
Wipawadee Budda
Protein and Enzyme Technology Research Unit and Center of Excellence for Innovation in Chemistry, Department of Chemistry, Faculty of Science, Mahasarakham University, Maha Sarakham, 44150, Thailand
Rakrudee Sarnthima
Protein and Enzyme Technology Research Unit and Center of Excellence for Innovation in Chemistry, Department of Chemistry, Faculty of Science, Mahasarakham University, Maha Sarakham, 44150, Thailand
Saranyu Khammuang
Protein and Enzyme Technology Research Unit and Center of Excellence for Innovation in Chemistry, Department of Chemistry, Faculty of Science, Mahasarakham University, Maha Sarakham, 44150, Thailand
Nipa Milintawisamai
Department of Biochemistry, Faculty of Science, Khon Kaen University, 40002, Thailand
Sawait Naknil
Phoenix Pulp and Paper Public Company Limited, Khon Kaen, 40310, Thailand
Journal of Biological Sciences
Year: 2012 | Volume: 12 | Issue: 1 | Page No.: 25-33







ABSTRACT
In nature, lignin can be degraded by ligninolytic enzymes from several of white rot fungi. Since, they can produce and secrete ligninolytic enzymes for lignin degradation and use as a carbon source. In this study, ligninolytic enzymes from Lentinus polychrous were investigated using CuSO4 at various concentrations (0-2.4 μmol g-1 solid substrate) as laccase inducer. It was found that ligninolytic enzymes production by the fungus showed similar pattern. The main enzyme activities detected was laccase, following with manganese independent peroxidase and manganese dependence peroxidase, respectively. No lignin peroxidase activity was detected. Carbohydrate degrading enzymes, β-glucosidase and cellulase activities have been found in a low quantity. Optimum conditions for catalysis were at pH 3-4 and temperature of 55°C. The crude laccase showed a good stability in distilled water as well as in buffers pH 6-8. The highest stability was at pH 7 and 30°C. The crude laccase of Lentinus polychrous was also evaluated for black liquor decolorization.
PDF Abstract XML References Citation
Received: August 04, 2011;
Accepted: October 21, 2011;
Published: January 20, 2012
How to cite this article
Wipawadee Budda, Rakrudee Sarnthima, Saranyu Khammuang, Nipa Milintawisamai and Sawait Naknil, 2012. Ligninolytic Enzymes of Lentinus polychrous Grown on Solid Substrates and its Application in Black Liquor Treatment. Journal of Biological Sciences, 12: 25-33.
DOI: 10.3923/jbs.2012.25.33
URL: https://scialert.net/abstract/?doi=jbs.2012.25.33
DOI: 10.3923/jbs.2012.25.33
URL: https://scialert.net/abstract/?doi=jbs.2012.25.33
INTRODUCTION
White rot fungi are well recognized as lignin degrader. These fungi secrete ligninolytic enzymes for hydrolyzing their substrates. Ligninolytic enzymes mainly contain lignin peroxidase, manganese-dependent peroxidase and laccase. Some strains of white rot fungi produce all three ligninolytic enzymes but some produce only one or two enzyme (Baldrian, 2006). Lignin peroxidase and manganese-dependent peroxidase require hydrogen peroxide as co-substrate but there is no need for laccase. Therefore, laccase is more practically used when it needs for applications. Laccase catalyzes the oxidation of several substrates including phenolic compounds, aromatic amines, thiols and some inorganic compounds using molecular oxygen as electron acceptor (Pezzella et al., 2009). The low substrate specificity makes this enzyme interesting for biotechnology purposes in various industries such as pulp and paper and textiles and bioremediation of industrial pollutants (Arora and Sharma, 2010). High amount ligninolytic enzymes productions in considerable low cost substrates are of interest. In addition researches have been investigated to find laccase with new properties or interesting properties for their applications. The whiterot fungus, Lentinus polychrous is edible mushroom which is popular to eat in Northeast and North of Thailand. In our laboratory, we have reported ligninolytic enzymes activity of crude extract of the fungus grown on Solid-state Fermentation (SSF) and their potential uses in dye decolorization (Khammuang and Sarnthima, 2007; Sarnthima et al., 2009). In this study, therefore, we extended investigated production of ligninolytic enzymes from Lentinus polychrous on SSF in the presence of copper(II)sulfate as laccase inducer as well as evaluated their ability in back liquor decolorization.
