
P. Songsri
Department of Plant Science and Agriculture Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
B. Suriharn
Department of Plant Science and Agriculture Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
J. Sanitchon
Department of Plant Science and Agriculture Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
S. Srisawangwong
Khon Kaen Agricultural Research and Development Center, Thapra, Khon Kaen 40000, Thailand
T. Kesmala
Department of Plant Science and Agriculture Resources, Faculty of Agriculture, Khon Kaen University, Khon Kaen 40002, Thailand
Journal of Biological Sciences
Year: 2011 | Volume: 11 | Issue: 3 | Page No.: 268-274







ABSTRACT
Narrow genetic base is a main problem hindering the progress of physic nut breeding. The objective of this study was to compare the effects of different levels of gamma radiation on germination percentage, survival percentage and growth of five physic nut genotypes. The seeds were treated with six levels of gamma radiation (0, 200, 400, 600, 800 and 1000 Gy) and the 5x6 treatment combinations were arranged in a randomized complete block design with three replications. Data were recorded for germination percentage, survival percentage at 15 Days After Germination (DAG) (GR50(15)) and survival percentage at 30 DAG (LD50(30)). As the seedlings did not survive at 600, 800 and 1000 Gy, these treatments were deleted from the successive evaluations of plant height, stem diameter and leaf number. Higher doses of gamma radiation resulted in the significant reductions in germination percentage, survival percentage and plant height. The germination percentages were from 4.0% for 1000 Gy to 82.5% for 0 Gy. The seeds treated with gamma radiation higher than 600 Gy resulted in complete mortality. Difference among genotypes was not significant for these characters. The dosages of gamma radiation to obtain LD50 based on survival percentage 30 DAG was 425 Gy and was not dependent on genotypes. The information is useful for gamma radiation induction of mutation breeding for physic nut.
PDF Abstract XML References Citation
Received: May 09, 2011;
Accepted: July 12, 2011;
Published: August 15, 2011
How to cite this article
P. Songsri, B. Suriharn, J. Sanitchon, S. Srisawangwong and T. Kesmala, 2011. Effects of Gamma Radiation on Germination and Growth Characteristics of Physic Nut (Jatropha curcas L.). Journal of Biological Sciences, 11: 268-274.
DOI: 10.3923/jbs.2011.268.274
URL: https://scialert.net/abstract/?doi=jbs.2011.268.274
DOI: 10.3923/jbs.2011.268.274
URL: https://scialert.net/abstract/?doi=jbs.2011.268.274
INTRODUCTION
Physic nut (Jatropha curcas L.) is originated in Central America and the commercial plantations with economic importance were first established in the Cape Verde Islands for lighting oil and other products. Physic nut was introduced into Africa and Asia by Portugal traders and now it is important as the oil crop for biodiesel production in the tropics (Heller, 1996).
Ambrosi et al. (2010) found that seeds of physic nut commercialized worldwide include a few closely related genotypes which are not representative of the original Mexican gene pool, revealing high degrees of homozygosity for single varieties and very low genetic diversity between varieties. Narrow genetic base would be expected in the physic nut varieties used for cultivation in Asia and Africa. The lack of wide genetic base hinders the progress in physic nut breeding and the use of un-adapted varieties leads to low and unpredictable yield.
Tanya et al. (2011) found low ISSR variability within groups (37%) of physic nut germplasm collected in Thailand, Vietnam and China but the variability of related species was rather high (63%). Among 225 accessions of physic nut from Africa, Asia and Latin America, Montes et al. (2009) found low genetic variability in accessions from Africa and Asia but high in accessions from Guatemala and other Latin American countries. In Kenya, moderate to high genetic variation within populations and among populations of physic nut was reported and the high genetic variation in this case is possibly due to several introductions from many countries (Machua et al., 2011).
Although genetic variability of physic nut is high in the center of origin, most physic nut breeding programs have been carried out outside the center of origin. To the best of our knowledge, the international organization for conservation and free transfer of germplasm has not been established for physic nut. Therefore, physic nut breeders outside the center of origin face difficulty in obtaining good germplasm sources for their breeding programs (Divakara et al., 2010; Datta, 2009).
By using available germplasm, mutation breeding is an alternative way to increase genetic variations of economically important characters. This could be achieved through chemical and physical mutations. Gamma radiation is another way to induce mutation and create genetic variations in narrow-based germplasm (Datta, 2009).
