
Z.C. Cob
Marine Ecosystem Research Center, Faculty of Science and Technology, National University of Malaysia, 43600 Bangi, Selangor, Malaysia
A. Arshad
Department of Biology, Faculty of Science, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
J.S. Bujang
Department of Biology, Faculty of Science, University Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
M.A. Ghaffar
Marine Ecosystem Research Center, Faculty of Science and Technology, National University of Malaysia, 43600 Bangi, Selangor, Malaysia
Journal of Applied Sciences
Year: 2009 | Volume: 9 | Issue: 18 | Page No.: 3287-3297







ABSTRACT
The age, growth, mortality and population structure of Strombus canarium Linnaeus, 1758 were examined in the Johor Straits, Malaysia from January to December 2005. A total of 2088 conchs were sampled where females were more abundant than males with monthly sex ratio of 1.72±0.17 (N = 12). The estimated growth parameter showed higher asymptotic length (L∞) and growth coefficient (K) in females (L∞ = 70.20 mm, K = 1.50 year-1) compared with the males (L∞ = 69.30 mm, K = 1.20 year-1). This resulted in better overall growth performance of females (φ’ = 3.81) compared with the males (φ’ = 3.48). The growth pattern of females and males showed positive allometric nature of growth (b>3, p<0.05), with estimated maximum life span of 2.0 and 2.5 year, respectively. The recruitment pattern was continuous, displaying only a single major peak event per year. The estimated natural mortality rate (M) was 0.95 year-1 in females and 0.86 year-1 in males, while the total mortality rate (Z) was 2.56 and 2.72 year-1, respectively. The fishing mortality rate (F) was 1.61 year-1 in females and 1.86 year-1 in males, which were higher than the natural mortality rates, thus indicating an unbalanced position of the stock. In addition, the exploitation rate (E) was higher than the maximum allowable limit of exploitation (EMSY), which was a further indication of overexploitation. For sustainable utilization of the resource, serious efforts should immediately be taken in reducing the exploitation rates of S. canarium in the study area.
PDF Abstract XML References Citation
How to cite this article
Z.C. Cob, A. Arshad, J.S. Bujang and M.A. Ghaffar, 2009. Age, Growth, Mortality and Population Structure of Strombus canarium (Gastropoda: Strombidae): Variations in Male and Female Sub-Populations. Journal of Applied Sciences, 9: 3287-3297.
DOI: 10.3923/jas.2009.3287.3297
URL: https://scialert.net/abstract/?doi=jas.2009.3287.3297
DOI: 10.3923/jas.2009.3287.3297
URL: https://scialert.net/abstract/?doi=jas.2009.3287.3297
INTRODUCTION
The dog conch, Strombus canarium Linnaeus, 1758 is a tropical conch snail that was widely distributed within the coastal areas of the Indo-Pacific region (Poutiers, 1998). In Malaysia, they are very abundant within the seagrass beds of the Johor Straits (Cob et al., 2008a, b, 2009) and constitutes among the largest and most dominant herbivorous mollusks in the area (Cob et al., 2009). The species is one of the most important shellfish traditionally zand currently gathered in Malaysia (Purchon and Purchon, 1981; Poutiers, 1998; Cob et al., 2009) and elsewhere in the Indo-Pacific region (Amini, 1986; Amini and Pralampita, 1987; Erlambang and Siregar, 1995; Erlambang, 1996).
However, the species is relatively less known and not extensively studied as compared to other commercially important mollusks in the region (Amini, 1986; Erlambang and Siregar, 1995; Cob et al., 2009). Information regarding the biology, ecology and fishery of the species is very limited and currently there is no regulation concerning the fishery of the species. Since, it has long been exploited there is urgent needs to assess the status of the population stock, which is crucial for fishery management and sustainable utilization of the resource. Knowledge of various population parameters such as asymptotic length (L∞), growth coefficient (K), mortality (natural and fishing) and exploitation rates (E) are therefore extremely important and urgently needed.
There are many analytical tools that can be applied in assessing the exploitation level and stock status of marine fishery resources. Among them the FiSAT (FAO-ICLARM Stock Assessment Tools) has been most frequently employed in estimating the population parameters (Mancera and Mendo, 1996; Blaber et al., 1998; Al-Barwani et al., 2007; Amin et al., 2008), primarily because it requires only length-frequency data (Gayanilo and Pauly, 1997). The objective of the present study was therefore to estimate the age and various population parameters such as asymptotic length, growth co-efficient, mortality, recruitment and level of exploitation, in order to assess the stock status of the species from Johor Straits, Malaysia. This study hopefully provides better understanding of the species and contributes to the establishment of the local conch fishery management.
