for using it as animal feed. It was obtained from the Delta Company of sugar industry in Khafr El-Seikh province.
Enzymes sources
Tomoko®: A commercial enzymes source of Biogenkoji Research Institute - Japan. The enzyme product was made from Aspergillus Awamori (3 million cells/g) including 1000 unit g-1 of acidic protease, 30 unit g-1 of pectinase, 25 unit g-1 of xylanase, 20 unit g-1 of α-amylase, 10 unit g-1 of phytase, 5 unit g-1 of glucoamylase and 4 unit g-1 of cellulase.
Asperozym: Laboratory produced fibrolytic enzymes from Aspergillus niger. Each gram contains 30 units of pectinase and 4 unit of cellulase.
Enzymes assay: The pectinase and carboxymethyl-cellulase activities (CMC) for resultant crude enzyme extract (Asperozym) and commercial enzymes source (Tomoko®) were determined according to Buga et al. (2010) and Mandels et al .(1974), respectively. One unit of pectinase activity was defined as the amount of enzyme that produced one μmole of D-galacturonic acid per minute at 40°C and pH 5.0 (Soares et al., 1999), while One cellulase unit was defined as the amount of enzyme that liberates reducing sugar at the rate of 1 μmol mL-1 min-1 under assay condition (Miller, 1972).
Enzymatic treatment of banana fiber: Banana fibers were collected by hand stripping, dried and then treated with Asperozym (60 U pectinase and 8 U cellulase /1 g DM of banana fiber/1000 mL-1 buffer acetate at pH 6.5) for 24 h at 40°C and 100 rpm in a rotary shaker. The enzymatic untreated banana fiber (control) was kept in flask at the same condition of treated banana fiber with no enzyme addition. Finally, scanning electron micrographs of both treated and untreated fibers were taken to observe the effect of enzyme treatment.
In vitro study: Thirty five incubation flasks (250 mL-1 volume) were used to determine the in vitro dry matter and organic matter disappearance (IVDMD and IVOMD) for dried sugar beet pulp. Samples of 1 g of dried sugar beet pulp powder were accurately weighed into each flask. These flasks were separately supplemented with solution of Asperozym and Tomoko (5 flasks per each enzyme level) at different levels (0, 1, 1.5 and 2 g kg-1 DM). The in-vitro technique was carried out according to Fondevila and Perez-Espes, 2008. The procedures were using flasks filled with 140 mL of incubation solution prepared under a CO2 atmosphere, including a buffer solution, macro-mineral and trace mineral solution, a reduction solution and rumen inoculum. Rumen fluid was obtained from rams fed berseem hay ration using stomach tube. Whole rumen contents were obtained before morning feeding squeezed through four layers of gauze and fluid was collected in a pre-warmed thermos flask. Flasks were sealed and maintained at 39°C in a shaking water bath (20 oscillations/ min) for 48 h.
Digestibility and Lactating trails
Experimental animals: Fifteen mid lactation buffaloes at third to fifth seasons of lactation and weighed on average (620 kg) were used in the present study. Buffaloes were randomly divided after 3 months of parturition into three groups of five animals each using complete random design. The entire experimental period was 63 days (9 weeks). The first 3 weeks were considered as adaptation period on the new rations. Milk samples were taken after this adaptation period each week up to the end of experimental period. Blood, feed and feces samples were taken twice, one after one month of the beginning of the experimental period and the second by the end of experimental period.
Experimental rations: Buffaloes were individually fed according to 3% of body weight changed continuously according to animal weight changes. The first group of buffaloes were fed on 45% Concentrate Feed Mixture (CFM), 30% corn silage ,15% dried sugar beet pulp and 10% rice straw (control ration). The second group was fed control ration supplemented with Asperozym at 2 g kg-1 DM (R1), while the third group was fed control ration supplemented with Tomoko® at 2 g kg-1 DM. (R2). The concentrate feed mixture consisted of 45% yellow corn, 20% soybean meal , 15% wheat bran, 15% Distillers Dried Grains with Soluble (DDGS) 3% limestone, 1% minerals and 1% NaCl. Chemical compositions of CFM, corn silage, dried sugar beet pulp and rice straw are shown in Table 1.
