
M.E. Abu Ziada
Department of Botany, Faculty of Science, Mansoura University, Egypt
I.A. Mashaly
Department of Botany, Faculty of Science, Mansoura University, Egypt
M. Abd El-Monem
Department of Botany, Faculty of Science, Mansoura University, Egypt
M. Torky
Department of Botany, Faculty of Science, Mansoura University, Egypt
Journal of Applied Sciences
Year: 2008 | Volume: 8 | Issue: 8 | Page No.: 1395-1405







ABSTRACT
The present study provides quantitative assessment of the vegetative yield, growth characteristics, metabolic products, elemental composition and antimicrobial bioactivity of five common macrohydrophytes: Bolboschoenus glaucus (Cyperaceae), Veronica anagallis-aquatica (Scrophulariaceae), Nymphaea lotus (Nymphaceae), Pistia stratiotes (Araceae) and Myriophyllum spicatum (Haloragidaceae). These plants tend to flourish vegetatively during the summer season (June-August). Their relative growth rate, relative assimilating surface growth rate and net assimilation rate were higher during early vegetative stage (February-May). The highest percentages of protein and lipids content were recorded in Nymphaea, while the crude fiber content was higher in Bolboschoenus than in other species. The macronutrient elements were detected with relatively high concentration and sodium cation appeared to be an essential accumulatent as compared with K, Ca and Mg. Myriophyllum appeared to be the major accumulator species of heavy metals, while Pistia appeared to be the minor one. Sterols, alkaloids, flavonoids, tannins, saponins and resins were detected in these plants. Nymphyaea was found to have the most effective antimicrobial activities than the other studied species.
PDF Abstract XML References Citation
How to cite this article
M.E. Abu Ziada, I.A. Mashaly, M. Abd El-Monem and M. Torky, 2008. Economic Potentialities of Some Aquatic Plants Growing in North East Nile Delta, Egypt. Journal of Applied Sciences, 8: 1395-1405.
DOI: 10.3923/jas.2008.1395.1405
URL: https://scialert.net/abstract/?doi=jas.2008.1395.1405
DOI: 10.3923/jas.2008.1395.1405
URL: https://scialert.net/abstract/?doi=jas.2008.1395.1405
INTRODUCTION
Five macrohydrophytes representing the different forms of aquatic vegetation in the Nile Delta region have been selected for the present investigation. Bolboschoenus glaucus and Veronica anagallis-aquatica are belonging to emergent hydrophytes, Nymphaea lotus is belonging to rooted floating hydrophytes, Pistia stratiotes is belonging to free floating hydrophytes and Myriophyllum spicatum is belonging to rooted submerged hydrophytes.
B. glaucus is a small grass-like perennial sedge of saline to fresh watershores (Browning, 1998). It was able to bioaccumulate and phytostabilization of Cd and Pb in its roots (Almedia et al., 2006).
V. anagallis-aquatica is a perennial herb, often 4-angled towards the base, commonly spread in marshy ground, river-banks and irrigation channels (Boulos, 2002). Pandey and Sirvastava (1989), Harput et al. (2004) and Kupeli et al. (2005) mentioned that, Veronica, a semi-aquatic weed, is a potential source of leaf protein and iridoid glycosides.
N. lotus is herbaceous aquatic plant, whose leaves floats or submerged in water. It is a good phytoaccumulator and can selectively bioaccumulate heavy metals particularly zinc and lead (Khedr and Hegazy, 1998).
P. stratiotes is a free-floating stoloniferous herb commonly found in ponds and streams. Its leaves are obovate, light green in colour and have many prominent longitudinal veins (Arber, 1991). The oil extracted from Pistia is used in the treatment of worm infestations, tuberculosis and dysentery and is applied externally to treat skin diseases, inflammation, piles, ulcers and burns (Kirtikar and Basu, 2000). Pistia leaves possess antifungal properties that explain the use of this plant in folk medicine for the treatment of various diseases whose symptoms might involve fungal infections (Premkumar and Shyamsundar, 2005).