In physic nut, attempts have been made to induce mutation through radiation treatment of the seeds. Treated seeds with gamma radiation at 50 to 250 grays showed no significant difference between treated seeds and un-treated seeds for germination percentage and survival percentage. Therefore, the medial lethal dose that causes 50% mortality of the seeds (LD50) is to be determined and seeds of different genotypes that may respond differently is also needed to be addressed. The objectives of this study were to find out the optimum doses for radiation treatment of physic nut seeds and to study the sensitivity of different physic nut genotypes to gamma radiation treatments. The information obtained will be useful as a guideline for mutation breeding using gamma ray.
MATERIALS AND METHODS
Five physic nut genotypes collected from different parts of Thailand were used in this study. These genotypes including 18/36, 19/18, 20/99, 18/42 and Check-I were kindly provided by the Khon Kaen Agricultural Research and Development Center at Thapra, Khon Kaen province. The Check-I is a promising variety selected from multiple yield trials. The seeds of these genotypes were treated with acute gamma radiation at the doses of 0, 200, 400, 600, 800 and 1000 grays at the Department of Applied Radiation and Isotope, Kasetsart University in Bangkok.
The M1 seeds were used in this study. Germination test was carried out in a greenhouse at the Horticulture Department, Khon Kaen University in 2010. The seeds were germinated in trays with holes for drainage containing sand. A 5x6 factorial experiment in a randomized complete block design with three replications was used. Five physic nut genotypes were assigned as factor A and six gamma radiation doses were assigned as factor B. There were 25 seeds for each sampling unit. The data were recorded for germination percentage at 5-10 days after sowing. The germinated seeds were later transferred to plastic bags at 15 days after germination when survival percentage at 15 days after germination was recorded. The plants were allowed to grow in plastic bags for 15 days and survival percentage at 30 Days After Germination (DAG) was recorded.
The survival percentage at 15 days after germination was then calculated as follows:
The survival percentage at 30 DAG was then calculated as follows:
The survival as percent of control at 15 DAG (GR50(15)) was then calculated as follows:
The survival as percent of control at 30 DAG (LD50(30)) was then calculated as follows:
The healthy seedlings with one-month old were later transplanted into pots containing rice husk, compost, chopped coconut husk and soil at the ratio of 2:2:2:1 by volume. As there was no survival seedling for gamma radiation treatments at 600, 800 and 1000 Gy, these treatments were deleted from the experiment. A 5x3 factorial experiment in a randomized complete block design with six replications was further carried out. Five physic nut genotypes were assigned as factor A and three gamma radiation doses were assigned as factor B. There were four pots for each sampling unit each of which contained one plant. The plants were allowed to grow until 72 days after transplanting and the data were evaluated for stem diameter, plant height and leaf number per plant at seven-day intervals from 30 days after transplanting until 72 days after transplanting. Other morphological characters such as leaf color and leaf shape were also observed.
Statistical analysis: The data were subjected to analysis of variance according to a factorial experiment in a randomized complete block design (Gomez and Gomez, 1984). Where the main effects were significant, mean separation was carried out using Duncan’s Multiple Range Test (DMRT). All calculations were done in MSTAT-C and graphical presentation was done in Microsoft Excel.
RESULTS
Analysis of variance showed significant differences (p≥0.01) among gamma ray treatments for germination percentage, survival percentage at 15 Days After Germination (DAG), survival percentage at 30 DAG (Table 1), stem diameter, plant height and number of leaf per plant (Table 2). The genotypes were not statistically different for these characters and the interactions between gamma ray treatment and genotype were also not significant (p≥0.05) (Table 1, 2).