MATERIALS AND METHODS
Production and preparation of crude enzyme: Lentinus polychrous Lév. was kindly donated by Rujira mushroom farm, Ka La Sin province, Thailand. It was maintained at 4°C on Potato Dextrose Agar (PDA) plates and sub-cultured every 1-2 months. Ligninolytic enzymes were obtained by culturing the fungus on solid substrates of rice husk and bran as previously reported by Sarnthima et al. (2009). The laccase enzyme was induced by adding copper two ions (CuSO4 0.04-2.4 μmol g-1 solid substrate in distilled water) into the culture. The fungal cultures were performed at room temperature (28±2°C) for 5 weeks. The enzymes were extracted from the fungal culture media using distilled water in the extraction ratio of 1 g of the fungal culture media per 3 mL of distilled water. The extraction was stirred by a stirring rod manually for 10 min. The filtered through a double layer of cheesecloth was then centrifuged at 10,000 rpm for 20 min at 10°C to get a clear supernatant called the crude enzyme.
Protein determination and ligninolytic enzymes assay: Protein content in crude extract was determined by the method of Bradford (1976) using Bio-Rad protein assay reagent and Bovine Serum Albumin (BSA) as a protein standard.
Laccase (Lcc) activity was determined using 2,2-azinobis-(3-ethylbenzo-6-thiazoline-sulfonic acid) (ABTS) as a substrate. Briefly, the reaction was performed in 0.1 M sodium acetate buffer (pH 4.5) at 32°C for 10 min as previous described by Khammuang and Sarnthima (2007). One unit of laccase activity is the amounts of enzymes that oxidizes 1.0 μmole substrate and generate 1.0 μmole product per minute at assay condition.
Lignin peroxidase (LiP) activity was measured by the oxidation of Veratyl Alcohol (VA) in 0.1 M phosphate buffer, pH 6.5 in the presence of H2O2. After incubation at 32°C for 10 min, the absorbance was read at 310 nm (ε = 11.4x103 M-1 cm-1) or using the principle of Azure B decolourization (Sarnthima et al., 2009).
Manganese peroxidase (MnP) activity was measured as described previously (Sarnthima et al., 2009) using 3-methyl-2-benzithiazolinone hydrazone (MBTH) and dimethylaminobenzoic acid (DMAB) as substrates in 0.1 M sodium acetate buffer, pH 4.5 in the presence of H2O2 and Mn2+. The absorbance was read at 590 nm (ε = 5.3x103 M-1 cm-1).
Manganese-independent Peroxidase (MIP) activity was measured using the same method as the manganese peroxidase activity but adding EDTA for cheated ion of Mn2+. The real MnP activity was corrected by subtraction of MIP activity from peroxidase activity.
Carbohydrate degrading enzymes assay: The xylanase activity was determined by the method of Bailey et al. (1992). The substrate solution, 1% birchwood xylan solubilized in 0.5 M citrate-phosphate buffer (pH 6.0). The reaction mixture consisted of 1.8 mL substrate solution and 0.2 mL properly diluted enzyme. After 5 min of incubation at 50°C, the liberated reducing sugars (xylose equivalent) were estimated by the dinitrosalicylic acid method (DNS) according to Miller (1959). One unit of xylanase is defined as the amount of enzyme releasing 1 μmole of reducing sugar per minute per milliliter under the assay conditions described.
The cellulase activity was determined as Filter Paper Activity (FPA) according to Ghose (1987). Whatman No. 1 filter paper strip (1.0x6.0 cm) was incubated with 0.5 mL of crude extract at appropriate dilution in 0.05 M Na-citrate buffer (pH 4.8) and with 1 mL sodium citrate buffer. After 60 min of incubation at 50°C, the liberated reducing sugars (glucose equivalent) were estimated by the DNS method according to Miller (1959).
Beta-glucosidase activity was determined by the hydrolysis reaction of p-nitrophenyl-β-D-glucoside (Glcβ-O-pNP) in 0.1 M citric acid-0.2 M Na2HPO4 (McIlvaine buffer), pH 5 at 30°C and measured the release of p-nitrophenol (p-NP) spectrophotometrically at a wavelength of 400 nm (Srisomsap et al., 1996). One unit of Beta-glucosidase activity is defined as the amount of enzyme that releases 1 μmole of p-NP per minute.
Enzyme characteristics: Optimum pH and optimum temperature for catalysis of the crude enzyme were investigated using ABTS as laccase substrate. For pH optimum, the assay reactions were performed as previously described but in various pH conditions ranging from pH 2-9. For temperature optimum, the reactions were incubated at various temperatures ranging from 30-75°C.