M. spicatum is a fresh water angiosperm that contains high concentrations of tannins and secondary metabolites known for their antimicrobial properties (Walenciak et al., 2002). Elisabeth et al. (1996) stated that aqueous acetone extract of the shoot of M. spicatum exhibit an inhibitory action against various coccoid and filamentous Cyanobacteria.
The present study aims at evaluation of the periodical changes in the vegetative yield, growth characteristics, chemical constituents and antimicrobial bioactivities of the investigated plants.
MATERIALS AND METHODS
The plant samples were collected monthly for one year from their natural habitats using quadrates (50x50 cm).Samples were taken along two parallel transects located in the central portion of the representative stand and new quadrate location were selected so that production was not influenced by previous sampling (Clark and Clay, 1984). Ten individuals of each species were randomly chosen and used for measurement of the growth parameters; mean height of stems or length of stolons, number of leaves and their areas. Plants of each quadrate were air-dried and the biomass of the different plant parts were measured separately and expressed as g dry wt. m-2 (Cochran, 1963; Polisetty et al., 1984). Data of the successive estimation of the assimilating surface area and the biomass were applied to estimate growth characteristics as described by Radford (1967), Chapman (1976) and Coombs and Hall (1982). The growth characteristics measured are: Relative Growth Rate (RGR), Relative Assimilating Surface Growth Rate (RASGR), Net Assimilation Rate (NAR) and Leaf Area Ratio (LAR). For phytochemical analysis, plant samples were collected, handly cleaned, air dried and ground to fine powder. In each sample, moisture content, total ash, water soluble ash, acid insoluble ash, total nitrogen, total protein, total lipid, crude fiber, total soluble sugars, glucose, sucrose and polysaccharides were determined according to the methods adopted by Ward and Johnson (1962), Handel (1968) and Thayumanavan and Sadasivam (1984). Phytochemical screening was carried out using the powdered samples and the alcoholic extracts to detect the active principles: Glycosides, sterols, alkaloids, flavonoides, tannins, saponins and resins according to Claus (1967), Harper (1975) and Markham (1982). For extraction of the different elements, 0.1 g of air-dried powder was digested by concentrated HNO3, heated gently until the solution turned quite clear. The samples were made up to a known volume by distilled water. Na, K and Ca were determined by flame photometer, while Mg, Fe, Mn, Zn, Cu, Ni, Cd, Pb and As were estimated by atomic absorption spectrometer (Allen et al., 1974). The elements were expressed as mg/100 g dry weight. For antimicrobial screening, methanolic extracts were prepared using 100 g of each powdered samples and 400 mL of 80% methanol by refluxing for 3 h. A stock solution of extract was prepared in dimethyl sulfoxide (DSMO) and kept at -20°C for antimicrobial assay (Mehraban et al., 2005). The bacterial strains that used as tested organisms are Bacillus subtilis, Erwinia cartovora, Escherichia coli, Pseudomonas fluorescence and Staphylococcus aureus, while the tested fungi are Alternaria alternata, Aspergillus niger, Bibolaris oryza, Botrytis faba, Fusarium oxysporium and Penicillium chrysogenum. The extracts were screened for their inhibitory activities against the tested bacteria and fungi using agar diffusion technique (Calvo et al., 1986; Deans and Ritchie, 1987). After inoculation with constant inoculums, the plates were incubated for 24 h for the bacterial strains and 3-4 days for fungi. Controls had solvent (DSMO) without extracts of the tested plants. The antimicrobial bioactivity was determined by the measuring the diameters of inhibition zones in cm.
RESULTS AND DISCUSSION
Vegetative yield: Records of the monthly variations in the assimilating surface area (cm2 m-2) and biomass content (g dry wt. m-2) of B. glaucus, M. spicatus, N. lotus, P. stratiotes and V. anagallis-aquatica are demonstrated in Fig. 1-10. The maximum assimilating surface area of the five species (4277.3, 94580.0, 66652.9, 89347.7 and 11774.2 cm2 m-2, respectively) were attained in August except veronica in June. The phytomass showed a similar trend, increasing gradually from February till reached its peak in August (16.6, 39.8, 251.8, 80.2 and 57.9 g dry wt. m-2, respectively) and coincided with the maximum leaf, stem and root biomass. At maturity stage, which begins in September, it showed a gradual decline. The maximum necromass was in January.
