
Hashem Al-Sheikh
Department of Biology, College of Science, King Faisal University, AlHassa, Saudia Arabia
H.M.A. Abdelzaher
Department of Botany and Microbiology, Faculty of Science, Minia University 61519, El-Minia City, Egypt
Journal of Biological Sciences
Year: 2010 | Volume: 10 | Issue: 3 | Page No.: 178-189







ABSTRACT
This research was undertaken to study the biological control of Pythium spinosum var. spinosum, the damping-off organism of soybean using three selected soil fungi of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii. Pythium spinosum var. spinosum was isolated from rhizosphere soil and rhizoplane of healthy and infected soybean roots cultivated in an agricultural field located in Shahean district, Minia city, Egypt in June of 2003 and 2009 using NARM (Nystatin Ampicillin Rifampcin Miconazole) selective medium. Rhizosphere and rhizoplane mycoflora isolated from the same sites were tested for their antagonism against Pythium spinosum in agar plates. Among the isolated fungi, Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii were proved to be potent biological agents in the plate assay. Their field effectiveness was evaluated in either autoclaved or nonsterilized soil. Coating soybean seeds and roots with spores and mycelia of these three antagonists gave germinating seeds and seedlings a very good protection from root-rot, pre- and post-emergence damping-off caused by P. spinosum. Applying these biocontrol agents to autoclaved and nonsterilized soil infested with P. spinosum provided an excellent way of protection.
PDF Abstract XML References Citation
How to cite this article
Hashem Al-Sheikh and H.M.A. Abdelzaher, 2010. Isolation of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii from Agricultural Soil and their Biological Activity Against Pythium spinosum, the Damping-Off Organism of Soybean. Journal of Biological Sciences, 10: 178-189.
DOI: 10.3923/jbs.2010.178.189
URL: https://scialert.net/abstract/?doi=jbs.2010.178.189
DOI: 10.3923/jbs.2010.178.189
URL: https://scialert.net/abstract/?doi=jbs.2010.178.189
INTRODUCTION
Recently, soybean (Glycine max L.) is a strategically important crop in Egypt, as it is grown in a wide area, especially, the upper part of the country. Soybean is a summer crop, cultivated in late May and harvested in early September. High seed yields are attained in Minia governorate with high summer temperatures. High production and income compared with other crops including maize, as well as being fairly disease resistant, have encouraged farmers to grow soybeans to an increasing extent in the last two decades in most cropping areas of upper Egypt.
Soybeans are also well-suited to rotation with a wide range of summer crops. One of many advantages of planting soybeans as a rotation crop includes the fact that they are nitrogen-fixers, thus improves soil nitrogen status and saves fertilizer costs on successive rotations.
Soybeans are an important source of high protein and vegetable oil for human consumption and livestock feed. They are also used as meat extenders in hamburgers, sausages, pet foods and vegetarian products etc. In soybean protein, ten amino acids essential for human nutrition are contained in more nearly ideal proportions than in any other vegetable oil (Caldwell and McLaren, 2004).
In the summer of 2003, symptoms of damping-off of soybean appeared in a field at Shaheen district, Minia, Egypt. Diseased seedlings were stunted and the foliage turned light green to yellow and eventually desicated. Some plants showed damping-off in which the fungus attacked the basal part of the stems, eventually led to collapse of the plant on the soil surface. Same symptoms appeared in the same field in the summer of 2009.
Several studies have been made to characterize the fungal flora of soil adhering to the roots of numerous growing or cultivated plants in many parts of the world (Abdel-Hafez et al., 1990, 1995; Mazen and Shaban, 1983; Nagaraja, 1991; Rajendra and Saxena, 1991).
Microbial antagonism have been used with some success for controlling damping-off diseases of plants. Cliquet and Scheffer (1996) and Abd El-Haleem et al. (2001) reported that damping-off of many green house-grown crops induced by Rhizoctonia solani and Pythium ultimum was appreciably reduced by suing Trichoderma sp. Isolates of Trichoderma sp. often showed antagonism to Pythium ultimum, P. debaryanum, P. aphanidermatum, P. splendens and Pythium sp. (Bolton, 1978; Johnson, 1954; Knauss, 1976). Pythium was also suppressed by a number of fungi such as Aphanomyces, Aspergillus, Chaetomium, Myrothecium, Penicillium and Laetisaria (Joshi and Keshwal, 1970; Chamswarng and Gesnara, 1988; Abdelzaher et al., 2000). Cotton seeds treated with suspensions of Trichladium sp., Penicillium sp. and some bacteria significantly reduced Pythium infection (Chamswarng and Gesnara, 1988). Maghazy et al. (2008) found that species of Aspergillus, Penicillium and other genera of soil fungi gave significant control of alfalfa root-rot caused by Pythium spinosum. Damping-off of wheat seedlings was controlled by dusting the grains with spores of Gliocladium roseum and Trichoderma harzianum (Abdelzaher, 2006).
The aim of this research is to study the antagonistic effects of fungi isolated from the rhizosphere of soybean plants against the damping-off fungal pathogens which cause severe loss to soybean and to study the possibility of employing the antagonistic ones for disease control.
MATERIALS AND METHODS
Isolation of soil fungi from rhizosphere and rhizoplane of soybean plants: Rhizosphere soil of soybean plants was collected by brushing the adjacent soil of the root systems of 100 healthy and diseased plants growing in a field at Shahean district, Minia city, Egypt during June of 2009. From these soil samples, fungi were isolated by the dilution plate method as described by Abdel-Hafez et al. (1990) and with some modification of Maghazy et al. (2008). Glucose-Czapek's agar medium supplemented by chloramphinicol (0.5 mg L-1) and rose bengal (30 μg L-1) was incubated at 28°C for fungal isolation. Five replications were performed for each sample. The developing fungi were identified and the number of fungi was calculated per gm dry soil.
