
Khosrow Chehri
Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Sanandaj Branch, Iran
D. Zafari
Faculity of Agriculture, Bu-Ali Sina University, Hamedan, Iran
M. Nurhazrati
School of Biological Sciences, Universiti Sains Malaysia, 11800 USM, Penang, Malaysia
B. Salleh
School of Biological Sciences, Universiti Sains Malaysia, 11800 USM, Penang, Malaysia
K.R.N. Reddy
School of Biological Sciences, Universiti Sains Malaysia, 11800 USM, Penang, Malaysia
E. Karami
Department of Agronomy, Faculty of Agriculture, Islamic Azad University, Sanandaj Branch, Iran
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 8 | Page No.: 795-799







ABSTRACT
The objective of this study was to identify Fusarium species associated with root and stalk rot of maize and their pathogenicity on root and stalk of maize in Kermanshah Province, Iran. In this survey 480 Fusarium strains were isolated and identified from maize ears collected from different geographic regions in Kermanshah province, Iran during 2006-2009. All these 480 strains belonged to 22 Fusarium species. This Fusarium strains were evaluated for their pathogenicity on maize and observed that F. verticillioides, F. subglutinans and F. pseudoanthophilum are the most pathogenic on stalk. Foot rot assessment revealed that F. verticillioides as the most damaging species. This is the first comprehensive report on identity and distribution of Fusarium spp. from maize in west of Iran while F. pseudonygamai was reported for the first time from west of Iran.
PDF Abstract XML References Citation
Received: November 21, 2010;
Accepted: January 13, 2011;
Published: February 26, 2011
How to cite this article
Khosrow Chehri, D. Zafari, M. Nurhazrati, B. Salleh, K.R.N. Reddy and E. Karami, 2010. Natural Occurrence of Fusarium Species Associated with Root and Stalk Rot of Maize in Kermanshah Province, Iran. Journal of Biological Sciences, 10: 795-799.
DOI: 10.3923/jbs.2010.795.799
URL: https://scialert.net/abstract/?doi=jbs.2010.795.799
DOI: 10.3923/jbs.2010.795.799
URL: https://scialert.net/abstract/?doi=jbs.2010.795.799
INTRODUCTION
Maize (Zea mays L.) is one of the most important cereal crops being cultivated in Iran. Worldwide reports have been shown that maize is one of the most prevalent crops contaminated by Fusarium (Fandohan et al., 2003). Fusarium spp. can cause a variety of diseases in many agricultural, horticultural and forestry crops. Due to their significant annual losses, studies on different aspects of its life cycle, biology, ecology, taxonomy and its pathogenicity has been regarded by plant pathologists. Fusarium species are a serious threat to susceptible crops worldwide, particularly maize (Nicolaisen et al., 2009). They cause root, stalk and ear rot, with severe reductions in crop yield, often estimated between 10 and 30% annually (Logrieco et al., 2002). There are two types of maize ear rot, called red ear rot and pink ear rot. Red ear rot universally caused by Discolor section and pink ear rot caused by Liseola section (Aliakbari et al., 2007). Fusarium spp. can also produce mycotoxins which relevant to animal and human health (Woloshuk and Shim, 2000; Velluti et al., 2001). F. graminearum, F. culmorum, F. cerealis, F. poae are the major producers of trichothecenes (Logrieco et al., 2002). Maize grains may be contaminated with different Fusarium toxins such as trichothecenes, zearalenone, fumonisins and moniliformin in grains that are harmful to both human and animals (IARC, 2002). Some of the strains of F. verticillioides do not produce disease in maize and are an endophyte, because it does not cause visible damage to the plant (Stone et al., 2000). The aim of this study is to identification of Fusarium species associated with maize and their pathogenicity assay on root and stalk of maize in Kermanshah province of Iran.