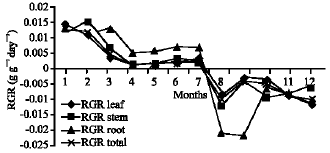
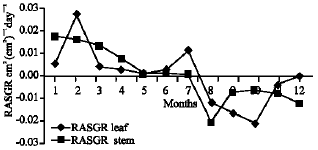
Growth characteristics: The monthly changes in the Relative Growth Rate (RGR) of the studied species are presented in Fig. 11-15. It is obvious that, the total RGR is generally higher at early vegetative stage than at maturity (fruiting stage). The maximum RGR of B. glaucus and M. spicatum (0.0081-0.0129 g g-1 day-1, respectively) were recorded in winter (January-February), those of N. lotus and P. stratiotes were 0.0408-0.0132 g g-1 day-1, respectively in spring (March-May) and that of V. anagallis-aquatica was 0.0515 g g-1day-1 during October. The results demonstrated in Fig. 16-20 indicated that, the highest Relative Assimilating Surface Growth Rate (RASGR) of leaves and stem of B. glaucus were 0.015 and 0.03 cm2 (cm2)-1 day-1, respectively then tend to decline and even became negative sign. The RASGR of M. spicatum leaves (0.001-0.027 cm2 (cm2)-1 day-1) and stem (0.001-0.017 cm2 (cm2)-1 day-1) were increased at the vegetative stage and beginning of flowering. At fruiting, RASGR as well as RGR suddenly became a negative sign. The monthly changes of RASGR of leaf lamina of N. lotus fluctuated between 0.0396 and 0.0572 cm2 (cm2)-1 day-1. Those of P. strariotes ranged between 0.007 and 0.033 cm2 (cm2)-1 day-1 and V. anagallis-aquatica had (0.002-0.065 cm2 (cm2)-1 day-1).
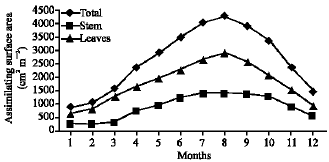 | |
| Fig. 1: | Monthly variation in the assimilating surface area of Bolboschoenus glaucus |
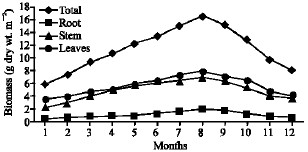 | |
| Fig. 2: | Monthly variation in the biomass of B. glaucus |
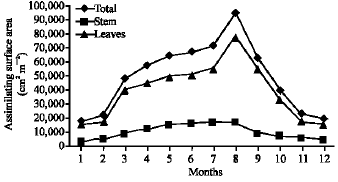 | |
| Fig. 3: | Monthly variation in the assimilating surface area of Myriophyllum spicatum |
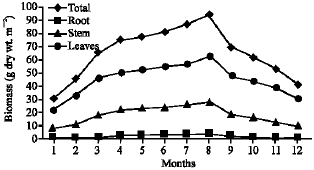 | |
| Fig. 4: | Monthly variation in the biomass of Myriophyllum spicatum |
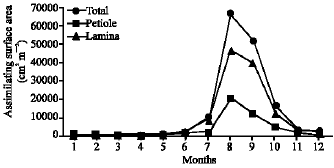 | |
| Fig. 5: | Monthly variation in the assimilating surface area of Nymphaea lotus |
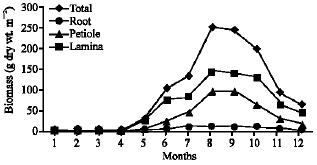 | |
| Fig. 6: | Monthly variation in the biomass of Nymphaea lotus |
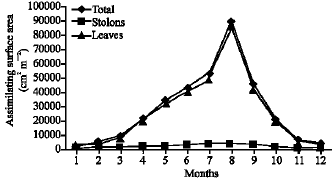 | |
| Fig. 7: | Monthly variation in the assimilating surface area of Pistia stratiotes |