For isolation of rhizoplanic fungi from healthy and diseased soybean plants roots were subjected separately to a series of washings with sterile distilled water then throughly dried between sterile filter papers. The roots were cut into equal segments (5 mm) and the pieces were placed on glucose-Czapek's agar medium supplemented with chloramphenicol (0.5 mg L-1) and rose-bengal (30 ppm). Five plates were used for each sample which and the developing fungi were counted and identified (Booth, 1977; Domsch and Gams, 1972; Domsch et al., 1980; Ellis, 1971; Klich, 2002; Raper and Thom, 1949; Raper and Fennel, 1965; Sivanesan, 1984). The numbers were calculated per 400 root segments.
Isolation of Pythium species from rhizosphere, rhizoplane and infected soybean plants: Two methods were employed for isolation of Pythium species from soil samples collected from soybean roots as previously described (Abdelzaher, 1999):
| • | Particles of soil were placed on Petri-dishes containing (NARM) medium (Senda et al., 2009) for selective isolation of Pythium species. The emerging hyphal tips were transferred to water agar medium for further purification from bacterial contamination |
| • | The usual method of setting baits was employed. Autoclaved segments of Echinochloa colonum, leaf blades and autoclaved cucumber seeds were used as baits. Five grams of soil were placed each on a Petri-dish. Ten milliliter of sterilized distilled water (Abdelzaher et al., 1995) were added to enable the baits to float on the surface. After five days of incubation at 25°C, the baits were removed, washed thoroughly with sterilized distilled water and blotted dry with sterile filter paper. Four baits were then placed on the edge of a Petri-dish containing NARM medium for isolation of Pythium sp., selectively. The baits were incubated at 20°C for three days or until the appearance of colonies. The emerging hyphal tips were transferred to water agar medium for further purification from bacterial contamination |
Soybean roots were washed thoroughly in running tap water for 30 min followed by sterilized distilled water and then blotted dry with sterile filter paper. Four root pieces were placed on the edge of a Petri-dish containing NARM medium and incubated at 20°C for three days. Isolation of Pythium sp. growing from each piece was performed by cutting a small block of agar medium from the distal end of the colony and transferring to water agar medium for further purification.
Infected soybean roots were washed thoroughly in running tap water for 30 min, rinsed for 3 min in 70% ethanol then washed by sterilized distilled water and blotted dry with sterile filter paper. Four pieces of each of diseased root and infected basal parts of the stem were placed on the edge of a Petri-dish containing NARM medium and incubated at 20°C for three days. Isolation of Pythium sp. growing from each piece was performed by cutting a small block of agar medium and transferring to water agar medium for further purification. A thin piece containing a single hyphal tip was taken from the margin of the surface of the Pythium colony and transferred to a Corn Meal Agar (CMA) slant. The slant culture was checked for bacterial contamination by transferring the CMA block with the fungus to a tube containing Nutrient Broth (NB) medium and incubating at 28°C for 48 h.
Identification: For purification and morphological identification of isolated pythia, the NARM selective medium was found to be effective in inhibiting the development of bacteria whilst not affecting Pythium species. Morphological identification was done with the Key of Van der Plaats-Niterink (1981) and Dick (1990) were principally used for identification. Keys and descriptions by Waterhouse (1967), Middleton (1943) and Abdelzaher (1999) were also consulted for comparison or confirmation of identification.
To induce zoospore formation, the colonized Echinochloa colonum leaf blades by Pythium spinosum were put in Petri-dishes (7 cm diam) containing 10 mL of sterilized distilled water and incubated at different temperatures (5, 10, 15, 20, 25, 30 and 35°C). For obtaining sexual structures, cultures were grown on V-8 juice agar, corn meal agar, potato dextrose agar each supplemented with 500 μg mL-1 of wheat germ oil. For Pythium group G, cross mating with each isolate and with other isolates within the same group was unsuccessful. Thirty measurements were made for each structure of each isolate whenever possible.
DNA extraction: Mycelia were grown in V8 agar medium at 25°C for 7 days or until adequate growth was observed. To extract the total genomic DNA, mycelia from the edge of Pythium colony from a culture plate were suspended in 200 μL of PrepMan Ultra Sample Preparation Reagent (Applid Biosystems, CN, USA) in a 2.0 mL microcentrifuge tube. Samples were vortexed for 10 to 30 sec and then heated for 10 min at 100°C in dry thermo unit or water bath. Samples were spin for 30 min at 15000 g. Supernatants were transferred into a new microcentrifuge tubes and were ready for PCR (Senda et al., 2009).
DNA amplification and sequencing: The nuclear rDNA region of the internal transcribed spacer (ITS), including the 5.8S rDNA, was amplified with the universal primers ITS4 (5´ TCCTCCGCTTATTGATATGC 3´) and ITS5 (5´ GGAAGTAAAAGTCGTAACAAGG 3´). Depending on the experiment, primers of ITS1 (5´ TCCGTAGGTGAAC CTGCGG3´) and ITS2 (5´GCTGCGTTCTTCATCGATGC 3´) were used as described by White et al. (1990) and Matsumoto et al. (1999). The amplicons were 700-900 bp long. On the other hand, 563 bp of the cox II gene was amplified in certain pythia with the primer pair FM66 (5´TAGGATTTCAAGATCCTGC3´) and FM58 (5´CCACAAATTTCACTACATTGA 3´) (Martin, 2000).
