
A. Mebarkia
Department of Agronomy, Faculty of Sciences Nature and Life, University Setif 1, Algeria
Y. Abdi
Department of Agronomy, Faculty of Sciences Nature and Life, University Setif 1, Algeria
S. Rahmani
Department of Agronomy, Faculty of Sciences Nature and Life, University Setif 1, Algeria
A. Abjabi
Department of Agronomy, Faculty of Sciences Nature and Life, University Setif 1, Algeria
Journal of Agronomy
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 298-303







ABSTRACT
The aim of this study was to evaluate the toxicity of pea albumin 1 subunit b (PA1b) homologous against granary weevil (Sitophilus granarius L.). Adults of less than 5 days were maintained at 27±0.2°C and 70±5% relative humidity, on bread wheat (Triticum aestivum L. subsp. Aestivum) flour dumplings (control) and on pea flour, chickpea, bean, lentil, broad bean (toxic) during 15 days. The concentrations of used pea flour were 10, 20, 40, 60 and 80%. On PA1b reference: Pea/wheat with 80% pea flour, we have observed on the 3rd day, a mortality rate over 10%. From 7 days there were no survivors. For concentrations of 40 and 60%, the mortality rate was 73 and 93%, respectively; but at 10 and 20% of pea, the rate was low not exceeding the 20% threshold. A significantly positive correlation (R2 = 0.91) was observed between the concentration of pea flour extract and the mortality rate of Sitophilus granarius L. The linear regression of semi-logarithmic curve is highly significant (p = 0.012, <0.05) and allow to determine the LD50 of which is 25.63±3.26% or 64.075±8.14 mg. At the maximum concentration of 80% on lentil, bean and broad bean, there was a total mortality of the insect at 7 days; others are less toxic such as pea with 30% and chickpea, whose mortality rate does not exceed the 23% threshold. Furthermore, complete mortality of the insect was observed after 14 days in all grain legumes.
PDF Abstract XML References Citation
Received: July 31, 2015;
Accepted: September 02, 2015;
Published: November 11, 2015
How to cite this article
A. Mebarkia, Y. Abdi, S. Rahmani and A. Abjabi, 2015. Insecticidal Activity of Pea Albumin 1b Homologous against Granary Weevil (Sitophilus granarius L.). Journal of Agronomy, 14: 298-303.
DOI: 10.3923/ja.2015.298.303
URL: https://scialert.net/abstract/?doi=ja.2015.298.303
DOI: 10.3923/ja.2015.298.303
URL: https://scialert.net/abstract/?doi=ja.2015.298.303
INTRODUCTION
Wheat is the most important food crop for more than half of the world’s population. Losses in wheat due to insect pests, from harvest to consumption, affect food availability for large number of people (Abumhadi et al., 2012). Control measures of these insect populations around the world are mainly based on continued applications of synthetic insecticides (Shaaya et al., 1997; Daglish, 2004; Islam et al., 2010; Ali et al., 2012). These treatments are effective, inexpensive and readily available in developing countries where 90% of world production of wheat is performed (De Kouassi, 2001). The constant use of these molecules have caused selection of resistances to insecticides in agricultural pests, environmental pollution with negative side effects on human health and on non-target arthropods (Khambay et al., 2003; Desneux et al., 2007; Pimentel et al., 2009; Raj et al., 2013). Due to this awareness, there has been increasing worldwide interest in the development of alternative and sustainable means for modern pest management strategies (Rajendran and Sriranjini, 2008; Aliakbarpour et al., 2011), including new biological insecticides (Grewal et al., 2010) . These were evaluated from the higher plants (McLaren, 1986; Cantrell et al., 2012). Thus, the insecticidal activity of many plant derivatives, against several stored-product pests has been demonstrated (Su et al., 1972; Jilani and Su, 1983; Malik and Naqvi, 1984; Saxena et al., 1992; Golob, 1999; Nathan et al., 2007; Prakash et al., 2008; Regnault-Roger et al., 2012). But, the search for entomotoxic components in plant and specifically in plants that are consumed by mammals, could be a valuable approach in order to develop biopesticides for sustainable and healthy agriculture (Gressent et al., 2011). Several studies have reported that legume seeds contain a wide range of allelochemicals with toxic and deterrent effects against insect pests. The insecticide effect of pea flour on rice weevil (Sitophilus oryzae L.) have received the most attention (Hou and Fields, 2003; Pretheep-Kumar et al., 2004; Fields, 2006). Pea protein is repellent to several stored-product insects (Fields et al., 2001; Kumar et al., 2004) and toxic (Pretheep-Kumar et al., 2004). This toxicity is primarily correlated with the biological activity of a major albumin isolated from pea grains, pea albumin1 subunit b (PA1b) (Gressent et al., 2003). The entomotoxic high activity of PA1b is also coupled to a high stability of this protein. It is able to retain its biological activity during several years in dry seeds (Petit et al., 2005). The high toxicity of PA1b protein towards cereal weevils, its stability during storage and drying of the seeds, associated with the fact that it is consumed in quantity by man and animals, without signs of toxicity or allergenicity, make this peptide as an ideal candidate to produce transgenic plants, especially cereals, resistant to stocks beetles (Taylor et al., 2004; Louis et al., 2007). In addition, these authors noted that the existence of several isoforms of PA1b protein in pea as in other Fabaceae permits to define the PA1b protein as a new class of plant entomotoxin.