MATERIALS AND METHODS
Isolation and Identification of Fusarium spp.: During 2006-2009, diseased maize tissue sample were collected from 120 maize fields from different regions of Kermanshah province from West of Iran. Roots and stalks of diseased samples were washed under tap water. Then the samples were surface sterilized with 70% ethanol and cut into small blocks. The blocks were soaked in 1% sodium hypochlorite solution for 3 min and rinsed in several changes of sterile distilled water. All the sterilized samples were placed on Peptone-Pentachloronitrobenzene Agar (PPA) plates, a selective medium for Fusarium (Nash and Snyder, 1962). The plates were incubated under standard incubation conditions (Salleh and Sulaiman, 1984) for 24 h. The resulting single-spore Fusarium colonies were transferred to fresh Potato Dextrose Agar (PDA) plates and maintained at 4°C for further studies. To study the growth rates and pigment production of Fusarium spp. all the strains were transferred onto PDA plates and incubated at ambient temperature. Ten replications were maintained for each Fusarium strain. For microscopic observations, all the strains of Fusarium were transferred to Carnation Leaf Agar (CLA) (Fisher et al., 1982), Spezieller Nahrstoffarmer Agar (SNA) (Nirenberg, 1976) and Potassium Chloride Agar (KClA) (Fisher et al., 1983) medium. The species were identified on the basis of macroscopic characteristics such as pigment production and growth rates on PDA plates, as well as their microscopic features including size of macroconidia, presence of microconidia and its production in chains or false heads, type of conidiogenous cells (monophialidies and polyplialidies conidiophores) and also absence or presence of chlamydospores (Gerlach and Nirenberg, 1982). Identification of species was based on species description of Gerlach and Nirenberg (1982), Nelson et al. (1983) and Leslie and Summerell (2006).
Pathogenicity assay: All of the identified Fusarium species were tested for their pathogenicity on apparently healthy maize and individual plants were used for inoculation tests on root and stalk. The root and stalk of the maize were washed and surface sterilized before inoculation. For inoculation, each strain of Fusarium species were grown on PDA plates as described by Salleh and Sulaiman (1984). Conidial suspension of each strain was prepared by scrapping the mycelium with sterile distilled water onto 7 day-old cultures, shaken thoroughly and the concentration was adjusted to 2x106 conidia mL-1 using haemocytometer. Twenty milliliter of the spore suspension of each Fusarium species was sprayed on the root and stalk of the plants. Control plants were sprayed with 20 mL of sterile distilled water. Three replications were maintained for each strain and the experiment was repeated twice.
RESULTS AND DISCUSSION
In this study, 480 Fusarium isolates were isolated from diseased plants and based on morphological characteristics, these isolates were identified as 22 Fusarium species including F. avenaceum, F. acuminatum, F. anthophilum, F. culmorum, F. chlamydosporum, F. equiseti, F. globosum, F. graminearum, F. lateritium, F. nygamai, F. oxysporum, F. poae, F. proliferatum, F. pseudoanthophilum, F. subglutinans, F. pseudonygamai, F. solani, F. semitectum, F. sambucinum, F. sporotrichioides, F. tricinctum and F. verticillioides (Table 1, 2). Of the Fusarium isolates collected in Kermanshah province, Iran, F. verticillioides was the most prevalent with a frequency of 51.04 % (245 of 480), followed by F. proliferatum with a frequency of 30.42% (146 of 480). Among isolates, 25% from root and 75% from stalk tissue were recovered. All of the species except F. oxysporum and F. lateritium were recovered from stalk rot of maize. The species identified from root rot of maize were F. verticillioides, F. oxysporum and F. lateritium. In stalk tissues F. verticillioides, F. proliferatum, F. subglutinans, F. chlamydosporum, F. pseudoanthophilum, F. poae and in root tissue F. verticillioides were predominant species. In this survey F. pseudonygamai is reported for the first time from Iran.
Eight days after inoculation, disease symptoms were observed on the roots. Initially, the roots illustrated brownish discoloration and eventually turned to dark color, which indicates rotting of the root tissues. Twelve days after inoculation, the first symptoms on maize stalks on yellowish and then degrees of rotting were observed on the stalk base. The results in pathogenicity tests indicated that F. verticillioides, F. subglutinans and F. pseudoanthophilum are the most pathogenic on stalk and that only F. verticillioides is pathogenic on the root.