Amplification of the sequencing template was carried out with DNA Thermal Cycler 2700 (Applied Biosystems) with a cycling profile of pre-PCR at 94°C for 5 min, followed by denaturation at 94°C for 1 min, primer annealing at 55°C for ITS, 52°C for cox II and elongation at 72°C, 2 min for 40 cycles, with a 7 min extension at 72°C after the final cycle. To check the presence of PCR products, 5 μL of the PCR reaction mixture was loaded in 2% L03 (Takara Bio) agarose gel, electrophoresed at 100 V, 20-30 min and stained with ethidium bromide. The sequencing templates were purified with GenElute PCR Clean-up kit (Sigma Chemical Co., St Louis, Missouri, USA) following the manufacturer’s instructions. Sequencing was performed with BigDye Terminator v3.1 Cycle Sequencing Reaction kit (Applied Biosystems) using the same primers in the initial PCR step. After purifying the sequencing reaction mixture through ethanol precipitation it was run on ABI 3100 DNA Sequencer (Applied Biosystems).
Antagonism between Pythium species and the isolated fungi on agar plates: Interactions between Pythium species and some of the isolated fungal species were tested in Potato-Dextrose Agar (PDA) plate. A disc containing the tested fungus was placed near the periphery of the Petri-dish. Pythium disc was placed at the other side of the dish at a distance of 5 cm from the tested fungus. Usually, the antagonist was placed 4-10 days before Pythium. For a fast-growing fungus, the antagonist and Pythium were placed at the same time. After inoculation, all plates were incubated at 25°C for 1 week before determination of inhibition of mycelial growth.
Pre- and post-emergence damping-off tests: For preparation of inocula of Pythium, a new method was developed. Two concentrations of 2.5 and 20% inocula concentrations and those of 5 and 20% inoculums concentration were used for pre-emergence damping-off test and post-emergence-damping-off experiment, respectively. Five grams of grass blade leaf segments (0.5x1 cm) and 2 g of glucose were moistened by adding distilled water (10 mL) each in 250 mL Erlenmeyer flask. After autoclaving at 121°C for 20 min, each flask was inoculated with three disks (7 mm dam) of agar inoculated with growing margins of Pythium sp. obtained from CMA culture medium. The inoculated conical were held at 25°C for 10 days. In pre-emergence damping-off tests, two inoculums concentrations of 2.5 and 20% were performed. The inoculum concentration of 2.5%, was obtained by mixing thoroughly the 1 g of colonized grass leaf segments in the Erlenmeyer flask with 50 g of oven-dried (70-80°C for 2 days) clay loam soil using a sterilized mortar and pestle. Two point five gms of this mixture were added to 97.5 g of clay loam soil which had been sterilized by autoclaving at 121°C for 60 min (pH 7.1 after autoclaving) and kept in plastic bag for 2-3 weeks at room temperature with 25% water content prior to use. To prepare 20% inoculums density, twenty g of this mixture were added to 80 g of clay loam soil which had been sterilized by autoclaving at 121°C for 60 min (pH 7.1 after autoclaving) and kept in plastic bag for 2-3 weeks at room temperature with 25% water content prior to use. Healthy and diseased seedlings were counted at regular intervals until the development of 2 true leaves in non-inoculated control. Pre-emergence damping-off was determined as difference in emergence between healthy control soil and diseased soil.
Inoculated soil (100 g) each were distributed into 10 replicate pots (100 mL capacity and 7.5 diam) and six soybean seeds were planted in each pot for pre-emergence damping-off test, whereas in case of post-emergence damping-off experiments the inocula were added to the soil after the seedling emergence.
Pre-emergence damping-off was determined as the difference in seedling emergence between healthy control soil and infested soil. Post emergence damping-off was determined from the number of diseased plants as a percentage of the emerged ones. The experiments were carried out in a growth illuminated cabinet (Precision, USA) at 30°C with 12 h photoperiod (91 μmol-2/sec) under humid condition (Abdelzaher et al., 2000).
Seed coating with tested fungi: Mycelial mats of the tested fungi were obtained from cultures grown in Potato dextrose liquid medium. Aliquots of 20 mL medium were dispensed in 9-cm-diam Petri-dishes and each was inoculated with a single (0.5-cm-diam) disc cut from the margin of the fungal colony grown on PDA. The cultures were incubated for 10 days at 28°C in darkness. Mycelial mats were then removed, washed in sterile distilled water and fragmented in distilled water using a tissue homogenizer. This preparation of mycelial fragments and conidia was stored at 5°C until use. This preparation was used to coat soybean seeds. Equal volumes (100 mL) of 3% solution of carboxymethylcellulose (CMC) and the preparation were mixed and about 100 seeds of surface sterilized soybean were added. After being allowed to soak for 2 min in the preparation, the seeds were removed, spread in sterile open Petri-dish and allowed to dry overnight in a laminar flow cabinet at room temperature. All seeds were then used immediately in damping-off experiments (Al-Hamdani et al., 1983).
Testing effect of fungal cultural filterates on soybean seed germination: In order to use fungi as biocontrol agents, they must not produce any metabolic toxins that retard plant growth and seed germination. In each sterilized Petri-dish, sterilized filter paper (9 cm in diam) was placed and moistened with 10 mL filter sterilized cultural filterates of each of the tested fungi. Twenty soybean seeds were placed in each dish and were incubated at 25°C until seeds germinated. Ten milliliter sterilized distilled water was used as control (Abdelzaher, 2006).