Feeding management: The concentrate feed mixture was offered twice daily at 8.00 a.m. and 4:00 p.m., corn silage and dried sugar beet pulp were offered at 9:00 a.m., while rice straw was offered at 5:00 p.m. The enzymes were introduced twice day to each animal of second and third group with the concentrate feed mixture. Fresh water was available to the animals all the time.
| Table 1: | Chemical composition of feed ingredients (on DM basis) |
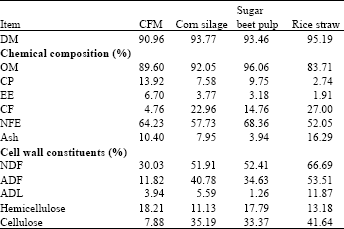 | |
| Hemicellulose: NDF-ADF, Cellulose: ADF-ADL, CFM: Concentrate feed mixture | |
Determination of digestion coefficients: Silica was used as an internal marker for determining the digestibility as described by Ferret et al. (1999). Fecal samples were collected from rectum of each animal by hand at 12:00 p.m. (after the distribution of morning CFM) for two successive days twice, the first one in days 31th-32th of the beginning of the experimental period and the second in days 61th-62th of the beginning of the experimental period. The collected feces were dried at 70°C for 48 h and then ground to pass a 1 mm sieve in a feed mill (FZ102, Shanghai Hong Ji instrument Co., Ltd., Shanghai, China) for chemical analysis. The digestibility coefficient of nutrient was calculated according to the following formula.
where, Dc is Digestion co-efficient, Sf is Silica in feed, Sc is Silica in feces, Nc is Nutrient in feces and Nf is Nutrient in feed.
Feed and fecal analysis: Feedstuffs and fecal samples were analyzed according to the AOAC (1995) methods to Determine Dry Matter (DM), Crude Protein (CP), Ether Extract (EE), Crude Fiber (CF) and ash contents. Organic matter (OM) and Nitrogen Free Extract (NFE) contents were calculated by difference. The Neutral Detergent Fiber (NDF), Acid Detergent Fiber (ADF) and Acid Detergent Lignin (ADL) contents were determined using the methods described by Van Soest et al. (1991).
Sampling and analysis of blood plasma: Blood samples were collected from the jugular vein of each animal at 12:00 pm (4 h after the distribution of morning CFM) for one successive day twice, the first one in 33th day of the beginning of the experimental period and the second in 63th day of the beginning of the experimental period. They were centrifuged at 4000 r.p.m. 20 min-1. The plasma was stored at -18°C till analysis. Plasma was collected and plasma total protein was determined as described by Armstrong and Carr (1964), albumin (Doumas et al., 1971), urea (Fawcett and Soctt, 1960), glucose (Siest et al., 1981) and plasma Aspartate aminotransferase (AST) and Alanin aminotransferase (ALT) (Reitman and Frankel, 1957). Globulin and albumin/globulin ratio were calculated.
Sampling and analysis of milk: Buffaloes were machine milked twice a day at 7:00 am and 7:00 pm. Milk samples were taken after the end of adaptation period during the last three days of each week up to the end of experimental period. Samples of milk were collected immediately from each animal after morning and evening milking and milk yield was recorded. The sample of each animal represented a mixed sample of constant percentage of the evening and morning yield. Milk samples were analyzed for total solids, fat, true protein and lactose by infrared spectrophotometry (Foss 120 Milko-Scan, Foss Q3 183 Electric, Hillerød, Denmark) according to AOAC (1995) procedures. Solids-not-fat (SNF) was calculated. Fat corrected milk (4% fat) was calculated by using the following equation:
FCM = 0.4 M+15 F |
where, M is milk yield (g) and F is fat yield (g).
Statistical analysis: Data obtained from this study were statistically analyzed by IBM (2011) using the following general model procedure:
Yij = μ+Ti+eij |
where, Yij is the parameter under analysis of the ij flask of laboratory trails or buffalo of farm trails, μ is the overall mean, Ti is the effect due to treatment on the parameter under analysis, eij is the experimental error for ij on the observation, the Duncan's multiple range tests was used to test the significance among means (Duncan, 1955).