In this study, the association was observed between maize plant and the species of Gibberella fujikuroi species complex (Nirenberg and O’Donnell, 1998). In the pathogenicity test, F. verticillioides was recovered from the inoculated root and stalk which indicated that this species was able to cause root and stalk rot of maize. F. verticillioides, F. proliferatum and F. subglutinans are the common pathogens of maize in different climates (Lew et al., 1991; Miller, 2001; Logrieco et al., 2002; Munkvold, 2003). The results in this study are in accordance with other findings in Iran (Ghiasian et al., 2004), Dawodee (2002), Mohammadi et al. (2002) and Naderpour (2004) had reported that F. verticillioides and F. proliferatum as the most comon species isolated from maize in Iran. This fungus can be seed borne internally in symptomless, apparently healthy maize kernels (Thomas and Buddenhagen, 1980). Also some strains of this fungus can produce potent mycotoxins associated with serious animal and human diseases (Wilson et al., 1985). Mycotoxin-producing Fusarium species such as F. verticillioides, F. graminearum and F. poae were isolated at high levels from disease samples in this study. This indicates that the presence of high infections of stalk rot on the ears thus supporting the farmers information on the presence of the disease in their fields. Thus, identification of the different species of Fusarium, including saprobe, pathogenic and toxin producing species, is of vital importance (Nelson et al., 1994).
| Table 1: | Morphological characters of Fusarium spp. associated with maize |
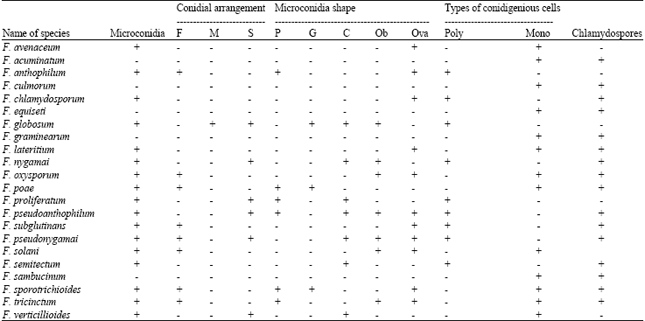 | |
| +: Presence, -: Absence, F: False heads (chains absent), M: Medium to long chains (>15), S: Short chains (<15), P: Pyriform, G: Globose, C: Clavate, Ob: Obovoid, Ova: Oval to allantoids/fusiod, Poly: Polyphialidic, Mono: Monophialidic | |
| Table 2: | Macroconidia characters of Fusarium spp. associated with maize |
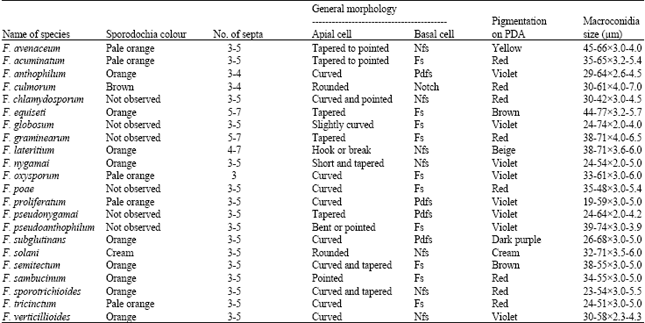 | |
| Pdfs: Poorly developed foot shape, Nfs: Notch or foot shape, Fs: Foot shape | |
Our results indicated that a section Liseola or complex of Gibberella fujikuroi species (Nirenberg and O’Donnell, 1998) could be pathogenic to the maize and suggested that F. verticillioides could be the main causal agent of stalk and root rots. However, strains of F. subglutinans and F. pseudoanthophilum were the main causal agents of stalk rot as well. In this survey, the widespread of species especially in Gibberella fujikuroi species complex offers the opportunity for relatively easy collection of large numbers of species by phylogenetically-based classification (O’Donnell, 1996). The identification of the Fusarium species contaminating maize in different areas is evidence for a need, not only for studying the levels of the interaction between Fusarium pathogens and maize, but also to obtain a precise picture of the toxicological risks related to the maize consumption by humans and animals (Logrieco et al., 2002). The present study demonstrates the importance of Fusarium stalk and root rot of maize in Iran. We believe that this study will serve as a foundation for the further identification of Fusarium species, particularly molecular level from Iran. This is the first comprehensive report on identification of many Fusarium spp. from maize in west of Iran.