Effect of tested fungi on infection of soybean seedlings by Pythium species: Testing inhibition activity of fungi on infection of soybean seedlings by Pythium spinosum was performed using soybean seeds coated with tested fungi as well as mixing inocula of these fungi with infested soil with P. spinosum in case of pre-emergence damping-off and inoculum mixture of antagonistic fungi and Pythium species in case of post-emergence damping-off test (Abdelzaher, 2006).
RESULTS
Fungi from rhizosphere and rhizoplane of soybean plants: Thirty nine species in addition to 1 species variety belonging to 15 genera were isolated from the rhizosphere of soybean plants on glucose-Czapek's agar at 28°C. Aspergillus was the most common genus and was recovered from all samples representing 55.6% of total fungi. It was represented by 10 species of which A. niger, A. flavus, A. terreus and A. fumigatus were the most common. They were recovered from all samples comprising 22.4, 20.8, 18.6 and 17.2% of total Aspergillus and 12.5, 11.5, 10.3 and 9.6% of total fungi, respectively. Aspergillus versicolor, A. ustus, A. flavipes and A. sulphureus were less frequent. These species were frequently encountered in various types of Egyptian soils as reported by Abdel-Fattah et al. (1977), Moubasher and Abdel-Hafez (1978), Mazen and Shaban (1983) and Abdel-Hafez et al. (1990, 1995) (Table 1).
Penicillium ranked second in the frequency of isolation. It emerged in 90% of the sample constituting 14.8% of total fungi. Of the genus 8 species were collected of which P. chrysogenum was the most prevalent. It was recovered from 70% of the samples matching 66.5% of total Penicillium and 9.9% of total fungi. Penicillium aurantiogriseum, P. funiculosum and P. citrinum were isolated moderately, but P. purpurogenum, P. islandicum and P. islandicum were isolated in low and rare occurrence, respectively. Abdel-Hafez et al. (1995) listed Penicillium chrysogenum among the most common fungi in the soil of sugar cane fields at Qena governorate, Egypt. Mazen and Shaban (1983) reported that P. citrinum, P. chrysogenum, P. cyclobium (= P. aurantiogriseum), P. jensenii, P. nigricans (= P. janczewskii) were the most common Penicillium species in wheat field in Egypt (Table 1). Trichoderma ranked third in the frequency of isolation.
| Table 1: | Total fungal counts from rhizosphere (per g dry soil) and rhizoplane of soybean roots [per 400 root (5 mm) segments], number of cases of isolation (NCI; out of 20 cases), occurrence remarks (OR) and percentage of total counts (TC%) of various fungal genera and species recovered from maize plant on Czapek's agar at 28°C |
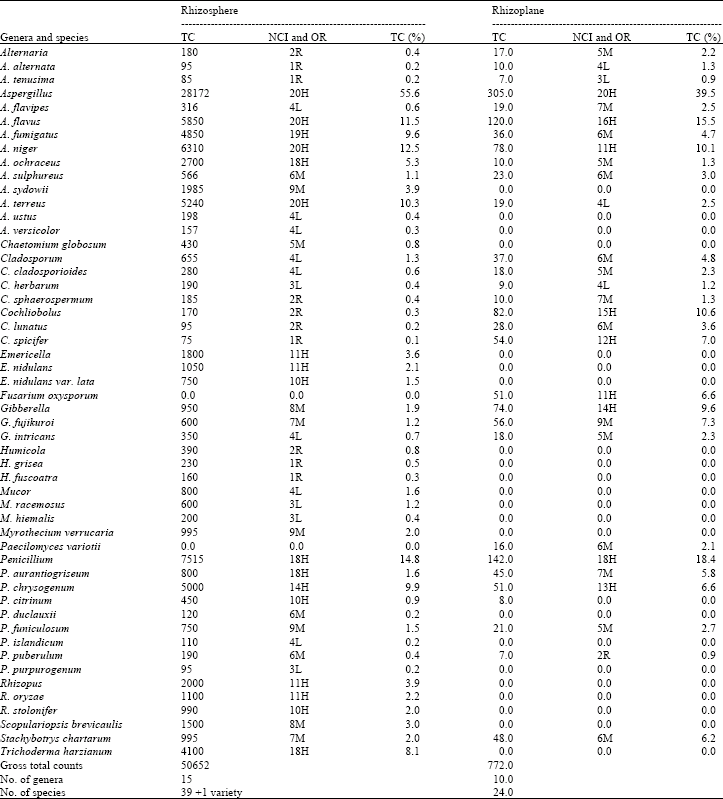 | |
| Occurrence remarks: H = High occurrence, between 10~20 cases (out of 20); M = Moderate occurrence, between 5~9 cases; L= low occurrence, 3 or 4 cases; R = Rare occurrence, 1 or 2 cases | |
It emerged in 90% of the sample constituting 8.1% of total fungi. It was represented by T. harzianum. Rhizopus and Emericella occupied the fourth place among the most common genera. They emerged in 55% of the samples matching 3.9 and 3.6% of total fungi (Table 1).
Myrothecium verrucaria, Scopulariopsis brevicaulis and Stachybotrys chartarum were isolated with moderate frequency of occurrence; emerging from 30-45% of the samples constituting 2-3% of total fungi. The remaining genera and species were isolated in low or rare frequency of occurrence (Table 1).