MATERIALS AND METHODS
This study was conducted at Merambong Shoal, Johor Straits, Malaysia (01°19.778’ N, 103°35.798’ E) (Fig. 1), which is the traditional fishing ground for S. canarium and one of the most extensive seagrass covered subtidal shoal in Malaysian waters (Japar et al., 2006). Sample collection was conducted monthly, from January to December 2005, using belt-transect method (30×4 m).
Since, the species showed sexual dimorphism (Cob et al., 2008a, b), male and female S. canarium were analyzed separately. Shell-length was determined to the nearest 0.01 mm using a vernier caliper and animal weight (wet weight and ash free dry weight, AFDW) to the nearest 0.001 g using an analytical balance. A total of 2088 conch (766 males and 1322 females) were measured throughout the study period. The length-frequency data (Table 1) were analyzed using the FiSAT software (Gayanilo and Pauly, 1997). In order to identify the modes (cohorts) in the monthly length-frequency distributions, the Bhattacharya’s method (Bhattacharya, 1967), which was incorporated in the FiSAT package, was applied (Gayanilo and Pauly, 1997). The relationship between the total length (L) and weight (W) was computed using the formula of W = aLb (Quinn and Deriso, 1989), where a is the intercept (condition factor) and b is the slope (relative growth rate). The parameter a and b were estimated by least squares linear regression on log-log transformed data: log10 W = log10 a+b log10 L (Scherrer, 1984). The coefficient of determination (R2) was used as an indicator of the quality of the linear regression. Additionally, 95% confidence limits of the parameters a and b and the statistical significance level of the R2 were estimated.
The asymptotic length (L∞) and growth coefficient (K) of the Von Bertalanffy Growth Function (VBGF) were estimated by means of ELEFAN-I (Pauly and David, 1981). The VBGF is defined by the equation of:
where, Lt is mean length (mm) at age t, L∞ is asymptotic length (mm), K is growth coefficient (year-1), t is age (year) and to is the hypothetical age at which the length is zero (year) (Newman, 2002).
The estimated L∞ and K was used to calculate the growth performance index (φ’) using the equation of φ’ = 2 log10 L∞ + log10 K (Pauly and Munro, 1984).
 | |
| Fig. 1: | The study area. Monthly sample collections were conducted at the Merambong Shoal, Johor Straits, Malaysia (arrow) |
| Table 1: | Monthly length-frequency distribution of male (M) and female (F) Strombus canarium |
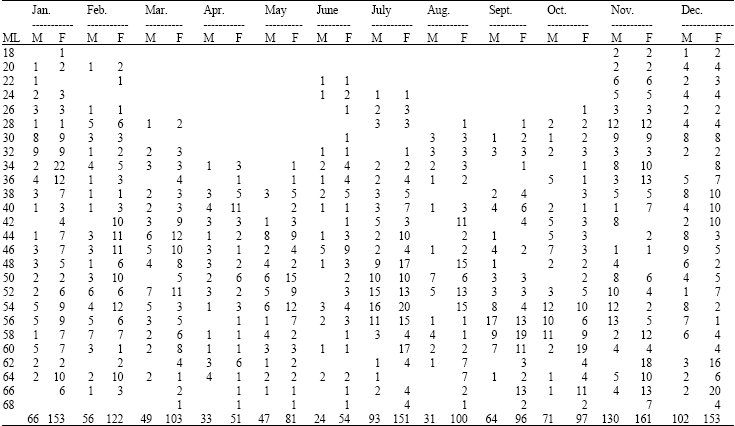 | |
| ML: Mid length class (mm) | |
The recruitment pattern was determined by backward projection on the length axis of the set of available length-frequency data, which determine the number of pulses per year and the relative strength of each pulse (Gayanilo and Pauly, 1997).
Once, the growth parameters of the VBGF were obtained, a linearized length-converted catch curve was constructed to estimate total mortality (Z) using the equation:
ln(Nt/Δt) = a+bt |
where, N is the number of individuals of relative age (t) and Δt is the time needed for the conch to grow through a length class. The slope (b) of the curve with its sign changed gives Z (Pauly, 1983).