RESULTS AND DISCUSSION
Enzymatic treatment of banana fibers: In higher plant tissues the cells are united together through the middle lamella which consists of mainly xylan and pectin (Jacob et al., 2008).
 | |
| Fig. 1(a-b): | Scanning electron micrographs of banana fibers. Scanning electron micrographs of banana fibers were taken (a) Before and (b) After treatment with Asperozym |
| Table 2: | Fibrolytic enzymes effect on dry matter and organic matter disappearance (in vitro) of dried sugar beet pulp |
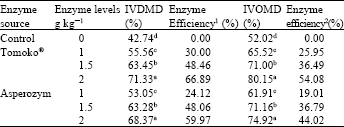 | |
| Enzyme efficiency % (DM): IVDMD% (sample)-IVDMD (%) (control)/IVDMD (%) (control)*100, Enzyme efficiency % (OM): IVOMD (%) (sample)-IVOMD (%) (control)/IVOMD (%) (control)*100, Each value of means obtained from five samples, a, b, c and d with different superscripts in the same column are significantly different (p<0.05) | |
It was evident from the photographs that the middle lamella of banana fibers was separated after treatment with Asperozym as a result of pectin hydrolysis (Fig. 1).
In vitro study: All levels of Asperozym and Tomoko® increased (p<0.05) DM and OM disappearance of dried sugar beet pulp compared with the untreated dried sugar beet pulp (Control), which gave the lowest values of IVDMD and IVOMD (Table 2). Increasing the Asperozym and Tomoko® supplementation levels up to 2 g kg-1 DM gave the highest values of both IVDMD and IVOMD of dried sugar beet pulp. Positive effects of fibrolytic enzymes on nutrients digestibility have been reported in different in vitro studies (Gado et al., 2007; Rodrigues et al., 2008; Azzaz et al., 2012).
| Table 3: | Fibrolytic enzymes effects on digestion coefficients and nutritive values of experimental rations fed to buffaloes |
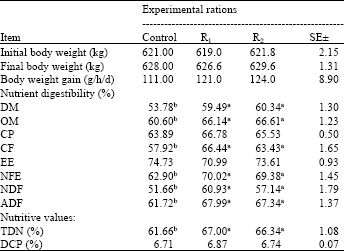 | |
| Means designated with the same letter in the same row are not significantly different at 0.05 level of probability, SE: Standard error | |
Dong et al. (1999) demonstrated that the effects of enzymes might start when the enzyme is in contact with the substrate, so enzyme-feed interaction appears as important. The enzyme addition onto feeds may create a stable enzyme-feed complex that protects free enzymes from proteolysis by the rumen microorganisms as reported by Kung et al. (2000).
Till now, little is known about the way that exogenous fibrolytic enzymes improve feed digestibility in ruminates. Several potential modes of action have been proposed. These include: (1) Increase in microbial colonization of feed particles (Yang et al., 1999), (2) Enhancing attachment and /or improve access to the cell wall matrix by ruminal microorganisms and by doing so, accelerate the rate of digestion (Nsereko et al., 2000) and (3) Enhancing the hydrolytic ability of the ruminal microorganisms due to added enzyme activities and /or synergy with rumen microbial enzymes (Morgavi et al., 2000).
In the present study Asperozym and Tomoko® might have the ability to degrade complex substrates (cellulose and pectin) to simpler ones which might have altered the structure of dried sugar beet pulp making them more amenable to ruminal microorganisms and allowing a faster ruminal microbial colonization and fermentation.
Digestibility and nutritive values: Data of Table 3 clearly show that both of rations supplemented with Asperozym (R1) and Tomoko® (R2) significantly (p<0.05) increased DM, OM, CF, NFE, NDF and ADF digestibility compared with the control ration. No significant (p>0.05) differences were detected in CP and EE digestibility among fibrolytic enzymes treated or untreated buffaloes. These findings are in line with other studies which reported increase in total tract digestibility of DM, OM and fiber fraction digestibility, following treatment with fibrolytic enzymes (Gado et al., 2009; Azzaz et al., 2012; Kholif et al., 2012). Exogenous fibrolytic enzymes would be expected to increase total tract digestibility by increasing the rate of ruminal digestion of the potentially digestible NDF fraction (Yang et al., 1999), but increases in total tract digestibility may also be, in part, due to reduced digesta viscosity (Hristov et al., 2000), alterations in ruminal fermentation (Nsereko et al., 2002) and/or enhanced attachment and colonization to the plant cell wall by ruminal microorganisms (Nsereko et al., 2000) and/or by synergism with enzymes in rumen fluid (Morgavi et al., 2000).