It is worth mentioning that all soil fungal species recovered in the present investigation were isolated previously, but with different numbers and incidence, from the rhizosphere of several cultivated plants in Egypt (Abdalla and Omar, 1981; Abdel-Fattah et al., 1977; El-Hissy et al., 1980; El-Abyad et al., 1982; Abdel-Hafez et al., 1990, 1995; Gherbawy and Abdelzaher, 1999).
Twenty-four species representing 10 genera of fungi were isolated from the rhizoplane of soybean plant on glucose-Czapek's agar at 28°C (Table 1).
Aspergillus flavus, A. niger, Cochliobolus spicifer, Fusarium oxysporum, Gibberella fujikuroi and P. chrysogenum were the most common fungal species on rhizoplane of soybean plants. They emerged in 55-80% matching 6.6-15.5% of total fungi. Aspergillus flavipes, A. fumigatus, A. terreus, Chaetomium globosum, Cladosporum cladosporioides, Cochliobolus lunatus, Gibberella intricans, Paecilomyces variotii, Penicillium aurantiogriseum, P. funiculosum and Stachybotrys chartarum were isolated in moderate occurrence representing 25-40% of the samples and constituting 2.2-6.2% of total fungi. The remaining genera and species were isolated in low and rare frequencies of occurrence (Table 1).
Most of the above fungal species were previously isolated from rhizoplane of healthy cotton (Moubasher and Abdel-Hafez, 1986), broad bean (Abdel-Fattah et al., 1977), lentil (Abdel-Hafez et al., 1990), wheat (Abdel-Hafez, 1982), sugar cane (Abdel-Hafez et al., 1995) and maize (Abdelzaher et al., 2000).
Occurrence of Pythium species in rhizosphere and rhizoplane and infected soybean plants: Results of Pythium isolation from rhizosphere soil of soybean plants indicated that, Pythium spinosum and Pythium group G were present in soil adhering to the soybean roots grown in the field studied.
Pythium spinosum was the only Pythium species which isolated from soybean rhizoplane and it was the only taxa isolated from necrotic soybean roots.
Pythium spinosum sawada: Colonies on Bacto-CMA have some cottony aerial mycelium. Main hyphae are up to 7 μm wide and septate when old (Fig. 1a, b, d, e). Zoospores are not formed at temperatures from 5 to 30°C. Hyphal swellings are usually lemon-form, rarely globose, up to 26 μm in diam and sometimes with projections (Fig. 1b). Oogonia are globose, rarely fusiform, usually intercalary, 23-28 μm in diam, 25 μm diam on average, with a varying number of blunt, digitate projections, 2-12 μm long and 2-4 μm wide at the base (Fig. 1b-f). Antheridia are 1 (-2) per oogonium, monoclinous and sometimes diclinous (Fig. 1, b-e). Oospores are plerotic, occasionally aplerotic, 19-25 μm in diam, 21 μm diam on average and their walls are up to 2 μm thick (Fig. 1e, f).
Pythium group G: Main hyphae are up to 7 μm wide. Zoosporangia are globose often 20-30 μm wide, 25 μm diam on average with evacuation tubes usually short up to 50 μm long, sometimes germinating by one or two germ tubes. Zoospores are produced at 20°C, after 24 h; encysted zoospores are 8-10 μm in diam and germinate mostly by one germ tube. No sexual reproduction has been observed.
Molecular identification: Sequencing of rDNA-ITS including the 5.8 S rDNA were analyzed for the Pythium species tested by the method of Kageyama et al. (2005) to confirm the species identification. The sequence of (El-U1199) was closely related with that of P. spinosum var. spinosum (AY598701.1) with 100% similarity.
Sequence of r-DNA-ITS including the 5.8 S rDNA for Pythium spinosum var. spinosum (El-U1199):
CCACACCTAAAAAACTTTCCACGTGAACTGTCATTATTTGTTGTGTGTCTGTGCGTTGTTGGCGTGCATTTGCTTACACTTTGGTGTTTGTGAGT
GTGTGTTGGCAGTGTGCGGACTGAACGAAGGTTGTGTGTTGTTATGTGCCTACTGCACTGCTGACTTTGCATTCATTTGTATGGTCTTGGCGGA
GTGGCGGGTACTGTGCATGCGCAACTGACTTATTTTTTCAAACCCCATACCTAAATGACTGATTATACTGTGAGAACGAAAGTTCTTGCTTTAA
ACTAGATAACAACTTTCAGCAGTGGATGTCTAGGCTCGCACATCGATGAAGAACGCTGCGAACTGCGATACGTAATGCGAATTGCAGAATTCA