Natural mortality rate (M) on the other hand, was estimated based on Beverton’s equation:
Lopt = L8×[3/(3+M/K)] |
where, Lopt is the optimum shell-length of the population i.e., length class with maximum sum of weight (Beverton, 1992).
This equation was derived from Beverton’s (1992) assumption that there is an intermediate age (topt) at which the biomass of a year class reaches a maximum. Solving this equation for M resulted in M = K×[(3 L∞/Lopt)–3]. To obtain an estimate of Lopt, the animal weight in each length classes (class size = 2 mm) were summed up and the length class with maximum weight was determined, as was described in Froese and Palomares (2000). The widely used Pauly’s empirical formula (Pauly, 1980), which was derived from data of 175 different fish stocks, was not applied to the species studied as this equation does not covered mollusks or any other invertebrates, other than the fish (Sparre and Venema, 1998).
Once the Z and M were obtained, fishing mortality (F) was estimated using the relationship of F = Z–M. The exploitation level (E) was obtained by the relationship of E = F/Z (Gulland, 1971). Relative Yield per Recruit (Y/R) and Biomass per Recruit (B/R) were estimated according to the method of Beverton and Holt (1966), using the knife-edge selection model. From the analysis Emax (maximum exploitation rate giving maximum relative yield-per-recruit) was estimated.
RESULTS
Population structure and length-weight relationship: There were wide variations in monthly length-frequency distribution and abundance for both male (Fig. 2) and female (Fig. 3) S. canarium. Females were found more abundant than males, with an average monthly density of 0.23±0.05 and 0.14±0.03 conch m-2, respectively.
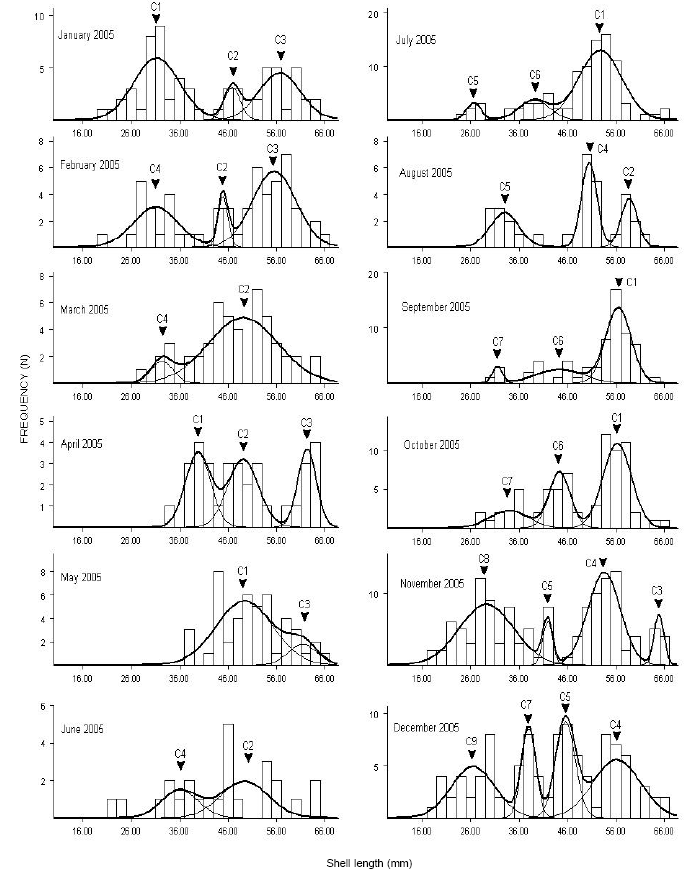 | |
| Fig. 2: | Monthly length frequency distributions of male Strombus canarium from Merambong Shoal, Johor Straits, Malaysia |