MATERIALS AND METHODS
This study was carried out at laboratory of Setif University during 2011-2014. The local strain of Sitophilus granarius L. was provided by storage cells of the cooperative of cereal and dry legumes of Setif. This specie was kept in laboratory culture for over 3 years and was reared on wheat seeds (Triticum aestivum L. subsp. Aestivum) and maintained at 27±0.2°C and 70±5% RH in the dark cited in Charles, 1997.
Plant material: It was made of wheat seeds (Triticum aestivum L. subsp. aestivum var. Siete Ceros.) and legume seeds species: Pea (Pisum sativum L. var. Languedoc.), Chickpea (Cicer arietinum L. var. Kabuli) bean (Phaseolus vulgaris L. var. Bordj Ménaïl), lentil (Lens esculenta M. var. Larissa) and broad bean (Vicia fabae L. var. Sidi Moussa 178/25). These varieties come from the different stations of the Technical Field Crop Institute (TFCI) of Algeria. Round mesh plastic cylinders (8 cm high and 11 cm diameter) were filled ¾ with seeds. They were put in closed containers at -20°C for a few days to destroy any insects that may be present. They were then stored at +4°C until use.
Preparation of flour: Pea seeds were finely ground in a mill and sieved on an AFNOR NF X 11-201 Prolabo sieve of 0.2 mm mesh size. Milling was continued so as to have the minimum non-sieving flour. Green flour thus obtained was used for extractions and preparation of wheat flour dumplings enriched pea flour. The same procedure was used for preparation of wheat flour and other grain legumes.
Preparation of artificial food: As described by Wicker (1984) cited in Delobel and Grenier (1993) improved by Gressent et al. (2003) for the preparation of the artificial food in a desired content in pea, we have weighed into an Eppendorf tube×mg of pea flour extract in milligram (PA1b), we have added 167 μL of ultra pure water and then we are completed the liquid sample with wheat flour by means a tube neck collar transfer for rod of 250 mg subsequently cut sliced (dumplings). For a concentration of 10% requires 25 mg of pea flour+225 mg of wheat flour. The assembly (Pea+water + wheat) is mixed well with a spatula. Cylinders of 0.3-0.5 cm in diameter and 5-10 cm long are shaped and dried for about 1 h at ambient laboratory temperature. They are then cut into slices of 0.3-0.5 cm. Then we allowed them to dry overnight at room temperature of laboratory without yeast or fermentation. Dumplings are then stored at +4°C in closed container until use. For the implementation of the test, 30 wheat weevil adults less than 5 days are placed in a tube of wheat flour dumplings (control) mixed with pea flour (reference toxicant) or other grain legumes, at concentration of 10, 20, 40, 60 and 80%. Cylinders are then placed in a controlled chamber at 27±0.2°C and 70±5% RH in the dark. These tests have 3 repetitions. Dead insects are counted and removed from the 3rd day for the study of mortality kinetics until the last survivor. The same process is carried out with maximal concentration of 80% flour for each kind of legumes. Data processing is performed by software XLStat.v7. Linear regression analysis was applied to define all dose-response relationships when correlation was found to be significant and the lethal dose for 50% of mortality is determined by the method of Miller and Tainter (1944) cited in Randhawa (2009). Analysis of variance (ANOVA) was applied to repellency data which were not significantly correlated with concentrations.