The crude enzyme was studied pH stability by incubation in buffers different pH ranging from 3-8 including in distilled water at 32°C, aliquots were taken periodically for laccase activity assay. Temperature stability was done by incubating the crude enzyme in buffer pH 7.0 at various temperatures of 30, 40, 50 and 60°C, aliquots were withdrawn periodically for laccase activity assay.
Polyacrylamide Gel Electrophoresis (PAGE) was performed under nondenaturing conditions. The separating and stacking gels contained 12 and 4% acrylamide, respectively. The gel was run according to Ornstein (1964) and Davis (1964) at constant volts of 100 V per gel. The gels were stained to visualize ligninolytic enzymes activity by using ABTS and syringaldazine for laccase activity and MBTH+DMAB+ H2O2+Mn2+ for peroxidase activity. The gel was also stained with coomassie brilliant blue R-250 for protein distributions.
Black liquor decolorization: Biodegradation of black liquor by the crude enzyme from L. polychrous was also investigated in order to confirm enzymatic delignification. Degradation of by-products was measured by a spectrophotometric technique. The solutions were scanned over the wavelength ranging from 200 to 600 nm by a UV-visible spectrophotometer (Lara et al., 2003). Reaction mixtures (2 mL) contained 0.2 U mL-1 laccase activities, the suitable diluted black liquors and without or with some mediators in 0.1 M sodium acetate buffer (pH 4.5), were incubated at 32°C for 48 h. The reactions were periodically monitored the decrease in absorbance at the maximum absorption wavelength (λmax). For black liquors decolorization, these were used to calculate decolorization percentage according to the following equation as previously described by Khammuang and Sarnthima (2007).
Where, A0 is an absorbance at the maximum absorption wavelength of the black liquor immediately measured after adding the enzyme solution and At is an absorbance at the maximum absorption wavelength of the black liquor after each time intervals.
RESULTS AND DISCUSSION
Extracellular lignin degrading enzymes production: The fungus L. polychrous was cultured on solid substrate of rice husk and bran (1:2 by weight) in total about 45 g substrate containing 0, 0.04, 0.16, 0.8 and 2.4 μmole CuSO4 as laccase inducer. During cultured at room temperature the ligninolytic enzymes were extracted by distilled water periodically for 5 weeks. Then, the extract was centrifuged and the supernatant was measured for the total volume, protein contents, ligninolytic enzymes activities including laccase, MnP, MIP and LiP. The results showed in Fig. 1a-d.
The crude enzymes from without and with inducer CuSO4 (0.04-2.4 μmole CuSO4/g solid substrate) showed similar in the protein pattern (Fig. 1a) but clearly different in ligninolytic enzymes production (Fig. 1b-d). However, the fungus cultured in the presence of CuSO4 as inducer showed protein concentration slightly higher than those of the absence one at all of observed times (Fig. 1a).
For laccase activity assay, the result showed a rapid increasing of laccase activity in the week of 3-4 cultivations (21-28 days) and after that, it is static or slightly decreasing (Fig. 1b).
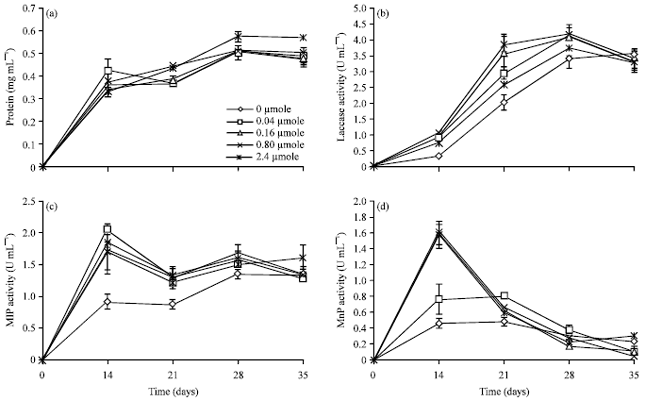 | |
| Fig. 1(a-d): | Protein content (a) Laccase, (b) MIP, (c) and MnP activity d) in the absence and presence of 0.04, 0.16, 0.8 and 2.4 μmole CuSO4 per gram solid substrate at different times of cultivation |
It was found that in the presence of CuSO4 inducer, laccase activity was increased to its maximum above 4 U mL-1 at CuSO4 concentration ranging from 0.04-0.8 μmole per gram solid substrate with laccase specific activity around 8 U mg-1. Culture of 2.4 μmole CuSO4 per gram solid substrate, laccase activity was lower. Therefore, L. polychrous culture in this rice husk and bran media in the presence of CuSO4 inducer between 0.04-0.8 μmole per gram solid substrate for 28 days seemed to be the most suitable condition for laccase production. At 35 days of cultivation, laccase activity from all cultures with CuSO4 induced was decreased, except culture of the fungus without inducer which activity was slightly increased (Fig. 1b). Copper(II)sulfate is most known laccase inducer of white rot fungi. This has been reported as a good inducer in many fungal strains and being involved with isoenzymes production (Palmieri et al., 2000; Levin et al., 2002; Cordi et al., 2007).