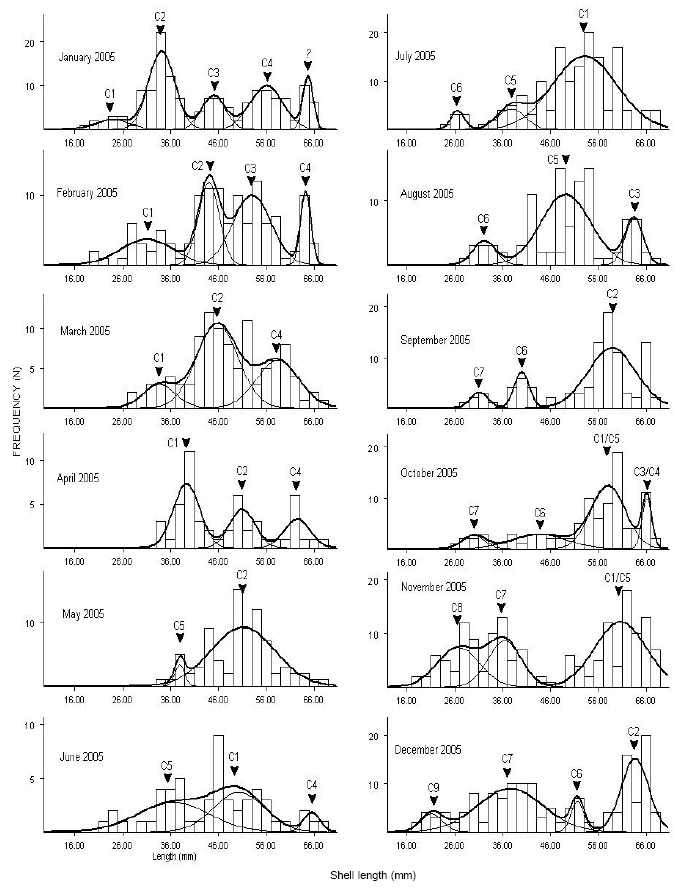 | |
| Fig. 3: | Monthly length frequency distributions of female Strombus canarium from Merambong Shoal, Johor Straits, Malaysia |
The mean monthly sex ratio (male/female) was 1/1.88±0.16 (range = 1/1.23 to 1/3.23, N = 12). The length-weight relationship is shown in Fig. 4, which can be summarized as Log W = –5.74 + 3.24 Log SL for male (R2 = 93.9; p<0.05) and Log W = –5.83 + 3.30 Log SL for female (R2= 93.2; p<0.05).
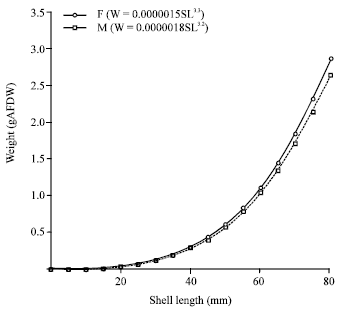 | |
| Fig. 4: | Length-weight relationship of Strombus canarium from Merambong Shoal, Johor Straits, Malaysia. M: Male, F: Female |
Ninety five percent confidence limit for parameter b ranged from 3.10 to 3.38 for male and from 3.15 to 3.45 for female. The covariance between shell length and weight (AFDW) was positive for both sexes (female = 4.14, male = 3.25), which suggests that as shell length increases, weight also increases. Further analysis using Peasrson’s correlation showed that shell length and shell weight was significantly correlated to each other (female = 0.81, p<0.05; male = 0.85, p<0.05).
Growth parameters: The asymptotic length (L∞) of the VBGF was estimated at 69.30 mm for males and 70.20 mm for females. The best estimated value of K was 1.20 year-1 for male (Rn = 0.146) and 1.50 year-1 for female (Rn = 0.170). Figure 5 shows the computed VBGF growth curve using these parameters, which indicating major spawning around February-March. The recruitment pattern was continuous throughout the year with one pulse of peak recruitment period between July-August for male and between June-July for female (Fig. 6). The growth performance index (φ’) was 3.81 for female and 3.48 for male. The predicted maximum length was 67.47 mm (95% CI = 66.29-68.65 mm) for male and 69.20 mm (95% CI = for 68.51-69.89 mm) female.