 | |
| Fig. 1: | Seedlings at germination of physic nut subjected to different levels of gamma radiation |
| Table 1: | Mean squares for germination, survival percentage at 15 days after germination (DAG) and survival percentage at 30 DAG of five physic nut genotypes subjected to six levels of gamma radiation |
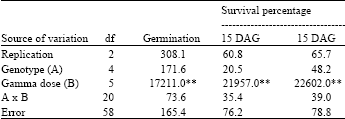 | |
| ** Significant at 0.01 probability level, df: Degree of freedom | |
| Table 2: | Mean squares for stem diameter, plant height and number of leaves per plant of five physic nut genotypes subjected to six levels of gamma radiation evaluated from germination to 72 Days After Transplanting (DAT) |
 | |
| ** Significant at 0.01 probability level df: Degree of freedom | |
| Table 3: | Germination percentage, survival percentage at 15 days after germination (DAG), survival percentage at 30 DAG, stem diameter (mm), plant height (cm) and number of leaves per plant of five physic nut genotypes subjected to six levels of gamma radiation evaluated from germination to 72 Days After Transplanting (DAT) |
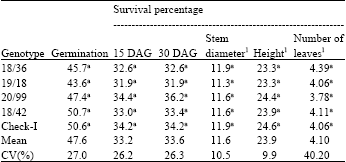 | |
| 1Stem diameter, height and leaf number were calculated from plants receiving 0, 200 and 400 grays and evaluated at 72 DAT. Means in the same column with the same letter(s) are not statistically different by Duncan’s multiple range test (DMRT) at p≤0.05 | |
| Table 4: | Effects of gamma radiation on germination percentage, survival percentage at 15 days after germination (DAG), survival percentage at 30 DAG, stem diameter (mm), plant height (cm) and number of leaves per plant of five physic nut genotypes subjected to six levels of gamma radiation evaluated from germination to 72 days after transplanting |
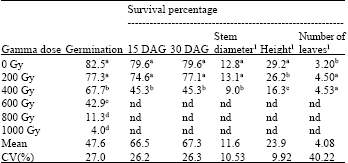 | |
| nd = not determined (High germination was still observed at 600 Gy but the germinated seedlings did not survive.). 1Stem diameter, height and leaf number were calculated from plants receiving 0, 200 and 400 grays and evaluated at 72 DAT. Means in the same column with the same letter(s) are not statistically different by Duncan’s multiple range test (DMRT) at p≥0.05 | |
The overall means of the experiment across genotypes and radiation treatments were 47.6% for germination, 33.2% for survival at 15 DAG, 33.6% for survival at 30 DAG, 11.6 mm for stem diameter, 23.9 cm for plant height and 4.08 leaves for number of leaves per plant (Table 3).
The differences among gamma ray treatments were observed for germination percentage, survival percentage at 15 DAG and survival percentage at 30 DAG, stem diameter, plant height and number of leaves per plant (Table 4). On average, germination percentages were from 4.0% for 1000 Gy to 82.5% for 0 Gy. The seeds treated with 600-1000 Gy did not survive and hence the calculation of survival percentage was done for 0, 200 and 400 Gy (Fig. 1). Survival percentages at 15 DAG were in a range of 45.3% for 400 Gy and 79.6% for 0 Gy and germination percentages at 30 DAG were in a range of 45.3% for 400 Gy and 79.6% for 0 Gy.
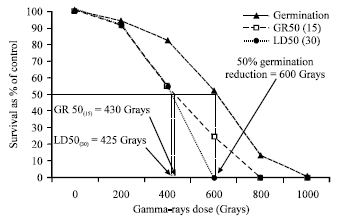 | |
| Fig. 2: | Germination percentage, survival as percent of control at 15 days after germination and survival as percent of control at 30 days after germination of five physic nut genotypes subjected to six levels of gamma radiation |
Stem diameters were lowest (9.0 mm) for the seeds receiving 400 Gy of gamma radiation and highest (13.1 mm) for the seeds receiving 200 Gy (Table 4). Plant heights were lowest (16.3 cm) for the seeds receiving 400 grays and highest (29.2 cm) for the seeds receiving 0 Gy. Leaf numbers, in contrast, were highest (4.53 leaves) for the seeds receiving 400 Gy and lowest (3.20 leaves) for the seeds receiving 0 Gy.
The seeds receiving 0 Gy and 200 Gy of gamma radiation were not statistically different for germination percentage, survival percentage at 15 and 30 DAG and stem diameter but they were statistically different for plant height and number of leaves per plant (Table 4). The reductions in germination, survival percentage at 15 DAG, survival percentage at 30 DAG and stem diameter were observed at 400 Gy but not at 200 Gy. Gamma radiation significantly reduced plant height observed at 200 Gy and 400 Gy. Numbers of leaves per plant were higher in the seeds receiving 200 and 400 Gy (4.53 and 4.50 leaves, respectively) than those receiving 0 Gy (3.20 leaves).