For lignin peroxidase activity assay, the result showed no activity of this enzyme in all of cultivation conditions when using VA or Azure B as substrates (assayed only in 28 days of cultivation conditions). Since, this fungus may not have gene of LiP enzyme or if any, it seemed not expressed. Some white rot fungi produce only laccase, no MnP and LiP for example laccase producing Pycnoporus cinnabarinus (Eggert et al., 1996; Geng and Li, 2002), laccase and MnP producing Ganoderma lucidum (D’Souza et al., 1999).
For manganese independent peroxidase activity assay, the result showed the highest MIP activity after 2 weeks of cultivation. After that, the activity was static or decreases (Fig. 1c). It was found that in the presence of CuSO4, MIP activity was higher than that of control. However, in the presence of CuSO4 (0.04-2.4 μmole g-1 solid substrate) showed no difference in MIP activity and found that at 0.04 μmole CuSO4 the highest MIP activity was observed. Figure 1d represents MnP activity increasing according to CuSO4 concentration. However, in the presence of CuSO4 (0.16-2.4 μmole g-1 solid substrate) showed no difference in MnP activity. When considered the MnP specific activity, it also showed consistent results (data not shown). The major ligninolytic enzymes produced by the fungus were laccase and followed with MIP and MnP, respectively with no LiP activity have been confirmed in this study. Similar observation in the culture on rice husk and bran without inducer previously reported (Sarnthima et al., 2009).
Extracellular carbohydrate degrading enzymes production: L. polychrous has not yet been reported whether it can produce and secrete carbohydrate-degrading enzymes, such as xylanase, cellulase and β-glucosidase. Thus, it is necessary to measure these enzyme activities if application of paper and pulp industry is in consideration which is because these enzymes have certain effects on pulp and paper quality. The results showed that the crude extract had no xylanase activity, while β-glucosidase and cellulase activities had been found in low quantities (Fig. 2a, b). The fungus gave very low cellulase activity when compared to T. versicolor, Phellinus sp., Daedalea sp. and P. coccineus (Liew et al., 2010). This low carbohydrate-degrading activity further suggests that this white rot strain is more interested applying for pulp biobleaching.
Take a close look at culture of 28 days; CuSO4 could induce ligninolytic enzymes activity as shown in Fig. 3a and carbohydrate degrading enzymes activity as shown in Fig. 3b. From the results, we chose 0.8 μmole CuSO4 per gram solid substrate for further experiment to produce a large quantity of laccase. This culture condition showed activity ratios of laccase to peroxidase, β-glucosidase to cellulases and ligninolytic enzymes to carbohydrate degrading enzymes were 2.0, 1.6 and 74.0, respectively (Fig. 3c).
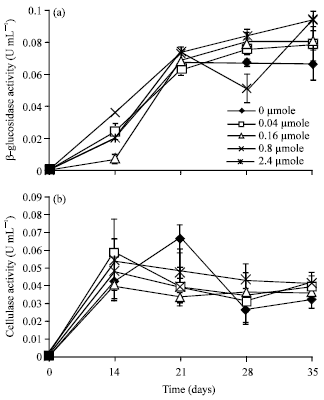 | |
| Fig. 2(a-b): | β-glucosidase (a) and Cellulase activity and (b) in the absence and presence of 0.04, 0.16, 0.8 and 2.4 μmole CuSO4 per gram solid substrate at different times of cultivation |
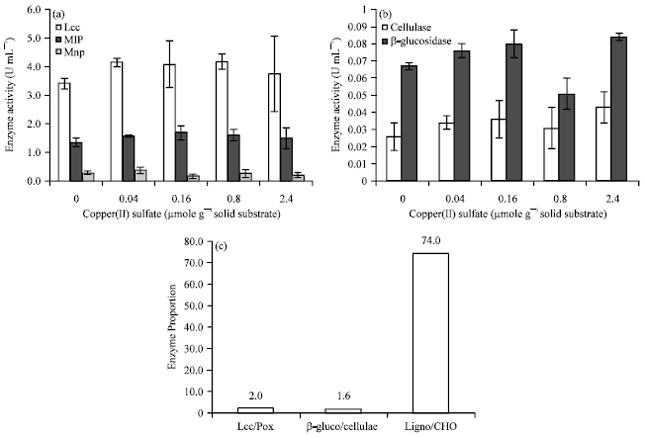 | |