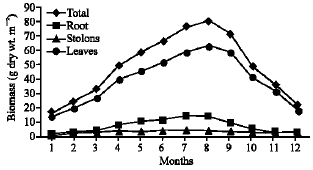 | |
| Fig. 8: | Monthly variation in the biomass of Pistia stratiotes |
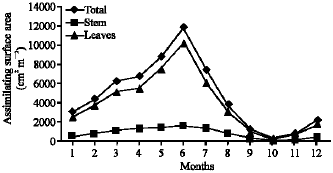 | |
| Fig. 9: | Monthly variation in the assimilating surface area of Veronica anagallis-aquatica |
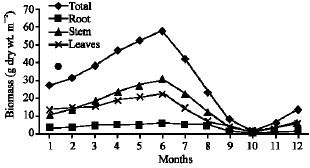 | |
| Fig. 10: | Monthly variation in the biomass of Veronica anagallis-aquatica |
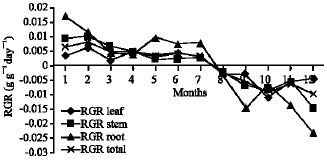 | |
| Fig. 11: | Monthly variation in RGR of Bolboschoenus glaucus |
 | |
| Fig. 12: | Monthly variation in RGR of Myriophyllum spicatum |
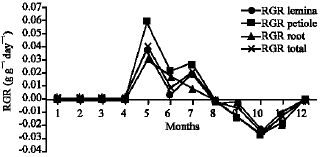 | |
| Fig. 13: | Monthly variation in RGR of Nymphaea lotus |
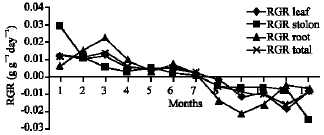 | |
| Fig. 14: | Monthly variation in RGR of Pistia stratiotes |
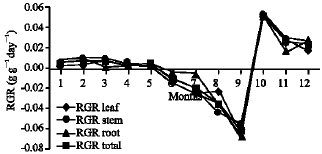 | |
| Fig. 15: | Monthly variation in RGR of RGR of Veronica anagallis-aquatica |
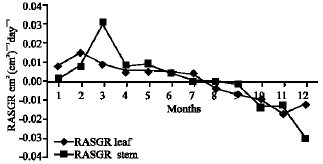 | |
| Fig. 16: | Monthly variation in RASGR of Bolboschoenus glaucus |
As illustrated in Fig. 21-25, the highest values of Net Assimilation Rate (NAR) were recorded in October for B. glaucus (0.00004 g (cm2)-1 day-1), in February for M. spicatum (0.0108 g (cm2)-1 day-1), in July for N. lotus (0.1332 g (cm2)-1 day-1) and in September for P. stratiotes (0.015 g (cm2)-1 day-1) and in December for V. anagallis-aquatica. The Leaf Area Ratio (LAR) of the studied plants showed gradual increase from July to October then decline during November and December (Fig. 26-30).
 | |
| Fig. 17: | Monthly variation in RASGR of Myriophyllum spicatum |
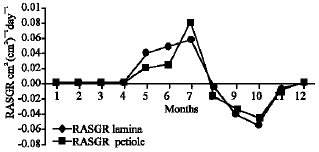 | |
| Fig. 18: | Monthly variation in RASGR of Nymphaea lotus |
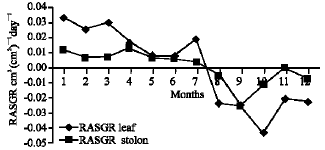 | |
| Fig. 19: | Monthly variation in RASGR of Pistia stratiotes |
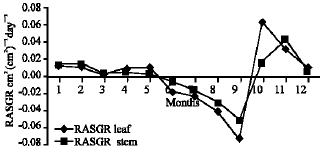 | |
| Fig. 20: | Monthly variation in RASGR of Veronica anagallis-aquatica |
From the above results it can be concluded that, the assimilating surface area and the biomass content increased gradually with advanced age then declined at the beginning of fruiting stage. Sometimes these two growth parameters elevated again due to appearance of new branches (El-Habibi et al., 1988).