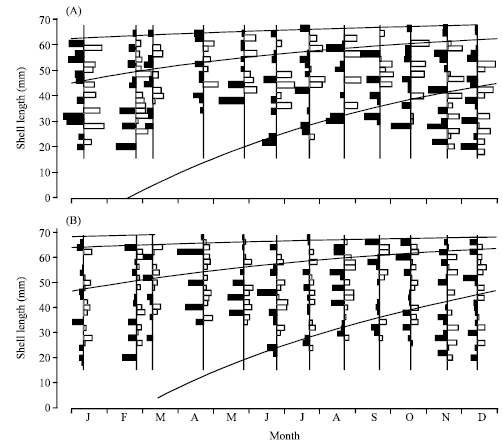 | |
| Fig. 5: | Restructured length-frequency distribution with growth curves superimposed using ELEFAN-1 for Strombus canarium from Merambong Shoal, Malaysia: (A) males (L∞ = 69.30 mm; K = 1.20 year-1) and (B) females (L∞ = 70.20 mm; K = 1.50 year-1) |
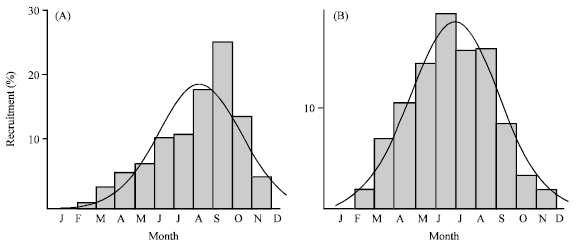 | |
| Fig. 6: | Recruitment pattern of Strombus canarium at Merambong Shoal, Johor straits, Malaysia, (A) male and (b) female |
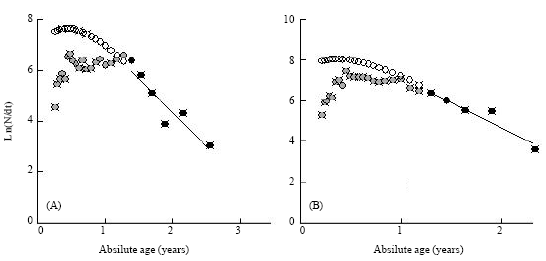 | |
| Fig. 7: | Length converted catch curve for male (A) and female (B) Strombus canarium from Merambong Shoal, Malaysia. Solid circles: Points used in calculating through least square linear regression and Open circles: Point either not fully recruited or nearing L∞ |
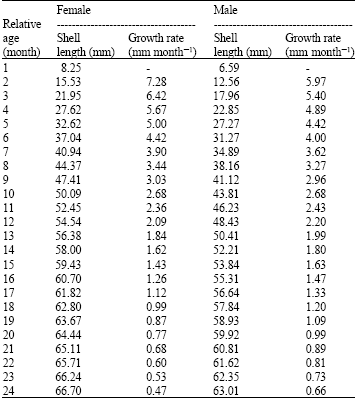
Age and growth: Based on growth coefficients, the maximum life span (Tmax = 3/K) of males and females was 2.5 and 2.0 year, respectively. By using the Bhattacharya’s method, there were at least 2 to 4 cohorts identified from the monthly length frequency distribution of the males and 2 to 5 cohorts of the females. Table 2 presented the length-at-age relationship estimated using the Bertalanffy’s model, by assuming to = 0 (Pauly and David, 1981). The sizes attained by males and females were 48.45 and 54.54 mm at the end of one year age, respectively.
Mortality and exploitation: The summed weight at specific length-class analysis suggested an optimum length (Lopt) of 56 mm in males and 58 mm in females. Solving the equation of M = Kx[(3 L∞/Lopt)-3], the natural mortality rate (M) was 0.86 year-1 for males and 0.95 year-1 for females.
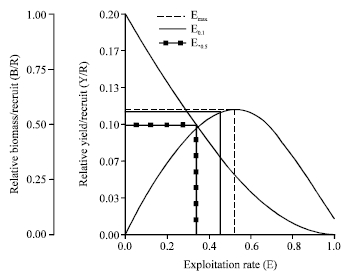 | |
| Fig. 8: | Relative yield per recruit (Y/R) and relative biomass per recruit (B/R) for male Strombus canarium at Merambong Shoal, Johor Straits, Malaysia |
| Table 2: | Length-at-age key for Strombus canarium from Merambong Shoal, Johor Straits, Malaysia estimated using the VBGF parameters (L∞ = 70.20 mm, K = 1.50 year-1 for females; L∞ = 69.30 mm, K = 1.20 year-1 for males) |
 | |
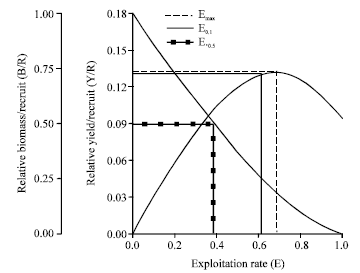 | |
| Fig. 9: | Relative yield per recruit (Y/R) and relative biomass per recruit (B/R) for female Strombus canarium at Merambong Shoal, Johor Straits, Malaysia |
Using the length converted catch curve analysis (Fig. 7) the total mortality rate (Z) was 2.72 year-1 (95% C.I. = 1.29- 4.14 year-1) for males and 2.56 year-1 (95% C.I. = 1.22-3.90 year-1) for females. From the equation of F = Z–M, the fishing mortality rates was 1.86 year-1 for males and 1.61 year-1 for females. The current exploitation rates (E = F/Z) were 0.68 for males and 0.63 for females, which were higher than the optimum level of exploitation (E = 0.50).