| Fig. 3(a-c): | Ligninolytic enzymes (a) carbohydrate degrading enzymes, (b) production by L. polychrous grown for 28 days on solid-state fermentation with various concentrations of CuSO4 and comparison in enzyme ratios of 0.8 μmole CuSO4 per gram solid substrate and (c) Activity of Lcc, laccase; Pox, peroxidase; β-gluc, β-glucosidase; Ligno, ligninolytic enzyme; CHO, carbohydrate degrading enzyme |
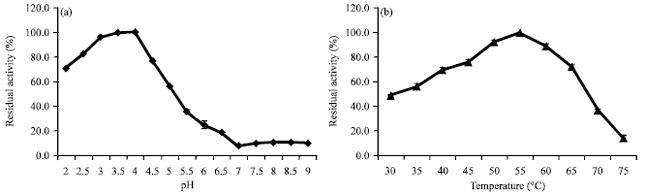 | |
| Fig. 4(a-b): | pH optimum (a) and temperature optimum (b) of crude laccase from L. polychrous grown on solid-state fermentation with 0.8 μmole CuSO4 per gram solid substrate |
Some white rot fungi secrete carbohydrate-degrading enzymes, apart from ligninolytic enzymes during lignin degradation in nature including xylanase, cellulase and glucosidase (Leonowicz et al., 1999; Quiroz-Castaneda et al., 2009).
Enzyme characteristics: The crude enzyme of L. polychrous from SSF supplemented with 0.8 μmole CuSO4 per gram solid substrate was studied it’s optimize condition for catalysis. The results showed that the optimum pH for laccase catalysis was in the range of 3-4 (Fig. 4a) and optimum temperature was at 55°C (Fig. 4b). The pH and thermal stability of crude laccase of this fungus were also experimented. The crude laccase was well stabilized in distilled water as well as in buffers pH 6-8 (approximately 90 and 80% residual activity at 24 h compared to the original (Fig. 5a).
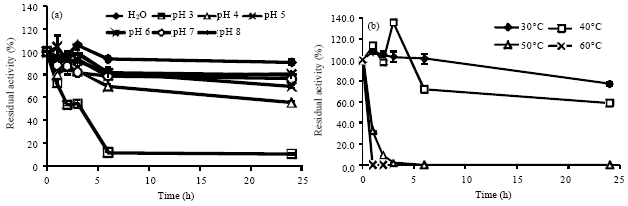 | |
| Fig. 5(a-b): | pH stability (a) and thermal stability, (b) of crude laccase from L. polychrous grown on solid-state fermentation with 0.8 μmole CuSO4 per gram solid substrate |
The thermal stability of the crude laccase, when the temperature is high, crude laccase tended to denature. The crude laccase was well stabilized at 30°C in 0.1 M sodium acetate buffer, pH 4.5 with residual activity more than 80% after 24 h incubation (Fig. 5b). Moreover, at temperature of 40°C, laccase could well stabilize for several hours, even though its activity was decreased to about 70% residual activity at 5 h incubation but at 24 h its activity was about 60% left. Even though its optimum temperature for catalysis seemed as high as 50-55°C (Fig. 4b), in this temperature the enzyme lost its activity quickly and completely inactive at 2-3 h incubation (Fig. 5b). These aspects of the enzymes from this fungus are important to consider if temperature is a key in industrial process in order to optimize the application of enzyme biocatalysts.
Electrophoresis of the crude enzyme in native condition revealed that there are at least two isoenzymes (or isoforms) of laccases as it appeared two bands of activity stains with laccase substrates ABTS and syringaldazine (Fig. 6 lane 1, 2). Interestingly, those bands with laccase activities also stained with peroxidase substrates (Fig. 6 lane 3, 4). These results were agreed with previous report of crude enzyme of L. polychrous culture in solid state of rice husk and bran without CuSO4 inducer by Sarnthima et al. (2009) in which more than one laccase isoenzymes (or isoforms) observed. However, instead of three bands of activity zymogram, only two bands observed here in this work. These results indicate that CuSO4 have certain effect on extracellular enzyme production and secretion by this fungus. The protein bands of coomassie staining showed more than two bands by two of them were at the same electrophoretic mobility corresponding to extracellular enzymes (Fig. 6 lane 5).