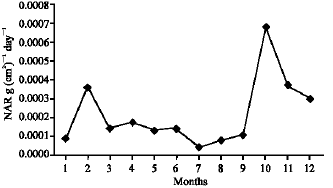 | |
| Fig. 21: | Monthly variation in NAR of Bolboschoenus glaucus |
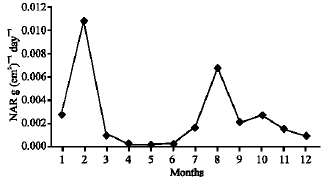 | |
| Fig. 22: | Monthly variation in NAR of Myriophyllum spicatum |
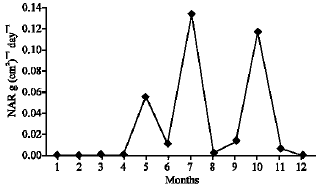 | |
| Fig. 23: | Monthly variation in NAR of Nymphaea lotus |
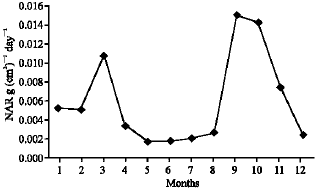 | |
| Fig. 24: | Monthly variation in NAR of Pistia stratiotes |
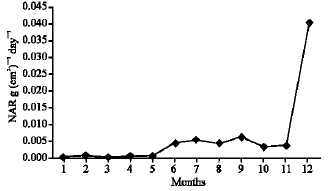 | |
| Fig. 25: | Monthly variation in NAR Veronica anagallis-aquatica |
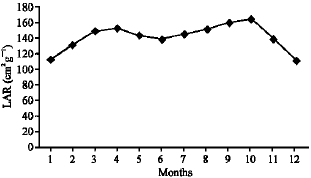 | |
| Fig. 26: | Monthly variation in LAR of Bolboschoenus glaucus |
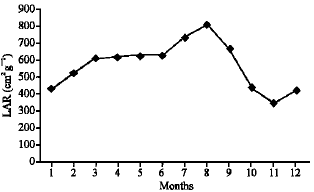 | |
| Fig. 27: | Monthly variation in LAR of Myriophyllum spicatum |
The relative assimilating surface growth rate showed the same trend of relative growth rate of these plants. The periodical fluctuation in the growth characteristics may be attributed to temperature changes. These findings are in accordance with those of Parsons (1980) and Papchenkov (1985). Abo El-Lil (1987) stated that, at the period of maturity, the dehydrated nutrient substances accumulated in the ripening seeds and with fruiting the lower leaves are about to fall. The reduction of weight may be related to these senescence phenomena in addition to competition.
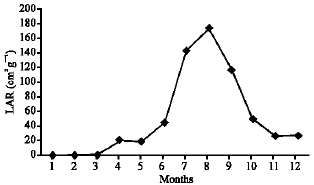 | |
| Fig. 28: | Monthly variation in LAR of Nymphaea lotus |
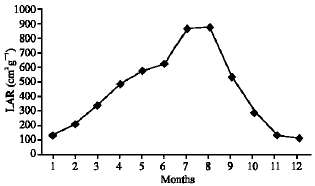 | |
| Fig. 29: | Monthly variation in LAR of Pistia stratiotes |
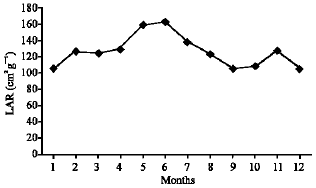 | |
| Fig. 30: | Monthly variation in LAR of Veronica anagallis-aquatica |
Phytochemical analyses
Chemical constituents: The obtained data in Table 1 indicated that, the highest values of moisture content (13.64%), crude fiber (40.0%), total soluble salts (339.3 mg g-1 dry wt.), glucose (11.6 mg g-1 dry wt.), sucrose (353.6 mg g-1 dry wt.), polysaccharides (456.4 mg g-1 dry wt.) and total carbohydrates contents (1160.9 mg g-1 dry wt.) recorded in B. glaucus while those of total nitrogen (230.0 mg g-1 dry wt.) total protein (120.5 mg g-1 dry wt.), total lipid (1.82 mg g-1 dry wt.) and water soluble ash (9.46%) contents were recorded in N. lotus. M. spicatum is attained the highest total ash (37.58%) and acid insoluble ash (17.8%) contents.