The germination of the control seeds was lower than 100% and the data were then converted to 100% germination and the germination of other gamma radiation treatments were calculated accordingly (Fig. 2). The data for germination percentage, survival as percent of control at 15 DAG (GR50(15)) and 30 DAG (LD50(30)) were plotted against gamma radiation dosages to determine Lethal Dose (LD50) of the radiation. LD50 for germination percentage could be as high as 600 Gy. However, at 600 Gy, the germinated seeds did not survive and germination percentage alone was not a good prediction for LD50. The GR50(15) and LD50(30) are good predictions for LD50 and the LD50 for these criteria was at 430 and 425 Gy, respectively (Fig. 2).
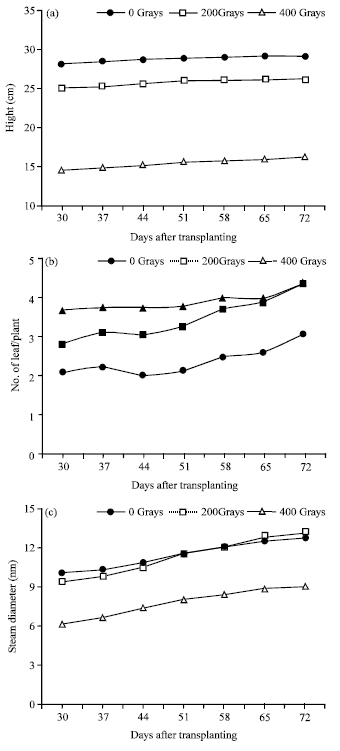 | |
| Fig. 3(a-c): | Means for plant height (a), number of leaves per plant (b) and stem diameter (c) of five physic nut genotypes seedlings subjected to different levels of gamma radiation |
Figure 4 showed growth patterns of the seeds treated with 0, 200 and 400 Gy for plant height, stem diameter and number of leaves per plant. Gamma radiation greatly reduced plant height and the effect was highest at 400 Gy (Fig. 3).
 | |
| Fig. 4: | Seedlings at 30 days after germination of physic nut subjected to different levels of gamma radiation (0, 200 and 400 grays, respectively) |
Gamma radiation also reduced stem diameter but the effect was significant at 400 Gy. Gamma radiation significantly increased leaf number for both at 200 and 400 Gy.
DISCUSSION
Exposure to gamma radiation is known to produce morphological mutants, physiological mutants and biochemical mutants. As physic nut is a long-lived perennial shrub crops, these types of mutants have been sought to increase genetic variation (Nambisan, 2007). Gamma radiation has been reported to have beneficial effects on many crops. In African violets (Saintpaulia ionantha H. Wendl.), gamma radiation at 15 Gy improved plant architecture of the mutants (Seneviratne and Wijesundara, 2007). In wheat, gamma radiation improved germination, plant height, grain per plant and grain yield at 200 Gy (Jamil and Khan, 2002) and 300 to 400 Gy (Irfaq and Nawab, 2001). In this study, the estimation of medial lethal dose and the sensitivity of different physic nut genotypes to gamma radiation were reported.
In earlier studies, Wang et al. (2009) found that LD50 doses of gamma radiation were 178 to 198 Gy for seeds of physic nut from different sources in China and India. Dhakshanamoorthy et al. (2010) exposed physic nut seeds to gamma radiation at 50 to 250 Gy and found that these doses were still lower than LD50 dose. In this study, LD50 dose was estimated roughly as 425 Gy and there was no difference among seed sources. LD50 doses found in this study were rather higher than those reported previously.
In snap bean (Phaseolus vulgaris), however, the LD50 for germination could not be determined from the doses 300 Gy to 800 Gy because the radiation had small effect on germination and some treatments had higher germination than control (Ellafa et al., 2007). Kon et al. (2007) reported in long bean (Vignas esquipedalis) that LD50 for germination ranged between 600-800 Gy and also in tomato and okra (Norfadzrin et al., 2007). In contrast, gamma radiation doses of 200 to 800 Gy did not have significant effect on germination in pepper grass (Lepidum sativum L.) (Majeed et al., 2009) and castor bean (Ricinus communis) was sensitive to gamma radiation higher than 2000 Gy for germination (Kobori et al., 2010). LD50 dose of 425 Gy in this study was much lower than those in previous studies.