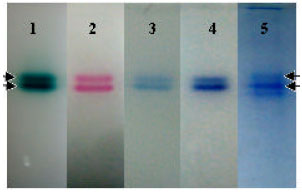 | |
| Fig. 6: | Native-PAGE of crude laccase from L. polychrous grown on solid-state fermentation with 0.8 μmole CuSO4 per gram solid substrate, stained with substrates of ligninolytic enzymes and with coomassie brilliant blue R-250. 1: with ABTS, 2: with Syringaldazine, 3: MBTH+DMAB+Mg2++ H2O2, 4: Syringaldazine followed with MBTH+ DMAB+Mg2++H2O2, 5: with coomassie brilliant blue R-250. Arrows indicate expected ligninolytic isoenzymes |
It seemed that the fungus secrete proteins mainly ligninolytic enzymes with low amount of carbohydrate degrading enzymes which might be other two bands that not corresponding with ligninolytic activities (highest and lowest electrophoretic mobility). If possible, activity stain with β-glucosidase and cellulase substrates might reveal more insight information of the crude enzyme of this fungus. Moreover, successful purification of those enzymes would be very useful in order to further study about their properties, structure and function relationship. Versatile peroxidases are hybrids of lignin peroxidase and manganese peroxidase with a bifunctional characteristic (Wong, 2009).
Black liquor decolorization: Dark colors in pulp and paper mill effluent come from lignin and its derivatives. Lignin compounds are polymer substances which complex in structure and not simple to degrade. Since the fungi, especially the white rot fungi are reported about their ability to degrade lignin in nature, because they can produce and secrete ligninolytic enzymes for lignin degradation and use as a carbon source. Lininolytic enzymes from L. polychrous mainly are laccase and MnP but no LiP activity could be detected. They have potential uses in many areas, especially dye decolorization. Therefore, the crude laccase was tested for black liquor decolourization.
To evaluate the potential application of the crude enzyme from L. polychrous for black liquor decolorization, reactions were done at various pH conditions (pH 3-9) using laccase activity 0.2 U mL-1 in the absence and presence of ABTS and HBT redox mediators (0.05 mM) at 32°C for 24 h. The results are as shown in Fig. 7a-c. In the absence of redox mediator and in the presence of ABTS, only about 10% black liquor decolorization in buffer pH 8.0 could be observed at 24 h. Black liquor decolorization have been reported by some other white rot fungi such as Cyathus bulleri and others accompanying with certain redox mediators (Da Re and Papinutti, 2011).
Black liquor decolorization at pH 3 occurred precipitation of lignin in reactions in all tested condition. At this pH, in the absence of ABTS redox mediator showed highest percentage decolorization of 52.2% at 5 h (Fig. 7a), while in the presence of ABTS gave the highest percentage decolorization of 56.8% at 5 h (Fig. 7b) and about 50% at 24 h in the presence of HBT redox mediator (Fig. 7c). At the rest pH conditions the black liquor reactions increased in its maximum absorbance (minus graphs), suggesting that polymerization might occurred instead of decolorization (Fig. 7a-c).
During black liquor decolorization reactions, the residual activity of laccase was monitoring in each reaction tube at different times. In the experiment, both absence and presence of ABTS or HBT redox mediators, the laccase activity tended to decrease when the time progresses (Fig. 8a-c). The crude laccase was well stabilized in pH 4-6 whereas in other pH conditions, laccase activity clearly decreased with reaction time.
The crude enzyme seemed to not able to decolorize black liquor even in the presence of ABTS or HBT redox mediator even though the crude laccase fairly stable throughout experimental time scales. However, at pH 3 it introduced a lignin precipitation and made paler black liquor where the laccase was least stable. Lignin could precipitate in acid condition, at pH 3 was also observed lignin precipitation in this work after stand for 24 h. However, in enzymatic reaction of the crude enzyme such precipitation could be observed within 3 h. This result suggest that reaction of lignin by the crude enzyme might accelerate precipitation of lignin compounds in the black liquor.