| Table 1: | Mean values of different metabolic products of the studied plants |
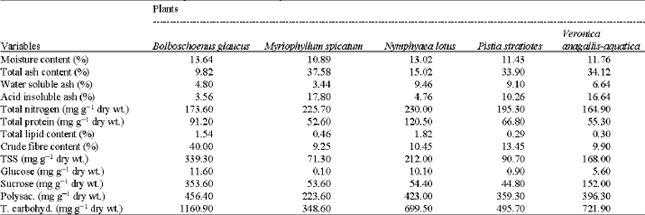 | |
| TSS = Total Soluble Sugars, Polysac. = Polysaccharides and T. carbohyd. = Total carbohydrates | |
| Table 2: | Concentrations of elements (expressed as mg/100 g dry wt.) in the investigated plants |
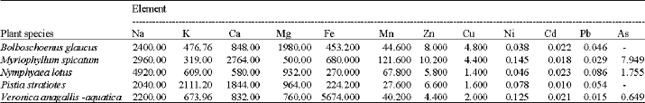 | |
| (-) sign = Undetectable value | |
| Table 3: | The inhibitory activity of the plant extracts against the tested bacteria as demonstrated by diameters of inhibition zones |
 | |
| Table 4: | The inhibitory activity of the plant extracts against the tested fungi as demonstrated by diameters of inhibition zones |
 | |
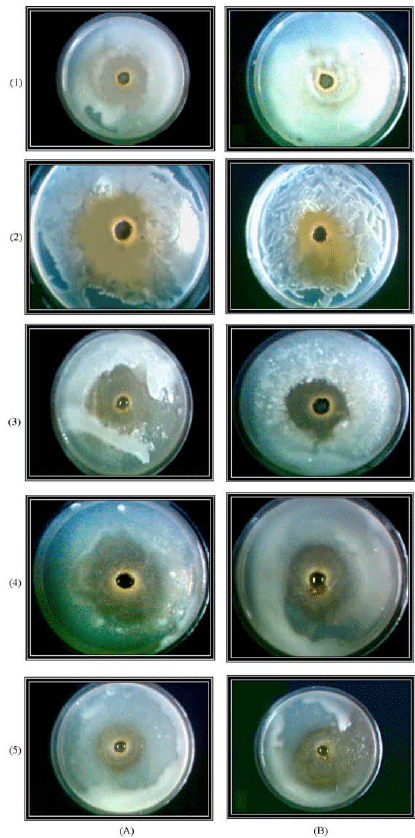 | |
| Plate 1: | The inhibitory activity of the methanolic extracts of the N. lotus (A) and V. anagallis-aquatica (B) against different bacterial strains. (1) Bacillus subtilis, (2) Erwinia carotovora carotovora, (3) Escherichia coli, (4) Pseudomonas fluorescence and (5) Staphylococcus aureus |
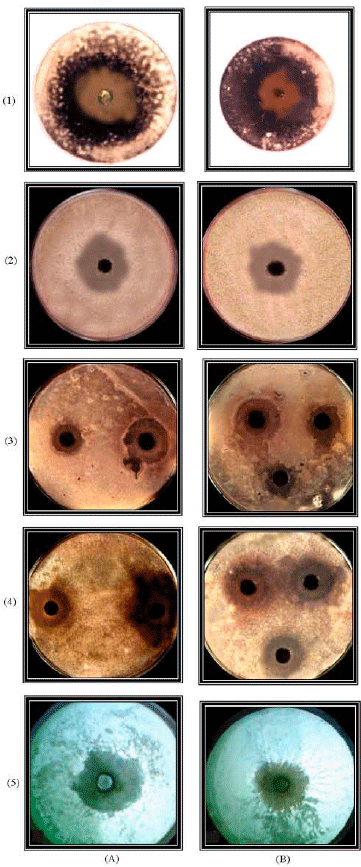 | |
| Plate 2: | The inhibitory activity of the methanolic extracts of the N. lotus (A) and V. anagallis-aquatica (B) against different fungal sspecies. (1) Alternaria alternata, (2) Aspergillus niger, (3) Bibolaris oryza, (4) Botrytis faba and (5) Penicillium chrysogenum |
In general, all the studied plants showed a relatively high concentration of carbohydrates.