GTGAGTCATCGAAATTTTGAACGCATATTGCACTTCCGGGTTATACCTGGAAGTATGTCTGTATCAGTGTCCGTAAATCAAACTTGGCTTTCTTC
CTTCCGTGTAGTCGGTGGAGGAGAGTTGCAGATGTGAAGTGTCTCGCTATGGTTGGCATTTGTAATGAATGCACAGCTTGCGAGTCCTTTTAAAT
GGACACGACTTTCTCTTTTTTGTATCTGCGTGGTGCTGTGTATGAACGCGGTGGTTTTCGGATCGCTCGCGGCTATCGGCGACTTCGGTGAATGC
ATTATGGAGTGGACCTCGATTCGCGGTATGTTGGGCTTCGGCTGGACAATGTTGCTTATTGTGTGTTTGTCCGTGTTCGCCTTGAGGTGTACTGG
TAGTTGTGGGATTGAACTGGTTACTGTTGTTAGTAGTGTGTAGTGCGTTGTCGTGGATGCATCTGTCTTTTGTGCACTTTTGTGTGTGCAGTTGAT
AGAAGAGGAGTTTGAATTTGGGAAATTAGTGTACTGTGGGTTAATCCTGCGTGTATATCTCAA
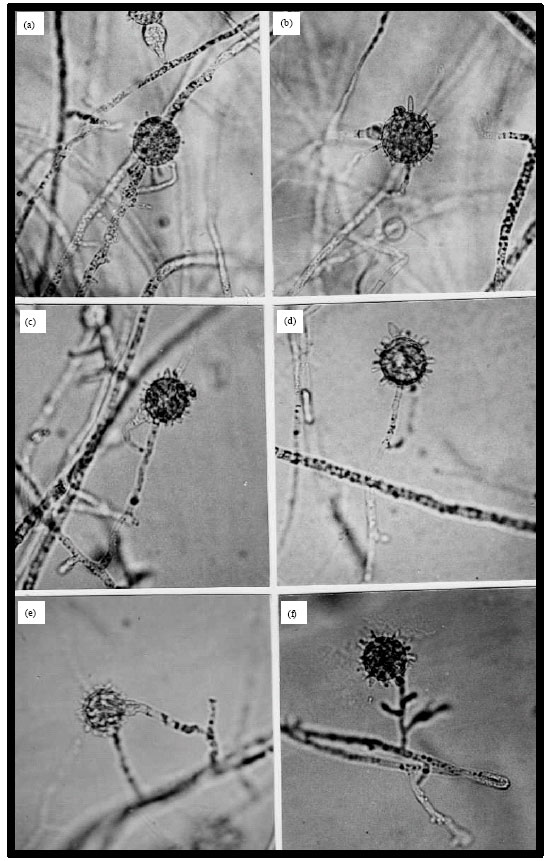 | |
| Fig. 1: | Morphology of Pythium spinosum El-U1199. (a, b, e) Mycelia.1. Hyphal swellings. (c, d, f) Monoclinous antheridia. 4. A diclinous antheridium. 6. A plerotic oospore. Bar (20 µm) on photo d is applicable to the rest of photos |
Description: Based on El-U1199 (deposited at the Department of Botany and microbiology, Faculty of Science, Minia University, Egypt). Molecular identification was performed by Dr. Hashem Al-Sheikh.
Isolation: On NARM medium from infected soybean roots in a field in Shahean district, Minia city, Egypt, 10 June 2009, by Hani M. Awad Abdelzaher.
Cardinal temperatures: Minimum 5°C, optimum 28°C, maximum 37°C. Daily growth rate on CMA at 25°C = 33 mm.
Pathogenicity tests
Pre- and post-emergence damping-off tests: Three isolates of Pythium spinosum and 2 isolates of Pythium group G were used. Pre- and post-emergence damping-off experiments revealed that P. spinosum isolates were highly virulent to soybean germinating seeds as a pre-emergence damping-off pathogens. Isolate III of P. spinosum was proved to be highly virulent fungus as a post-emergence damping-off parasite (at 5 and 25% inoculum density) whereas isolate I and isolate II of P. spinosum were moderetly virulent at 5% inoculum density and highly pathogenic at 25% inoculum density against soybean seedlings. Isolates of Pythium group G were avirulent fungi (Fig. 2, 3).
Antagonism between P. spinosum and Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii on agar plates: Measuring inhibition zones around each tested fungus grown against isolates of P. spinosum indicated that these fungi inhibit Pythium growth on PDA medium. A. sulphureus was the strongest followed by Paecilomyces variotii and P. islandicum showing average of 31, 25 and 19 mm inhibition zones against isolates of Pythium spinosum, respectively, when grown on PDA medium at 25°C.
Effect of fungal cultural filtrates on soybean seed germination: Results indicated that there were no obvious effects of the cultural filtrates on soybean seed germination.
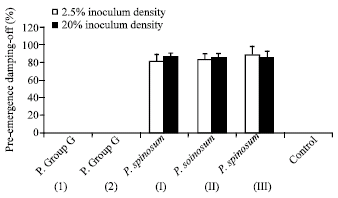 | |
| Fig. 2: | Pre-emergence damping-off of soybean grown in clay sand soil infested with two inoculum densities of isolates of Pythium spinosum and isolates of Pythium group G. Bars indicate standard deviations of three replicates |
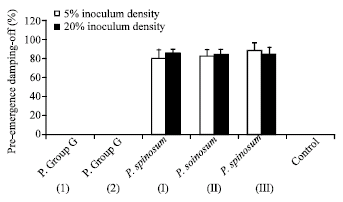 | |
| Fig. 3: | Post-emergence damping-off of soybean grown in clay sand soil infested with two inoculum densities of isolates of Pythium spinosum and isolates of Pythium group G. Bars indicate standard deviations of three replicates |
Effect of adding tested fungi on the pre- and post-emergence damping-off by P. spinosum: Pre-emergence damping-off of soybean was prevented when soybean seeds were either coated by Aspergillus sulphureus, Penicillium islandicum or Paecilomyces variotii or mixing them with infested soil by P. spinosum before planting. Emergence of soybean seedlings from not coated seeds with Pythium spinosum was reduced by 100% in case of using autoclaved soil and 98% in case of nonsterilized soil when planted in soil infested by P. spinosum. Seeds coated with 3% CMC only and planted in sterile soil gave 100% germination and emergence (Table 2).