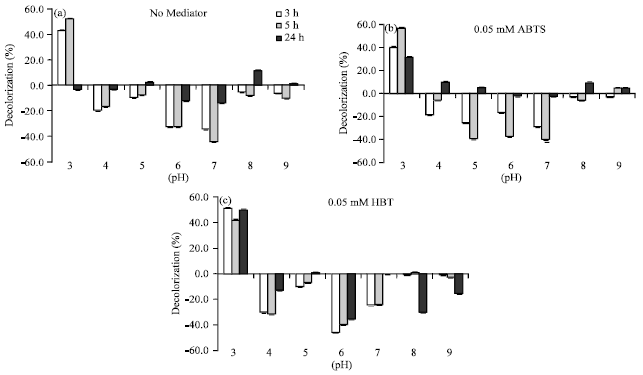 | |
| Fig. 7(a-c): | Black liquor decolorization in various pH conditions in the absence of redox mediator (a) in the presence of redox mediator, 0.05 mM ABTS (b) and 0.05 mM HBT (c) |
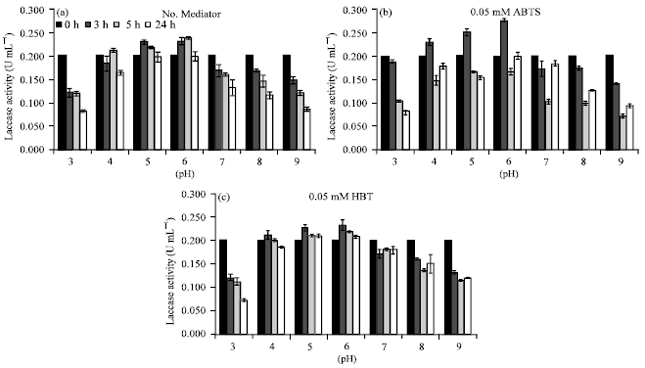 | |
| Fig. 8(a-c): | Remaining laccase activity in reactions of black liquor decolorization in various pH conditions in the absence of redox mediator (a) in the presence of redox mediator, 0.05 mM ABTS (b) and 0.05 mM HBT (c) |
This might be applied crude enzyme of the fungus for recovery of lignin for another purpose use instead of concentrate acid precipitation. Lignin precipitation by acid has been reported in many works (Koljonen et al., 2004; Fernando et al., 2010). Today, lignin extraction from black liquor is an interesting option for pulp mills due to possibility to increase the production capacity of pulp without increasing the load in the recovery boiler. Moreover, extracted lignin can be used as a solid biofuel or as a feed stock producing various chemicals. Application of crude laccase of L. Polychrous in acidic condition might shorten the precipitation process and lower use of concentrate acid.
CONCLUSION
Lentinus polychrous produced ligninolytic enzymes including laccase, MIP and MnP but no LiP activities. Copper(II)sulfate at low concentration could induce laccase production and affected isoenzyme pattern. No xylanase activity but low cellulase and β-glucosidase activities, fairly high optimum temperature and thermal stability as well as be able to work in pH 6-8 suggested the crude ligninolytic enzyme from this fungus a promising potential to apply for lignin removal in pulp biobleaching or treatment of wastewater.
ACKNOWLEDGMENTS
The authors would like to thank the Thailand Research Fund Master Research Grant No. MRG-WI525S104 as well as the Center of Excellence for Innovation in Chemistry (PERCH-CIC), Department of Chemistry, Faculty of Science, Mahasarakham University. Many thanks also go to the Phoenix Pulp and Paper Public Company Limited for a partial financial support.
REFERENCES
- Bailey, M.J., P. Biely and K. Poutanen, 1992. Interlaboratory testing of methods for assay of xylanase activity. J. Biotechnol., 23: 257-270.
CrossRefDirect Link - Baldrian, P., 2006. Fungal laccases-occurrence and properties. FEMS Microbiol. Rev., 30: 215-242.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Cordi, L., R.C. Minussi, R.S. Freire and N. Duran, 2007. Fungal laccase: Copper induction, semi-purification, immobilization, phenolic effluent treatment and electrochemical measurement. Afr. J. Biotechnol., 6: 1255-1259.
Direct Link - D'Souza, T.M., C.S. Merritt and C.A. Reddy, 1999. Lignin modifying enzymes of the white rot basidiomycete Ganoderma lucidum. Applied Environ. Microbiol., 65: 5307-5313.
PubMedDirect Link - Davis, B.J., 1964. Disc electrophoresis. II. Method and application to human serum proteins. Ann. N. Y. Acad. Sci., 121: 404-427.
CrossRefPubMedDirect Link - Eggert, C., U. Temp and K.E.L. Eriksson, 1996. The ligninolytic system of the white rot fungus Pycnoporus cinnabarinus: Purification and characterization of the laccase. Applied Environ. Microbiol., 62: 1151-1158.
Direct Link - Fernando, E.F., E.M. Vallejos and M.C. Area, 2010. Lignin recovery from spent liquors from ethanol water fractionation of sugar cane baggasse. Cellulose Chem. Technol., 44: 311-318.