Relative yield per recruit and biomass per recruit: Figure 8 and 9 show the results of the relative Y/R and B/R analysis for males and females S. canarium using two types of selection curves. The computed maximum allowable limit of exploitation (Emax) for the relative Y/R and B/R were 0.521 for males and 0.528 for females. These maximum allowable limits of exploitation correspond to fishing mortality values of 1.42 year-1 for males and 1.35 year-1 for females.
DISCUSSION
The findings reported in this study represent the first data on population dynamics of S. canarium, based on separate male and female sub-populations. The estimated growth parameter showed higher asymptotic length and growth coefficient in females (L∞ = 70.20 mm, K = 1.50 year-1) compared with the males (L∞ = 69.30 mm, K = 1.20 year-1). This resulted in better overall growth performance where the phi index (φ’) was higher in females (φ’ = 3.81) compared with the males (φ’ = 3.48). The result was in agreements with reported values of other Strombus species where the females generally bigger than the males (Reed, 1993; Barroso et al., 2005). The asymptotic length values (males and females) was however lower when compared with the neighboring population at Bintan Island, Indonesia (Amini and Pralampita, 1987), which can grow up to 82.50 mm shell length. This condition might be due to overfishing. The conch were harvested by manual picking, thus larger individuals have more chances to be collected, resulting in lower numbers of larger individuals within the population. Nevertheless, the growth constant (K) was quite similar and even comparable with some other gastropod species (Siddeek and Johnson, 1997; Malaquias and Sprung, 2005). The growth performance index (φ’) ofmale and female conch recorded in this study was well within the values recorded for marine gastropods from all latitudes that ranged between 2.2 to 4.7 (Wolff, 1994).
High value of natural mortality (M) was recorded in this study. Natural mortality is the mortality created by all other factors than fishing. Among factors that might contribute to the high mortality of the species studied includes predation, diseases, spawning stress and old age. Conch predators such as the volute snails (Cymbiola nobilis and Melo melo), rays, dugong etc., that present within the study area might have contributed to the high M-value, which however need further studies. Other factors that linked to the ambient ecosystem might also contribute to the high M, e.g., water quality parameters and pollution. Earlier study showed high levels of heavy metals and organotins in the sediment and water column of the study area (Tan, 1999; Sudaryanto et al., 2004), which might impacted the reproduction, spawning and recruitment of the species. Mollusks are very sensitive to organotins and have widely been used in marine pollutants monitoring exercises (Chinishikawa et al., 2004). More studies are therefore urgently needed in this context.
The annual recruitment pattern of male and female S. canarium consists of one pulse, which peaked between the months of July-August and June-July, respectively. These pulses however were not an indication of the major settlement period, but rather representing the period where group of juveniles (sub-adult stage) migrated into the adult colony (Stoner, 2003). The actual major settlement period should be around February and March as indicated by the back projection of the VBGF growth curves (Fig. 5). In this study, the early juvenile groups of less than 18 mm shell length was never sampled, which strongly suggesting the presence of specific nursery habitat (settlement sites) for the species. The location and characteristic of this habitat was however not investigated in the current study and was therefore subjected for further investigation. The establishment of conch nursery habitat have been reported in many studies where mass migration of the juveniles to the adult population occurred after they attained certain age/length size (Stoner et al., 1994, 1996; Stoner, 2003).
Gastropods fishery play an important role in the national economy of many countries (Stoner and Ray, 1996; Leiva and Castilla, 2001; Keegan et al., 2003). Due to their high economic values and excessive harvest many marine gastropods show serious problems of overexploitation (Hobday et al., 2001; Leiva and Castilla, 2001; Schmidt et al., 2002). The higher fishing mortality rates as oppose to the natural mortality rates observed in this study indicates an unequal position of the stock, which could lead to overexploitation (Amin et al., 2008). Indeed, the exploitation rates established for the species (E = 0.68 for males and E = 0.63 for females) was higher than the optimum level of exploitation suggested by Gulland (1971). The current exploitation rates were even higher than the projected maximum allowable limit of exploitation (Emax= 0.521 for males; 0.528 for females), which is a further indication of overexploitation. In addition the relatively low asymptotic length values estimated from this study also suggesting that the population was overfished.