The preliminary phytochemical screening revealed the presence of sterols, alkaloids, flavonoides, tannins, chlorides and sulphates in the studied plants. Resins were detected in B. spicatus and N. lotus, while saponins were absent in B. spicatus.
Elementary analysis: The highest value of sodium ion concentration was recorded in N. lotus (4920.0 mg/100 g dry wt.) and the lowest value was recorded in P. stratiotes (2040.0 mg/100 g dry wt.). Potassium ion concentration ranged between 319.0 mg/100 g dry wt. in M. spicatum and 2111.2 mg/100 g dry wt. in P. stratiotes. M. spicatum attained the highest value of calcium ion content (2764.0 mg/100 g dry wt.) while N. lotus attained the lowest value (580.0 mg/100 g dry wt.).
It is clear that, mg++ content of B. glaucus (1980.0 mg/100 g dry wt.) is relatively higher than that of other investigated plants. Its minimum value was that of M. spicatum being 500.0 mg/100 g dry wt. The results in Table 2 showed the obvious ability of these plants to absorb and accumulate heavy metals from the interstitial water. V. anagallis-aquatica has the highest value of ferric ion content (5674.0 mg/100 g dry wt.). The maximum values of manganese, zinc, nickel and arsenic (121.6, 10.2, 0.145 and 7.95 mg/100 g dry wt., respectively) were recorded in M. spicatum. The highest concentrations of both cadmium and lead ions were 0.023 and 0.086 mg/100 g dry wt., respectively in N. lotus, B. spicatus accumulated the highest copper ion content (4.8 mg/100 g dry wt.). The minimum concentrations of Fe, Mn and Cd were 224.2, 27.6 and 0.01 mg/100 g dry wt., respectively in P. stratiotes, those of Zn and Cu were 5.8 and 1.4 mg/100 g dry wt., respectively in N. lotus, Pb and As were 0.015 and 0.649 mg/100 g dry wt., respectively in N. anagallis-aquatica and that of nickel was 0.038 mg/100 g dry wt. B. glaucus and P. stratiotes showed undetectable values of arsenic.
The macro-nutrients (Na, K, Ca and Mg) were detected with relatively high concentrations. Sodium appeared to be an essential accumulatent in the investigated plants as compared with K, Ca and Mg. These results are coinciding with those obtained by Polisetty et al. (1984).
With respect heavy metals accumulation, M. spicatum appeared to be the major accumulator among the studied plants, while P. stratiotes appeared to be the minor one. According to the toxicological evaluations of the contaminants and naturally occurring toxicants carried out by the joint FAO/WHO (Food and Agricultural Organization/World Health Organization) expert committee on food additives for human consumption, the maximum permissible concentrations of the studied heavy metals: Fe, Zn, Cu, Cd, Pb and As are 0.8, 0.3-1.0, 0.05-0.5, 0.007, 0.025 and 0.015 mg kg-1 body weight, respectively (WHO, 1993, 1997). Consequently, the concentrations of all estimated heavy metals are obviously higher than the permissible levels and appeared to be harmful for human and therefore, the studied plants are not recommended as a fodder for animals consumption. The obtained data indicated that, these plants could be used as bioindicator for water pollution. Also, they appeared to have high potentiality for significant metals accumulation.
Antimicrobial potentialities
Antibacterial assay: The methanol extracts exhibited inhibitory activities against the tested bacterial strains with different degrees as demonstrated by measuring the diameters of inhibition zones (Table 3). The extracts of N. lotus and V. anagallis-aquatica showed the highest activity against the tested bacteria (Plate 1), while the extracts of B. glaucus and M. spicatum showed the lowest activity. The extract of P. stratiotes exhibited moderate range of antibacterial activity.