The nutritive values of the experimental rations expressed as total digestible nutrients (TDN) and digestible crude protein (DCP) are shown in Table 3. Digestible crude protein (DCP) was not affected by fibrolytic enzymes treatments. This may be due to less solubility of corn protein (zein) as reported by Nolan, (1976), zein consist most of ration’s protein fed to buffaloes in this study. These results are in line with those obtained by Knowlton et al. (2002) and Muwalla et al. 2007. They mentioned that apparent protein digestibility was not significantly affected by fibrolytic enzymes treatment. On the other hand, buffaloes fed on fibrolytic enzymes treated rations showed significant (p<0.05) higher TDN compared to those fed the control ration. This may be due to large amount of readily fermentable carbohydrate which liberated due to action of fibrolytic enzymes on cellulose and pectin of rations fed to buffaloes. This result is in line with those obtained by Azzaz et al. (2012) and Kholif et al. (2012).
Blood plasma parameters: Effect of fibrolytic enzymes (Asperozym and Tomoko®) treatments on plasma total protein, albumin, globulin concentration and albumin/ globulin ratio of mild lactating buffaloes received the different experimental rations are shown in Table 4.
| Table 4: | Blood plasma parameters of lactating buffaloes fed the different experimental ratios |
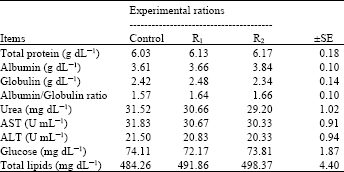 | |
| SE: Standard error | |
Fibrolytic enzymes supplemented rations were not significantly affected plasma total protein, albumin, globulin concentrations and albumin/globulin ratio. This may be due to insignificantly differences among fibrolytic enzymes treated or untreated (control) buffaloes in protein’s rations digestibility. Our results are in line with those obtained by Kholif et al. (2012). Who stated that exogenous enzymes supplemented rations were not significantly affected buffalo’s blood serum total protein, albumin, globulin and albumin/globulin ratio.
Data of Table 4 showed that blood plasma urea, AST and ALT values were not affected by fibrolytic enzymes treatments. These results are in line with those obtained by Kholif, (2006). Who found that animals fed on fibrolytic enzymes or fungi treated silage had no significant change in blood serum urea, GOT and GPT concentrations compared with animals fed on untreated silage. Also, Kholif et al. (2012) found that exogenous enzymes supplemented rations were not significantly affected buffalo’s blood serum urea, AST and ALT values.
There were insignificant differences (p>0.05) among all groups in the overall means of plasma glucose. These results are in line with those obtained by Kholif (2006) who found that animals fed on fibrolytic enzymes or fungi treated silage had no significant increase in serum glucose concentration. Also, Azzaz et al. (2012) found that cellulases addition to diets of lactating goats was not significantly affected plasma glucose concentration. Also, there were insignificant differences (p>0.05) among all groups in the overall means of plasma total lipids but plasma total lipids showed slightly increase by buffaloes fed rations supplemented with fibrolytic enzymes compared with control buffaloes. These results are in line with those obtained by Kholif (2006) who found that animals fed on fibrolytic enzymes or fungi treated silage had no significant increase in serum total lipids concentration. In addition, Azzaz et al. (2012) found that cellulases addition to diets of lactating goats was not significantly affected plasma total lipids concentration. Also, Kholif et al. (2012) found that exogenous enzymes supplemented rations were not significantly affected buffalo’s serum total lipids concentration.