The differences in LD50 doses of different studies might be possibly due to seed moisture rather than seed source and LD50 doses were also different among species. Seeds with low moisture seem to be more tolerant to gamma radiation than the seeds with high moisture. Although seed moisture was not determined in this study, it could be assumed that the seeds had low moisture (8-9%) because the seeds were sun-dried and stored for a period in cool room.
In comparison to physic nut cuttings that had much higher moisture than seeds, Dwimahyani and Ishak (2004) found that exposure of physic nut cuttings to 10-25 Gy doses of gamma radiation was sufficient to induce mutation and Li-Rong et al. (2008) found that LD50 doses for leaf segments, nodular stem segments and stem segment of Pogostemoncablin in vitro were 72, 64 and 66 Gy, respectively.
Gamma radiation at higher doses resulted in the reduction in germination (200-400 Gy) to complete fatality (600-1000 Gy) of physic nut seeds. Dhakshanamoorthy et al. (2010) found that seeds treated with 50 Gy of gamma rays revealed stimulatory effect except for germination, whereas 250 Gy dose showed inhibitory effects on traits compared to other treatments. This could be due to the damage in seed tissues and the severity of the damage depending on the dosage used (Datta, 2009). Gamma radiation had inhibitory effects on physiological and physical traits (Khan and Goyal, 2009) and also caused mutation (Dhakshanamoorthy et al., 2011). The inhibitory effects of gamma radiation were also reported in chickpea (Khan et al., 2005) mungbean (Khan and Goyal, 2009) and snap bean (Ellafa et al., 2007).
Dhakshanamoorthy et al. (2010) and Dhakshanamoorthy et al. (2011) used gamma radiation of 50-250 Gy and Datta (2009) used 60-240 Gy to induce mutation in physic nut. The previous studies used the rates that may be sufficient for induction of mutation inn physic nut seeds. In this study, we reported lethal dose of 600 Gy that induced complete fatality in physic nut seeds. The gamma radiation doses at the range of 200-400 Gy are suitable for the mutation induction in physic nut seeds. At these intensities, the M1 plants can survive and change in characters such as shorter plant, higher leaf number, shorter internodes and higher branch number that may contribute to higher yield.
In this study, the rates of gamma ray for mutation in physic nut were in the range of 425 to 430 Gy according to GR50 dose at 15 DAG and LD50 dose at 30 DAG, respectively. However, the observations were carried out in the seedlings of the M1 generation and further investigation in the more advanced generations are still required to ensure the stability of the traits such as shorter plant, early flowering, male and female flower ratio, fruiting and yield under field conditions. Molecular study for mutants and original plants is also necessary to identified genetic changes in the mutants.
CONCLUSION
Higher dosages of gamma radiation reduced germination of physic nut seeds, number of survival plants and plant height. The differences between physic nut varieties for these traits were not statistically significant. The dosages that are suitable for mutation induction in physic nut seeds are in the range of 425-430 Gy according to medial lethal dose at 15 and 30 days after germination. The information provides basic requirements for the use of gamma radiation for mutation induction in physic nut.
ACKNOWLEDGMENTS
We are grateful for the financial support provided by the Khon Kaen University and the Plant Breeding Research Center for Sustainable Agriculture, Faculty of Agriculture, Khon Kaen University, Khon Kaen, Thailand.
REFERENCES
- Ambrosi, D.G., G. Galla, M. Purelli, T. Barbi and A. Fabbri et al., 2010. DNA Markers and FCSS analyses shed light on the genetic diversity and reproductive strategy of Jatropha curcas L. Diversity, 2: 810-836.
CrossRef - Divakara, B.N., H.D. Upadhyaya, S.P. Wani and C.L. Laxmipathi Gowda, 2010. Biology and genetic improvement of Jatropha curcas L.: A review. Applied Energy, 87: 732-742.
CrossRefDirect Link - Dhakshanamoorthy, D., R. Selvaraj and A. Chidambaram, 2010. Physical and chemical mutagenesis in Jatropha curcas L. to induce variability in seed germination, growth and yield traits. Rom. J. Biol. Plant Biology, 55: 113-125.
Direct Link - Dhakshanamoorthy, D., R. Selvaraj and A.L.A. Chidambaram, 2011. Induced mutagenesis in Jatrophacurcas L. using γ rays and detection of DNA polymorphism through RAPD marker. Comptes Rendus. Biol., 334: 24-30.