In post-emergence damping-off experiments, soil infestation by both P. spinosum and any of the tested fungi reduced damping-off and gave an excellent way of protection. Dipping seedlings, root system and basal part of the stem, in inoculum preparation of each antagonist with the aid of 3% CMC and then transplanting in an infested soil also protect the plant from post-emergence damping-off (Table 3).
| Table 2: | Effect of adding Pythium spinosum and antagonistic fungi (Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii), on pre-emergence damping-off of soybean seedlings |
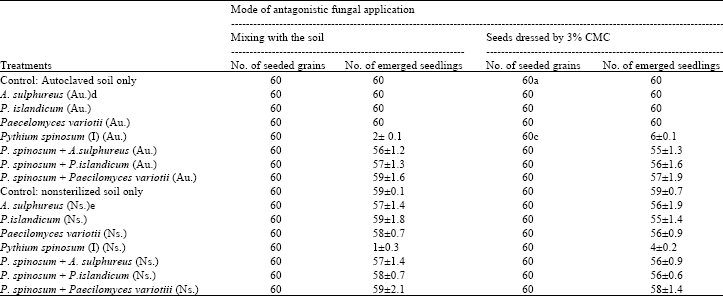 | |
| a: Soybean seeds were dressed by 3% CMC only before transplanted. b: Standard error for three replicates. c: Maize grains were dipped in 3% CMC before seeding in soil infested with P. spinosum. d: Autoclaved soil. e: Nonsterilized soil | |
| Table 3: | Effect of adding Pythium spinosum and antagonistic fungi (Aspergillus sulphureus, P. islandicum and Paecilomyces variotii), on post-emergence damping-off of soybean seedlings |
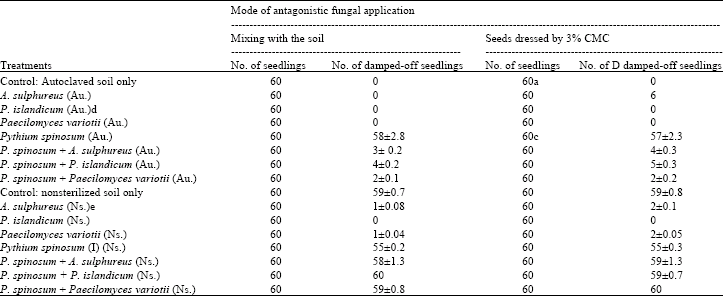 | |
| a: Soybean seedlings (dressing include basal part of the stems) were dressed by 3% CMC only before transplantation. b: Standard error for three replicates. c: Soybean seedlings (dressing include basal part of the stems) were dipped in 3% CMC before seeding in soil infested with P. spinosum. d: Autoclaved soil. e: Nonsterilized soil | |
DISCUSSION
Pythium spinosum is a cosmopolitan species and has a wide host range (Van der Plaats-Niterink, 1981). It infects the seeds and young roots of a broad range of plant species and is found in nearly every country of the world (Abdelzaher, 1999).
Isolates of P. spinosum investigated in this study were pathogenic to germinating seeds and seedlings of soybean. The fungus was isolated from soil and infected soybean roots growing in a field near Minia city, Egypt. It was observed that, on aging of the infected soybean plants, necrosis of many feeder roots occurred. Feeder roots necrosis is known to be a major factor limiting corn production (Bouhot, 1988). In Egypt, so far, there is no effective control method for this fungus to the farmers, although, this pathogenic microorganism is distributed easily from field to field. Therefore, its control is of importance and, it is urgent to find a way to overcome the disease. A major objective of this study was to isolate potential fungi from soil which might be effective in controlling the disease.
Few successful attempts to control diseases in nonsterile, naturally infested soil have been reported. Slagg and Fellows (1947) reduced infection of Ophiobolus graminis by adding Aspergillus flavus to soil naturally infested with the pathogen. Tveit and Moore (1954) found that antagonistic isolates of Chaetomium added to naturally or artificially infested nonsterile soil reduced infection of oat seedlings by Helminthosporum victoriae. The saprophytic ascomycete, Chaetomium globosum is a potential antagonist of Pythium ultimum, the causal agent of damping-off of sugarcane (Pietro et al., 1992), Abdelzaher et al. (2000) postulated that Aspergillus terreus, Myrothecium verrucaria and Chaetomium globosum adhered to maize grains enhance emergence when cultivated in soil infested with Pythium deliense.
Among the soil fungi isolated in this study, three fungal species namely; Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii were chosen according to their antagonism to growth of P. spinosum in agar plates. According to Johnson (1954), only organisms antagonistic to the pathogen in agar culture were promising to control Pythium root rot of corn. Also, a positive relationship between the decrease in severity of disease in soil and the magnitude of the zone inhibition of the causal fungus in Petri-dish culture was reported by Abdelzaher et al. (2000), Johnson (1954), Maghazy et al. (2008) and Stevenson (1956). Therefore, Aspergillus sulphureus, penicillium islandicum and Paecilomyces variotii were chosen to perform biological control tests according to their effectiveness in agar plates against Pythium spinosum var. spinosum.
Addition of Aspergillus sulphureus, penicillium islandicum and Paecilomyces variotii afforded a very good protection of soybean seedlings from pre- and post-emergence damping-off caused by P. spinosum. The effectiveness of these antagonistic fungi in reducing the severity of pre- and post-emergence damping-off of soybean caused by P. spinosum indicated their abilities to inhibit growth of the pathogen and then infection. Mixing the antagonist with the soil resulted in a better protection than dressing roots or grains with the fungus with the aid of CMC. This may suggest the importance of the proliferation of the antagonist in the rhizosphere. Antagonistic fungi with a high density in the rhizosphere produce sufficient substances for controlling the pathogen than dressing them on plant parts since the former provides a continuous colonization of the roots from the bulk soil. In autoclaved soil, pre- and post-emergence damping-off of soybean were more severe than in nonsterile soil and this may be due to competetion from soil microflora. Results here are compatible with previous works in which some soil fungi including Aspergillus, Penicillium and other genera were able to control damping-off diseases caused by some Pythium sp. (Srinivasan, 1968; Abdelzaher et al., 2000; Maghazy et al., 2008).