Finally, the concentrations of plasma proteins, glucose, total lipids, AST and ALT were not significantly affected by treatments and in the normal range for healthy animals. These results indicated that tested additives to lactating buffalo’s rations were not negatively affected liver activity or general animal’s health.
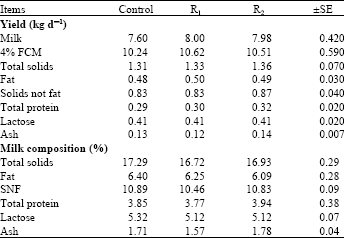
| Table 5: | Fibrolytic enzymes effects on buffalo’s milk yield and its composition |
 | |
| SE: Standard error | |
Milk yield and its composition: Data of Table 5 showed that there were no significant (p>0.05) differences among control and fibrolytic enzymes treated groups in milk composition and milk component’s yields. Adding Asperozym to lactating buffalo’s ration increased milk production by 5.3 and 4% fat corrected milk production by 3.71% , while adding Tomoko® to lactating buffalo’s ration increased milk production by 5 and 4% fat corrected milk production by 2.64% compared with control ration.
These results are in line with those obtained by Chen et al. (1995). who found that application of fibrolytic enzyme mixtures (DigestM®, Loveland Industries Inc., Greeley, CO, USA) onto sorghum grain did not improve milk production in Holstein cows. Similarly, addition of a nonstarch polysaccharidase to grains increased total tract digestibility in mid-lactation cows (Beauchemin et al., 2000) but no effect on milk yield was observed. Also (Bowman et al., 2002) reported that, despite the increase in total tract feed digestion, the response in milk production was not observed with fibrolytic enzymes supplementation to diets of dairy Holstein cows. Reddish and Kung (2007) reported that adding a dry enzyme mixture to lactating cow’s diets had no effect on milk production, or milk composition. Elwakeel et al. (2007) studied the impact of fibrolytic enzymes addition to cow’s diets in the presence and absence of supplemental yeast to determine whether the responses to enzymes might be influenced by yeast supplementation. The results indicated that milk production, milk efficiency and production of all milk components were not affected by the addition of either fibrolytic enzymes or yeast.
In this study, the specific reasons for a lack of the dry fibrolytic enzymes (Asperozym and Tomoko®) effects on performance of mild lactating buffaloes are unknown. The energy status of lactating animals which varied with stage of lactation changes may be made responses to fibrolytic enzymes addition to rations vary. This approach is supported by some studies, Schingoethe et al. (1999) reported that cows responded to enzyme-treated forages in early lactation, but responses were not apparent in later lactation. Zheng et al, 2000 observed that the effect of fibrolytic enzymes on milk production were more pronounced in early lactation period when cows were in negative energy balance while, (Beauchemin et al., 2000) observed that addition of fibrolytic enzymes to rations of cows which were in positive energy balance had no effect on milk yield and its composition. Knowlton et al. (2002) demonstrated that, the production responses to fibrolytic enzymes addition to diets were only in cows that began treatment during the first 100 day of lactation and milk yield was not affected in cows in mid-lactation at the start of treatment. Based on the previous studies, the positive energy status of our lactating buffaloes was attributed to the lack of response to Asperozym and Tomoko® treatment. On the other hand, we hypothesized that the increased amount of digestible energy due to addition of Asperozym and Tomoko® to rations which were not used for milk production were allocated to milk fat, milk protein and body reserves. This is supported by the numerical increase for change in body weight for buffaloes fed Asperozym and Tomoko® supplemented rations compared to buffaloes fed control ration (Table 3).
CONCLUSION
Finally, we can concluded that, although the evaluated fibrolytic enzymes (Asperozym and Tomoko®) were capable of increasing IVDMD, IVOMD of dried sugar beet pulp and most nutrient digestibility of treated rations, the enzymes did not significantly affect the lactational performance of dairy buffaloes. It’s possible that the dietary components to which the enzymes were added and lactation stage of buffaloes under study were limiting factors of their impact on lactation performance.
REFERENCES
- Azzaz, H.H., A.M. Kholif, H.A. Murad, M.A. Hanfy and M.H. Abdel Gawad, 2012. Utilization of cellulolytic enzymes to improve the nutritive value of banana wastes and performance of lactating goats. Asian J. Anim. Vet. Adv., 7: 664-673.