CrossRefDirect Link - Dwimahyani, I. and Ishak, 2004. Induced mutation on Jatropha (Jatropha curcas L.) for improvement of agronomic characters variability. Atom. Indonesia J., 30: 53-60.
Direct Link - Ellafa, K., O.H. Ahmed, S. Shaharudin and D.A. Rahman, 2007. Gamma radio sensitivity study on snap bean (Phaseolus vulgaris). Int. J. Agric. Res., 2: 844-848.
Direct Link - Gomez, K.A. and A.A. Gomez, 1984. Statistical Procedure for Agricultural Research. 2nd Edn., John Wiley and Sons, Hoboken, New Jersey, ISBN: 9780471870920, Pages: 704.
Direct Link - Irfaq, M. and K. Nawab, 2001. Effect of γ irradiation on some morphological characteristics of three wheat (Triticum aestivum L.) cultivars. J. Biol. Sci., 1: 935-937.
CrossRefDirect Link - Jamil, M. and U.Q. Khan, 2002. Study of genetic variation in yield components of wheat cultivar bukhtwar-92 as induced by γ radiation. Asian J. Plant Sci., 1: 579-580.
CrossRefDirect Link - Khan, M.R., A.S. Qureshi, S.A. Hussauin and M. Ibrahim, 2005. Genetic variability induced by γ irradiation and its modulation with gibberellic acid in M2 generation of Chickpea (Cicer arietinum L.). Pak. J. Bot., 37: 285-292.
Direct Link - Khan, S. and S. Goyal, 2009. Improvement of mungbean varieties through induced mutations. Afr. J. Plant Sci., 3: 174-180.
CrossRefDirect Link - Kobori, N.N., T. Mastrangelo, S.M. Cicero, P. Cassieri, M.H.D. Moraes and J.M.M. Walder, 2010. Effects of γ irradiation on physiological and phytosanitary qualities of brazilian castor bean seeds, Ricinus communis (cv. IAC Guarani). Res. J. Seed Sci., 3: 70-81.
CrossRefDirect Link - Kon, E., O.H. Ahmed, S. Saamin and N.M.A. Majid, 2007. Gamma radiosensitivity study on long bean (Vigna sesquipedalis). Am. J. Applied Sci., 4: 1090-1093.
Direct Link - Li-Rong, W.U., H. Hong, Z. Yan-Ling, L. Xing and L. Xiao-Hua, 2008. The effect of ~(60)COγ irradiation on culture in vitro of Pogostemoncablin cablin (BLANCO) Benth. J. Nucl. Agric. Sci., 22: 14-17.
Direct Link - Majeed, A., Z. Muhammad, H. Ahmad and A.U.R. Khan, 2009. Gamma irradiation effects on some growth parameters of Lepidium sativum. Am.-Eurasian J. Sustain. Agric., 3: 424-427.
Direct Link - Machua, J., G. Muturi, S.F. Omondi and J. Gicheru, 2011. Genetic diversity of Jatropha curcas L. populations in Kenya using RAPD molecular markers: Implication to plantation establishment. Afr. J. Biotech., 10: 3062-3069.
Direct Link - Nambisan, P., 2007. Biotechnological intervention in Jatropha for biodiesel production. Curr. Sci., 93: 1347-1348.
Direct Link - Norfadzrin, F., O.H. Ahmed, S. Shaharudin and D.A. Rahman, 2007. A preliminary study on γ radiosensitivity of tomato (Lycopersicon esculentum) and Okra (Abelmoschus esculentus). Int. J. Agric. Res., 2: 620-625.
CrossRefDirect Link - Seneviratne, K.A.C.N. and D.S.A. Wijesundara, 2007. First African violets (Saintpaulia ionantha, H. Wendl.) with a changing colour pattern induced by mutation. Am. J. Plant Physiol., 2: 233-236.
CrossRefDirect Link - Tanya, P., P. Taeprayoon, Y. Hadkam and P. Srinives, 2011. Genetic diversity among Jatropha and Jatropha-related species based on ISSR markers. Plant Mol. Biol. Repor., 29: 252-264.
CrossRefDirect Link - Wang, Z.Y., J.M. Lin, L. Luo and Z.F. Xu, 2009. Sensitivity of Jatropha curcas seeds to (60)COγ radiation and their medial lethal doses in radiation breeding. Nan Fang Yi Ke Da Xue Xue Bao, 29: 506-508.
PubMedDirect Link