Direct Link - Geng, X. and K. Li, 2002. Degradation of non-phenolic lignin by the white-rot fungus Pycnoporus cinnabarinus. Applied Microbiol. Biotechnol., 60: 342-346.
CrossRef - Ghose, T.K., 1987. Measurement of cellulase activities. Pure Applied Chem., 59: 257-268.
CrossRefDirect Link - Khammuang, S. and R. Sarnthima, 2007. Laccase from spent mushroom compost of Lentinus polychrous Lev. and its potential for remazol brilliant blue R decolourisation. Biotechnology, 6: 408-413.
CrossRefDirect Link - Koljonen, K., M. Osterberg, M. Kleen, A. Fuhrmann and P. Stenius, 2004. Precipitation of lignin and extractives on kraft pulp: Effect on surface chemistry, surface morphology and paper strength. Cellulose, 11: 209-224.
CrossRef - Lara, M.A., A.J. Rodriguez-Malaver, O.J. Rojas, O. Holmquist and A.M. Gonzalez et al., 2003. Blackliquor lignin biodegradation by Trametes elegans. Int. Biodeteriorat. Biodegrad., 52: 167-173.
CrossRef - Leonowicz, A., A. Matuszewska, J. Luterek, D. Ziegenhagen and M. Wojtas-Wasilewska et al., 1999. Biodegradation of lignin by white rot fungi. Fungal Genet. Biol., 27: 175-185.
CrossRef - Levin, L., F. Forchiassin and A.M. Ramos, 2002. Copper induction of lignin-modifying enzymes in the white-rot fungus Trametes trogii. Mycologia, 94: 377-383.
Direct Link - Liew, C.Y., A. Husaini, H. Hussain, S. Muid, K.C. Liew and H.A. Roslan, 2010. Lignin biodegradation and ligninolytic enzyme studies during biopulping of Acacia mangium wood chips by tropical white rot fungi. World J. Microbiol. Biotechnol., 27: 1457-1468.
CrossRef - Miller, G.L., 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem., 31: 426-428.
CrossRefDirect Link - Ornstein, L., 1964. Disc electrophoresis-I: Background and theory. Ann. N. Y. Acad. Sci., 121: 321-349.
CrossRefPubMedDirect Link - Palmieri, G., P. Giardina, C. Bianco, B. Fontallella and G. Sannina, 2000. Copper induction of laccase isoenzymes in the lignolytic fungus Pleurotus ostreatus. Applied Microbiol. Biotechnol., 66: 920-924.
Direct Link - Pezzella, C., F. Autore, P. Giardina, A. Piscitelli, G. Sannia and V. Faraco, 2009. The Pleurotus ostreatus laccase multi-gene family: Isolation and heterologous expression of new family members. Curr. Genet., 55: 45-57.
CrossRef - Quiroz-Castaneda, R.E., E. Balcazar-Lopez, E. Dantan-Gonzalez, A. Martinez, J. Folch-Mallol and C.M. Anaya, 2009. Characterization of cellulolytic activities of Bjerkandera adusta and Pycnoporus sanguineus on solid wheat straw medium. Electron. J. Biotechnol., Vol. 12, No. 4.
CrossRefDirect Link - Da Re, V. and L. Papinutti, 2011. Black liquor decolorization by selected white-rot fungi. Applied Biochem. Biotechnol., 165: 406-415.
CrossRefDirect Link - Sarnthima, R., S. Khammuang and J. Svasti, 2009. Extracellular ligninolytic enzymes by Lentinus polychrous Lev. under solid-state fermentation of potential agro-industrial wastes and their effectiveness in decolorization of synthetic dyes. Biotechnol. Bioprocess Eng., 14: 513-522.
CrossRefDirect Link - Arora, D.S. and R.K. Sharma, 2010. Ligninolytic fungal laccases and their biotechnological applications. Applied Biochem. Biotechnol., 160: 1760-1788.
CrossRefDirect Link - Srisomsap, C., J. Svasti, R. Surarit, V. Champattanachai and P. Sawangareetrakul et al., 1996. Isolation and characterization of an enzyme with β-glucosidase and β-fucosidase activities from Dalbergia cochinchinensis Pierre. J. Biochem., 119: 585-590.
PubMedDirect Link - Wong, D.W.S., 2009. Structure and action mechanism of ligninolytic enzymes. Applied Biochem. Biotechnol., 157: 174-209.
CrossRefDirect Link
Precious Reply
Whats the original source of Lentinus polychrous Lèv, thanks.
rakrudee sarnthima
The fungal strain isolated naturally from the forest in Thailand