RESULTS
Pea A1b toxicity: As shown in Fig. 1, that after 2 days of Sitophilus granarius L. feeding on wheat (Triticum aestivum L. subsp. Aestivum) and pea (Pisum sativum L.), there are virtually no mortality. On the third day, it noted in general one to two deaths on wheat as the pea. Beyond, this mortality has differences in function of the pea concentration. On bread wheat, it noted no increase in mortality during the 15 days period of the experiment. On pea, for a concentration of 80% (PA1b), we observed from the third day a rapid increase in mortality exceeding 10%. From 7 days, there are virtually no survivors. For concentrations of 40 and 60%, mortality rates were 73 and 93%, respectively; by against for other concentrations, rates are low, not exceeding the 20% threshold. In addition, the graphical analysis of regression supports this observation, show that the curve of percentage mortality of Sitophilus granarius L. (probit units) versus the logarithm of the doses administered (percentage units) fits well with a linear curve of the first degree for the species (y = 4.856x-1.841).
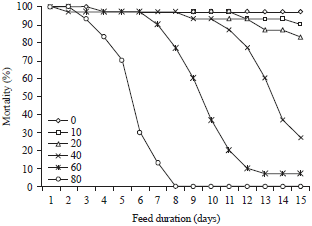 | |
| Fig. 1: | Cumulative mortality (%) of Sitophilus granarius L. adults on wheat (control) and mixture of wheat and pea (toxic), depending on pea concentrations (10, 20, 40, 60 and 80%) and the feed duration (Average of 3 repetitions) |
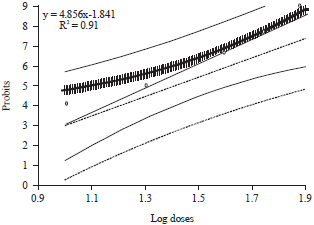 | |
| Fig. 2: | Curve expressing mortality of Sitophilus granarius L. (probit units) versus the logarithm of the dose (percentage units) |
This regression is highly significant (p = 0.012 <0.05), with a coefficient of determination R2 = 0.91 (Fig. 2). Linearization of this semi-logarithmic curve to determine the LD50 of which is 25.63±3.26%, or 64.075±8.14 mg.
Toxicity of PA1b homologous of legume seeds: We noted that all tested pulses are toxic to the Sitophilus granarius L. species and total mortality of the insect is recorded after 14 days in different species of grain legumes: Lentil (Lens esculenta M.), chickpea (Cicer arietinum L.), bean (Phaseolus vulgaris L.), broad bean (Vicia fabae L.) and pea (Pisum sativum L.), at concentration of 80% flour of these legumes (Fig. 3). We noted, however, differences in toxicity, considering the deaths before 14 days. Thus, at 7 days, the mortality rate is 100% on broad bean (Vicia fabae L.), bean (Phaseolus vulgaris L.) and lentil (Lens esculenta M.), while individuals are still alive on chickpea (Cicer arietinum L.) and peas (Pisum sativum L.).
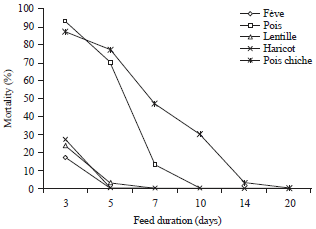 | |
| Fig. 3: | Cumulative mortality of Sitophilus granarius L. adults on the legumes species with a concentration of 80% as a function of feed duration (Average of 3 repetitions) |
| Table 1: | Analysis of the variance of mortality rate of Sitophilus granarius L. depending on the species of legumes and of the feed duration in days |
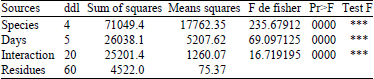 | |
| ***Highly significant to the error threshold 0.001 | |
The difference is seen much more clearly when we consider the mortality to 5 days. After this feed time, some species are toxic enough to cause mortality exceeds 96%. This is the case of the lentil (Lens esculenta M.), bean (Phaseolus vulgaris L.) and broad bean (Vicia fabae L.). Others are less toxic such as pea (Pisum sativum L.) with 30% and chickpea (Cicer arietinum L.), whose the mortality rate does not exceed the 23% threshold. Finally, it should be noted that only the chickpea (Cicer arietinum L.) requires quite a long time of about 20 days to reach maximum toxicity. Finally, it should be noted that only the chickpea (Cicer arietinum L.) requires a long time of about 20 days to reach a maximum mortality. Thus, the statistical analysis of variance (Table 1) shows that the difference between the averages of the species factors, days and the interaction between the two are very highly significant at 0.001 level errors and thus, have an effect on mortality of Sitophilus granarius L. In addition, Table 2 shows firstly that the ranking of average based on legume species has no significant difference at the 0.05 error threshold between the species of bean (Phaseolus vulgaris L.) lentil (Lens esculenta M.) and broad bean (Vicia fabae L.) but presents a significant difference between the species of chickpea (Cicer arietinum L.) and pea (Pisum sativum L.). Thus, the means are classified into three distinct groups and in a second time, it shows that there is a significant difference between the feeding times to the error threshold of 0.05 and thus, the averages are classified into 4 distinct groups.