CrossRefDirect Link - Bowman, G.R., K.A. Beauchemin and J.A. Shelford, 2002. The proportion of the diet to which fibrolytic enzymes are added affects nutrient digestion by lactating dairy cows. J. Dairy Sci., 85: 3420-3429.
CrossRefDirect Link - Buga, M.L., S. Ibrahim and A.J. Nok, 2010. Partially purified polygalacturonase from Aspergillus niger (SA6). Afr. J. Biotechnol., 9: 8944-8954.
Direct Link - Chen, K.H., J.T. Huber, J. Simas, C.B. Theurer and P. Yu et al., 1995. Effect of enzyme treatment or steam-flaking of sorghum grain on lactation and digestion in dairy cows. J. Dairy Sci., 78: 1721-1727.
CrossRef - Dong, Y., H.D. Bae, T.A. McAllister, G.W. Mathison and K.J. Cheng, 1999. Effects of exogenous fibrolytic enzymes, α-bromoethanesulfonate and monensin on fermentation in a rumen simulation (RUSITEC) system. Can. J. Anim. Sci., 79: 491-498.
CrossRefDirect Link - Doumas, B.T., W.A. Watson and H.G. Biggs, 1971. Albumin standards and the measurement of serum albumin with bromcresol green. Clin. Chim. Acta, 31: 87-96.
CrossRefPubMedDirect Link - Elwakeel, E.A., E.C. Titgemeyer, B.J. Johnson, C.K. Armendariz and J.E. Shirley, 2007. Fibrolytic enzymes to increase the nutritive value of dairy feedstuffs. J. Dairy Sci., 90: 5226-5236.
CrossRefDirect Link - Ferret, A., J. Plaixats, G. Caja, J. Gasa and P. Prio, 1999. Using markers to estimate apparent dry matter digestibility, faecal output and dry matter intake in dairy ewes fed Italian ryegrass hay or alfalfa hay. Small Rumin. Res., 33: 145-152.
CrossRefDirect Link - Fondevila, M. and B. Perez-Espes, 2008. A new in vitro system to study the effect of liquid phase turnover and pH on microbial fermentation of concentrate diets for ruminants. Anim. Feed Sci. Technol., 144: 196-211.
CrossRefDirect Link - Gado, H.M., A.Z.M. Salem, P.H. Robinson and M. Hassan, 2009. Influence of exogenous enzymes on nutrient digestibility, extent of ruminal fermentation as well as milk production and composition in dairy cows. Anim. Feed Sci. Technol., 154: 36-46.
CrossRefDirect Link - Hristov, A.N., T.A. McAllister and K.J. Cheng, 2000. Intraruminal supplementation with increasing levels of exogenous polysaccharide-degrading enzymes: Effects on nutrient digestion in cattle fed a barley grain diet. J. Anim. Sci., 78: 477-487.
PubMedDirect Link - Jacob, N., K.N. Niladevi, G.S. Anisha and P. Prema, 2008. Hydrolysis of pectin: An enzymatic approach and its application in banana fiber processing. Microbiol. Res., 163: 538-544.
CrossRefPubMedDirect Link - Knowlton, K.F., J.M. McKinney and C. Cobb, 2002. Effect of a direct-fed fibrolytic enzyme formulation on nutrient intake, partitioning and excretion in early and late lactation Holstein cows. J. Dairy Sci., 85: 3328-3335.
CrossRefDirect Link - Krueger, N.A., A.T. Adesogan, C.R. Staples, W.K. Krueger, S.C. Kim, R.C. Littell and L.E. Sollenberger, 2008. Effect of method of applying fibrolytic enzymes or ammonia to Bermudagrass hay on feed intake, digestion and growth of beef steers. J. Anim. Sci., 86: 882-889.
CrossRefPubMedDirect Link - Kung, Jr. L., R.J. Treacher, G.A. Nauman, A.M. Smagala, K.M. Endres and M.A. Cohen, 2000. The effect of treating forages with fibrolytic enzymes on its nutritive value and lactation performance of dairy cows. J. Dairy Sci., 83: 115-122.