As a conclusion, this study has successfully estimated the age, asymptotic length, growth rate, mortality rate and recruitment for both male and female S. canarium, which has never been reported before. In addition the current position and level of exploitation of the stock was also established. For sustainable utilization of the resource, the exploitation rate of S. canarium should therefore be reduced. This can be achieved by lowering the fishing mortality rates, or by introducing minimum catch size regulation that could maintain sufficient stock of reproductively active individuals.
ACKNOWLEDGMENTS
The authors would like to thank the deanery and staffs of the Biology Department, Faculty of Science, University Putra Malaysia and School of Environmental and Natural Resource Sciences, University Kebangsaan Malaysia for technical supports, equipments and laboratory facilities provided. First author would also like to thanks University Kebangsaan Malaysia for the research grant (ST-020-2005 and UKM-ST-08-FRGS0001-2008) which makes this study possible.
REFERENCES
- Al-Barwani, S.M., A. Arshad, S.M.N. Amin, S.B. Japar, S.S. Siraj and C.K. Yap, 2007. Population dynamics of the green mussel Perna viridis from the high spat-fall coastal water of Malacca, Peninsular Malaysia. Fish. Res., 84: 147-152.
Direct Link - Amin, S.M.N., M. Zafar and A. Halim, 2008. Age, growth, mortality and population structure of the oyster, Crassostrea madrasensis, in the Moheskhali Channel (Southeastern coast of Bangladesh). J. Applied Ichthyol., 24: 18-25.
CrossRefDirect Link - Beverton, R.J.H., 1992. Patterns of reproductive strategy parameters in some marine teleost fishes. J. Fish. Biol., 41: 137-160.
Direct Link - Bhattacharya, C.G., 1967. A simple method of resolution of a distribution into Gaussian components. Biometrics, 23: 115-135.
PubMed - Blaber, S.J.M., J. Staunton-Smith, D.A. Milton, G. Fry and T.V. Velde et al., 1998. The biology and life history strategies of Ilisha (Teleostei: Pristigasteridae) in the coastal waters and estuaries of Sarawak. Estuar. Coast. Shelf Sci., 47: 499-511.
Direct Link - Cob, Z.C., A. Arshad, J.S. Bujang and M.A. Ghaffar, 2008. Sexual maturity and sex determination in Strombus canarium Linnaeus, 1758 (Gastropoda: Strombidae). J. Biol. Sci., 8: 616-621.
CrossRefDirect Link - Cob, Z.C., A. Arshad, J.S. Bujang and M.A. Ghaffar, 2009. Species description and distribution of Strombus (Mollusca, Strombidae) in Johor straits and its surrounding areas. Sains Malaysiana, 38: 39-46.
Direct Link - Froese, R. and M.L.D. Palomares, 2000. Growth, natural mortality, length-weight relationship, maximum length and length-at-first-maturity of the coelacanth Latimeria chalumnae. Environ. Biol. Fish., 58: 45-52.
CrossRefDirect Link - Hobday, A.J., M.J. Tegner and P.L. Haaker, 2001. Overexploitation of a broadcast spawning marine invertebrates: Decline of the white abalone. Rev. Fish Biol. Fish., 10: 493-514.
Direct Link - Bujang, J.S., M.H. Zakaria and A. Arshad, 2006. Distribution and significance of seagrass ecosystems in Malaysia. Aquat. Ecosyst. Health Manage., 9: 203-214.
CrossRefDirect Link - Leiva, G.E. and J.C. Castilla, 2001. A review of the world marine gastropod fishery: Evolution of catches, management and the Chilean experience. Rev. Fish Biol. Fish., 11: 283-300.
CrossRefDirect Link - Keegan, W.F., R.W. Portell and J. Slapcinsky, 2003. Changes in invertebrate taxa at two pre-Columbian sites in Southern Jamaica, AD 800-1500. J. Arch. Sci., 30: 1607-1617.
Direct Link - Malaquias, M.A.E. and M.J. Sprung, 2005. Population biology of the cephalaspidean mollusc Haminoea orbygniana in a temperate coastal lagoon (Ria Formosa, Portugal). Estuarine Coastal Shelf Sci., 63: 177-185.