It should be mentioned that before large scale experiments are performed to control the disease using these antagonists, methods of increasing population of the antagonistic fungi in the natural soil as well as the mode of action of these fungi in biological control of Pythium diseases should be studied.
REFERENCES
- Abdalla, M.H. and F. Omer, 1981. Studies on the rhizosphere mycoflora of Euphorbia granulata Forsk. and E. heterophylla L. Mycologia, 73: 9-15.
CrossRefDirect Link - Abdel-Hafez, S.I.I., 1982. Rhizosphere and rhizoplane fungi of Triticum vulgare cultivated in Saudi Arabia. Mycopathologia, 78: 79-86.
CrossRefDirect Link - Abdelzaher, H.M.A., 2006. Biological control of damping-off and root rot diseases of soybean caused by Pythium spinosum Sawada var. spinosum using Rhizosphere species of Aspergillus sulphureus, Penicillium islandicum and Paecilomyces variotii. Jap. J. Phytopathol., 72: 247-248.
Direct Link - Abdelzaher, H.M.A., T. Ichitani, M.A. Elnaghy, S.K.M. Hassan and E.M. Fadl-Alla, 1995. Materials for Pythium flora of Japan. X. Occurrence, identification and seasonality of Pythium spp. in three pond waters and mud soils in Osaka. Mycoscience, 36: 71-85.
Direct Link - Al-Hamdani, A.M., R.S. Lutchmeah and R.C. Cooke, 1983. Biological control of Pythium ultimum induced damping-off by treating cress seeds with the mycoparasite Pythium oilgandrum. Plant Pathol., 32: 449-454.
Direct Link - Cliquet, S. and R.J. Scheffer, 1996. Biological control of damping-off caused by Pythium ultimum and Rhizoctonia solani using Trichoderma sp. applied as industrial film coatings on seeds Eur. J. Plant Pathol., 102: 247-255.
CrossRefDirect Link - Domsch, K.H., W. Gams and T.H. Anderson, 1980. Compendium of Soil Fungi. Academic Press, London, United Kingdom, ISBN: 9780122204012, Pages: 1264.
Direct Link - El-Hissy, F.T., S.I.I. Abdel-Hafez and M.I.A. Abdel-Kader, 1980. Rhizosphere fungi of five plants in Egypt. Z. Allg. Mikrobiol., 20: 177-184.
Direct Link - Gherbawy, Y. and H.M.A. Abdelzaher, 1999. Isolation of fungi from tomato rhizosphere and evaluation of the effect of some fungicides and biological agents on the production of cellulase enzymes by Necteria haematococca and Pythium ultimum var. ultimum. Czech Mycol., 51: 157-170.
Direct Link - Kageyama, K., A. Nakashima, Y. Kajihara, H. Suga and E.B. Nelson, 2005. Phylogenetic and morphological analyses of Pythium graminicola and related species. J. Gen. Plant Pathol., 71: 174-182.
CrossRefDirect Link - Martin, F.N., 2000. Phylogenetic relationships among some Pythium species inferred from sequence analysis of the mitochondrially encoded cytochrome oxidase II gene. Mycologia, 92: 711-727.
Direct Link - Matsumoto, C., K. Kageyama, H. Suga and M. Hyakumachi, 1999. Phylogenetic relationships of Pythium species based on ITS and 5.8S sequences of the ribosomal DNA. Mycoscience, 40: 321-331.
CrossRefDirect Link - Mazen, M.B. and G.M. Shaban, 1983. Seasonal fluctuations of non-rhizosphere soil fungi in wheat fields in Egypt. Qatar Univ. Sci. Bull., 3: 115-129.
Direct Link - Middleton, J.T., 1943. The taxonomy, host range and geographic distribution of the genus Pythium. Memories Torrey Bot. Club, 20: 1-171.
Direct Link - Moubasher, A.H. and S.I.I. Abdel-Hafez, 1978. Further study on seasonal fluctuation of Egyptian soil fungi. Mycopathology, 63: 11-19.
Direct Link - Srinivasan, K.V., 1968. Effect of a varietal mixture on sugarcane root rot susceptibility and on the rhizosphere microflora. Plant Sci., 68: 150-162.
Direct Link - Stevenson, I.L., 1956. Antibiotic activity of actinomycetes in soil and their controlling effects on root-rot of wheat. J. Gen. Microbiol., 14: 440-448.
Direct Link - White, T.J., T.D. Bruns, S.B. Lee and J.W. Taylor, 1990. Amplification and Direct Sequencing of Fungal Ribosomal RNA Genes for Phylogenetics. In: PCR Protocols: A Guide to Methods and Applications, Innis, M.A., D.H. Gelfand, J.J. Sninsky and T.J. White (Eds.), Academic Press, San Diego, CA, USA, ISBN-13: 9780123721808, pp: 315-322.
CrossRefDirect Link - Lewis, J.A., R.D. Lumsden, 2001. Boicontrol of damping-off of greenhouse-grown crops caused by Rhizoctonia solani with a formulation of Trichoderma spp. Crop Protect., 20: 49-56.
CrossRef