CrossRefDirect Link - Morgavi, D.P., K.A. Beauchemin, V.L. Nsereko, L.M. Rode and A.D. Iwaasa et al., 2000. Synergy between ruminal fibrolytic enzymes and enzymes from Trichoderma longibrachiatum. J. Dairy Sci., 83: 1310-1321.
CrossRefDirect Link - Muwalla, M.M., S.G. Haddad and M.A. Hijazeen, 2007. Effect of fibrolytic enzyme inclusion in high concentrate fattening diets on nutrient digestibility and growth performance of Awassi lambs. Livestock Sci., 111: 255-258.
CrossRef - Nsereko, V.L., K.A. Beauchemin, D.P. Morgavi, L.M. Rode and A.F. Furtado et al., 2002. Effect of a fibrolytic enzyme preparation from Trichoderma longibrachiatum on the rumen microbial population of dairy cows. Can. J. Microbiol., 48: 14-20.
PubMedDirect Link - Nsereko, V.L., D.P. Morgavi, L.M. Rode, K.A. Beauchemin and T.A. McAllister, 2000. Effects of fungal enzyme preparations on hydrolysis and subsequent degradation of alfalfa hay fiber by mixed rumen microorganisms in vitro. Anim. Feed Sci. Technol., 88: 153-170.
CrossRefDirect Link - Reddish, M.A. and L. Kung Jr., 2007. The effect of feeding a dry enzyme mixture with fibrolytic activity on the performance of lactating cows and digestibility of a diet for sheep. J. Dairy Sci., 90: 4724-4729.
CrossRefDirect Link - Rodrigues, M.A.M., P. Pinto, R.M.F. Bezerra, A.A. Dias and C.V.M. Guedes et al., 2008. Effect of enzyme extracts isolated from white-rot fungi on chemical composition and in vitro digestibility of wheat straw. Anim. Feed Sci. Technol., 141: 326-338.
CrossRefDirect Link - Soares, M.M.C.N., R. da Silva and E. Gomes, 1999. Screening of bacterial strains for pectinolytic activity characterization of the pgase produced by Bacillus species. Rev. Microbiol., 30: 299-303.
Direct Link - Tricarico, J.M., J.D. Johnston, K.A. Dawson, K.C. Hanson, K.R. McLeod and D.L. Harmon, 2005. The effects of an Aspergillus oryzae extract containing alpha-amylase activity on ruminal fermentation and milk production in lactating Holstein cows. Anim. Sci., 81: 365-374.
CrossRefDirect Link - van Soest, P.J., J.B. Robertson and B.A. Lewis, 1991. Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci., 74: 3583-3597.
CrossRefPubMedDirect Link - Yang, W.Z., K.A. Beauchemin and L.M. Rode, 1999. Effects of an enzyme feed additive on extent of digestion and milk production of lactating dairy cows. J. Dairy Sci., 82: 391-403.
CrossRefPubMedDirect Link - Yang, W.Z., K.A. Beauchemin and L.M. Rode, 2000. A comparison of methods of adding fibrolytic enzymes to lactating cow diets. J. Dairy Sci., 83: 2512-2520.
CrossRefPubMedDirect Link - Zheng, W., D.J. Schingoethe, G.A. Stegeman, A.R. Hippen and R.J. Treacher, 2000. Determination of when during the lactation cycle to start feeding a cellulase and xylanase enzyme mixture to dairy cows. J. Dairy Sci., 83: 2319-2325.
CrossRefDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Mandels, M., L. Hontz and J. Nystrom, 1974. Enzymatic hydrolysis of waste cellulose. Biotechnol. Bioeng., 16: 1471-1493.
CrossRefDirect Link - Schingoethe, D.J., G.A. Stegeman and R.J. Treacher, 1999. Response of lactating dairy cows to a cellulase and xylanase enzyme mixture applied to forages at the time of feeding. J. Dairy Sci., 82: 996-1003.
CrossRefDirect Link - Fawcett, J.K. and J.E. Scott, 1960. A rapid and precise method for the determination of urea. J. Clin. Pathol., 13: 156-159.
CrossRefPubMedDirect Link