Antifungal assay: The antifungal activity of methanol extracts of the five plants presented in Table 4 and Plate 2. The extracts of N. lotus and V. anagallis-aquatica also showed the highest inhibitory activity against the tested fungi. In contrast, the extract of M. spicatum showed the lowest antifungal activity.
It is apparent that, the methanolic extract of N. lotus is found to have the most effective antimicrobial activities and showed a wider inhibition zone than the extract of other plants. From the results of both the present and the previous studies (Walenciak et al., 2002; Premkumar and Shuamsundar, 2005), it may be concluded the therapeutic possibilities of these plants. In this respect, Kupeli et al. (2005) found that V. anagallis-aquatica contained iridoid glycosides with antinociceptive and anti-inflammatory activities.
REFERENCES
- Allen, S.E., 1974. Chemical Analysis of Ecological Materials. Blackwell Scientific Publications, London, UK., Pages: 565.
Direct Link - Almeida, C.M.R., A.P. Mucha and M.T.S.D. Vasconcelos, 2006. Comparison of the role of the sea club-rush Scirpus maritimus and the sea rush Juncus maritimus in terms of concentration, speciation and bioaccumulation of metals in the estuarine sediment. Environ. Pollut., 142: 151-159.
CrossRefDirect Link - Arber, A., 1920. The vegetative morphology of Pistia and the Lemnaceae. Proc. R. Soc. London Ser. B: Contain. Pap. Biol. Character, 91: 96-103.
Direct Link - Boulos, L., 2002. Flora of Egypt. Vol. 3, Al-Hadara Publication, Cairo, Egypt, ISBN: 977-5429-25-0.
CrossRefDirect Link - Calvo, M.A., F.J. Cabanes and L. Abarca, 1986. Antifungal activity of some mediterranean algae. Mycopathologia, 93: 61-63.
CrossRefDirect Link - Clark, W.R. and R.T. Clay, 1984. Standing crop of sagittaria in the upper mississippi river. Can. J. Bot., 63: 1453-1457.
CrossRefDirect Link - Van Handel, E., 1968. Direct microdetermination of sucrose. Anal. Biochem., 22: 280-283.
CrossRefDirect Link - Harput, U.S., M. Varel, N. Akito and S. Iclal, 2004. Acylated iridoid glucosides from Veronica anagallis-aquatica. Phytochemistry, 65: 2135-2139.
CrossRefDirect Link - Khedr, A.H.A. and A.K. Hegazy, 1998. Ecology of the rampant weed Nymphaea lotus L. Willdenow in natural and rice field habitats of the Nile delta, Egypt. Hydrobiologia, 386: 119-129.
CrossRef - Kupeli, E., U.S. Harput, M. Varel, E. yesilada and I. Saracoglu, 2005. Bioassay-guided isolation of iridoid glucosides with antinociceptive and anti-inflammatory activities from Veronica anagallis-aquatica L. Ethnopharm. J., 102: 170-176.
CrossRefDirect Link - Pandey, V.N. and A.K. Srivastava, 1989. Veronica anagallis-aquatica L., A potential source of leaf protein. Aquat. Bot., 34: 358-388.
CrossRefDirect Link - Premkumar, V.G. and D. Shyamsundar, 2005. Antidermatophytic activity of Pistia stratiotes. Indian J. Pharm., 37: 127-128.
Direct Link - Radford, D.J., 1967. Growth analysis formulae-their use and abuse. Crop Sci., 7: 171-175.
CrossRefDirect Link - Walenciak, O., W. Zwisler and E.M. Gross, 2002. Influence of myriophyllum spicatum derived tannins on gut microbiota of its herbivore acentria ephemerella. J. Chem. Ecol., 28: 2045-2056.
Direct Link - Mehraban, F., O.T. Nasim and J. Fereshteh, 2005. Antidermatophyte activities of Eucalyptus camaldulensis in comparison with griseofulvin Iran. J. Pharmacol. Therapeut., 4: 80-83.
Direct Link - Deans, S.G. and G. Ritchie, 1987. Antibacterial properties of plant essential oils. Int. J. Food Microbiol., 5: 165-180.
CrossRefDirect Link