CrossRefDirect Link - Mancera, E. and J. Mendo, 1996. Population dynamics of the oyster Crassostrea rhizophorae from the Cienaga Grande de Santa Marta, Colombia. Fish. Res., 26: 139-148.
CrossRefDirect Link - Newman, S.J., 2002. Growth, age estimation and preliminary estimates of longevity and mortality in the moses perch, Lutjanus russelli (Indian ocean form), from continental shelf waters off North-western Australia. Asian fish. Sci., 15: 283-294.
Direct Link - Pauly, D., 1980. On the interrelationships between natural mortality, growth parameters and mean environmental temperature in 175 fish stocks. J. Cons. Int. Explor. Mer, 39: 175-192.
CrossRefDirect Link - Pauly, D. and N. David, 1981. ELEFAN-I, a basic program for the objective extraction of growth parameters from Length frequency data. Meeresforschung, 28: 205-211.
Direct Link - Pauly, D. and J.L. Munro, 1984. Once more on the comparison of growth in fish and invertebrates. Fishbyte, 2: 21-21.
Direct Link - Purchon, R.D and D.E.A. Purchon, 1981. The Marine Shelled Mollusca of West Malaysia and Singapore. Part 1. General Introduction and an account of the collecting Stations J. Moll. Stud., 47: 290-312.
Direct Link - Schmidt, S., M. Wolff and J.A. Vargas, 2002. Population ecology and fishery of Cittarium pica (Gastropoda: Trochidae) on the Caribbean coast of Costa Rica. Rev. Biol. Trop., 50: 1079-1090.
Direct Link - Siddeek, M.S. and D.W. Johnson, 1997. Growth parameter estimates for Omani abalone (Haliotis mariae, Wood 1828) using length-frequency data. Fisheries Res., 31: 169-188.
CrossRefDirect Link - Stoner, A.W., 2003. What constitutes essential nursery habitat for a marine species? A case study of habitat form and function for queen conch. Mar. Ecol. Prog. Ser., 257: 275-289.
Direct Link - Stoner, A.W. and M. Ray, 1996. Shell remains provide clues to historical distribution and abundance patterns in a large seagrass-associated gastropod (Strombus gigas). Mar. Ecol. Prog. Ser., 135: 101-108.
Direct Link - Stoner, A.W., M.D. Hanisak, N.P. Smith and R.A. Armstrong, 1994. Large-scale Distribution of Queen Conch Nursery Habitats: Implications for Stock Enhancement. In: Queen Conch Biology, Fisheries and Mariculture, Appeldoorn, R.S. and B. Rodr�guez (Eds.). Fundaci�n Cient�fica Los Roques, Caracas, Venezuela, pp: 169-189.
- Stoner, A.W., P.A. Pitts and R.A. Armstrong, 1996. Interaction of physical and biological factors in the large-scale distribution of juvenile queen conch in seagrass meadows. Bull. Mar. Sci. 58: 217-233.
Direct Link - Barroso, C.M., M.H. Moreira and C.A. Richardson, 2005. Age and growth of Nassarius reticulatus in the Ria de Aveiro, North-West Portugal. J. Mar. Biol. Assoc., 85: 151-156.
CrossRef - Chinishikawa, J.J., S. Mamiya, T. Kanayama, T. Nishikawa, F. Shiraishi and T. Horiguchi, 2004. Involvement of the retinoid X receptor in the development of imposex caused by organotins in gastropods. Environ. Sci. Technol., 38: 6271-6276.
CrossRef - Cob, Z.C., A. Arshad, H.M. Idris, B. Japar Sidik and A.G. Mazlan, 2008. Sexual polymorphisms in a population of Strombus canarium Linnaeus, 1758 (Mollusca: Gastropoda) at Merambong shoal, Malaysia. Zool. Stud., 47: 318-325.
Direct Link - Sudaryanto, A., S. Takahashi, H. Iwata, S. Tanabe and A. Ismail, 2004. Contamination of butyltin compounds in Malaysian marine environments. Environ. Pollut., 130: 347-358.
CrossRefPubMedDirect Link - Tan, K.S., 1999. Imposex in thais gradata and Chicoreus capucinus (Mollusca, Neogastropoda, Muricidae) from the Straits of Johor: A case study using penis length, area and weight as measures of imposex severity. Marine Pollut. Bull., 39: 295-303.
CrossRefDirect Link