ACKNOWLEDGMENTS
Author acknowledges the Islamic Azad University, Sanandaj Branch, Iran and University Sains Malaysia, Penang, Malaysia for providing necessary facilities to carry out this research.
REFERENCES
- Aliakbari, F., M. Mirabolfathy, M. Emami, S.F. Mazhar and R.K. Osboo, 2007. Natural occurrence of fusarium species in maize kernels at gholestan province in Northern Iran. Asian J. Plant Sci., 6: 1276-1281.
CrossRefDirect Link - Fandohan, P., K. Hell, W.F.O. Marasas and M.J. Wingfield, 2003. Infection of maize by Fusarium species and contamination with fumonisins in Africa. Afr. J. Biotechnol., 2: 570-579.
Direct Link - Fisher, N.L., L.W. Burgess, T.A. Toussoun and P.E. Nelson, 1982. Carnation leaves as a substrate and for preserving cultures of Fusarium species. Phytopathology, 72: 151-153.
CrossRefDirect Link - Fisher, N.L., W.F.O. Marasas and T.A. Toussoun, 1983. Taxonomic importance of microconidial chains in Fusarium section Liseola and effects of water potential on their formation. Mycologia, 75: 693-698.
Direct Link - Ghiasian, S.A., P. Kord-Bacheh, S.M. Rezayat, A.H. Maghsood and H. Taherkhani, 2004. Mycoflora of Iranian maize harvested in the main production areas in 2000. Mycopathologia, 158: 113-121.
CrossRefDirect Link - Lew, H., A. Adler and W. Edinger, 1991. Moniliformin and the European maize borer (Ostrinia nubilalis). Mycotoxin Res., 7: 71-76.
CrossRef - Logrieco, A., G. Mule, A. Moretti and A. Bottalico, 2002. Toxigenic Fusarium species and mycotoxins associated with maize ear rot in Europe. Eur. J. Plant Pathol., 108: 597-609.
Direct Link - Miller, J.D., 2001. Factors that affect the occurrence of fumonisin. Environ. Health Perspect., 109: 321-324.
Direct Link - Mohammadi, A., F. Nejad and V. Minasian, 2002. Genetic diversity in population of Fusarium moniliforme isolate from sugarcane and corn seeds in Khuzistan using vegetative compatibility groups and relation of VCGs to the pathogenecity. Proceedings of 15th Iranian Plant Protection Congress, Sept. 7-11, Kermanshah, Iran, pp: 135-135.
- Munkvold, G.P., 2003. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol., 109: 705-713.
CrossRefDirect Link - Nelson, P.E., M.C. Dignani and E.J. Anaissie, 1994. Taxonomy, biology, and clinical aspects of Fusarium species. Clin. Microbiol. Rev., 4: 479-504.
Direct Link - Nirenberg, H.I. and K. O'Donnell, 1998. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia, 90: 434-458.
Direct Link - O`Donnell, K., 1996. Progress towards a phylogenetic classification of Fusarium. Sydowia, 48: 57-70.
Direct Link - Thomas, M.D. and I.W. Buddenhagen, 1980. Incidence and persistence of Fusarium moniliforme in symptomless maize kernels and seedlings in Nigeria. Mycologia, 72: 882-887.
Direct Link - Velluti, A., S. Marin, R. Gonzalez, A.J. Ramos and V. Sanchis, 2001. Fumonisin B1, zearalenone and deoxynivalenol production by Fusarium moniliforme, F. proliferatum and F. graminearum 713 in mixed cultures on irradiated maize kernels. J. Sci. Food Agric., 81: 88-94.
CrossRef - Wilson, T.M., P.E. Nelson and C.R. Knepp, 1985. Hepatic neoplastic nodules, adenofibrosis, and cholangrocarcinomas in male Fisher 344 rats fed maize naturally contaminated with Fusarium moniliforme. Carcinogenesis, 6: 1155-1160.
Direct Link - Nicolaisen, M., S. Suproniene, L.K. Nielsen, I. Lazzaro, N.H. Spliid and A.F. Justesen, 2009. Real-time PCR for quantification of eleven individual Fusarium species in cereals. J. Microbiol. Methods, 76: 234-240.
CrossRef