| Table 2: | Classification of averages depending on the species of legumes and the feed duration (days) |
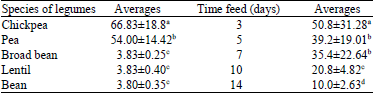 | |
| Different letters in brackets symbolize a significant difference to the error threshold of 0.05 | |
DISCUSSION
Pea A1bToxicity: A significantly positive correlation R2 = 0.91 was observed between the concentration of pea flour extract and the mortality rate of Sitophilus granarius L., accordance with the results announced by Fields et al. (2001). Thus, protein-rich pea flour is toxic to the wheat weevil (Hou et al., 2006). According to Fields (2006), the Sitophilus spp. is the most sensitive of the tested insects, followed by Cryptolestes ferrugineus, Tribolium castaneum and Rhyzopertha dominica. This was confirmed by the work of Hou et al. (2004), who noted that the wheat treatment with the pea flour enriched with protein, at a concentration of 0.04-0.1%, reduced the Sitophilus oryzae L. population of 26 and 79%, respectively and Cryptolestes ferrugineus of 27 and 43%, respectively. To a concentration of 1%, the death rate of Sitophilus oryzae L. reached 100% for 3 weeks on rice (Pretheep-Kumar et al., 2004). In addition, to the efficiency of the pea protein on reducing populations of Cryptolestes ferrugineus, Sitophilus spp., Tribolium castaneum and Tribolium confusum, Fields et al. (2001) noted that it is also repellent to concentrations of 0.1 and 1%, after 1 h of exposure. This repulsion is about 91.2% after 48 h of Sitophilus oryzae L. exposure to the treated rice at a concentration of 1% (Pretheep-Kumar et al., 2004). However, Hou et al. (2006) showed that the toxins of protein-rich pea flour can cross the insect cuticle and therefore the midgut tissue will be damaged with the presence of several gas bubbles. This sensitivity was determined by Gressent and his team, for the discovery of a high affinity binding site (Kd 2.6 nm) for the PA1b protein in membrane protein extracts of sensitive weevils and that seems absent or mutated in the resistant strains. The correlation between the presence of the binding site and the sensitivity to PA1b entomotoxin in weevils suggests that this site able to fix PA1b on its receptor might play a major role in the mechanism of toxicity (Gressent et al., 2003). These same authors noted that this protein binding site has been demonstrated in Coleoptera, Lepidoptera, Diptera, Hymenoptera and Hemiptera. However, the presence of this binding site does not necessarily imply that the insect will be sensitive to the PA1b protein.
Toxicity of Pea A1b homologous of legume seeds: These results are in agreement with those reported by Louis et al. (2004), where the toxicity of pea, soya bean, bean and alfalfa differs greatly from one plant species to another and at different concentrations (10, 20, 40, 60 and 80%). In fact, although in all cases, all the rice weevils (Sitophilus oryzae L.) die in 14 days regardless of the flour concentration. This report was also observed in individuals of Sitophilus granarius L. where we can say that the officer is the purified toxic protein. But it is not possible that the effects are combined with those of other secondary substances in these plant species as feeding deterrent and toxic effects because only 3 days of feeding, Vicia fabae L., Phaseolus vulgaris L. and Lens esculenta M. kill almost all insects, while it is only at this time that Pisum sativum L. begins to show its toxic effects and we are observed deaths on that food.
CONCLUSION
It is common for stored-product insects have large differences in sensitivity to synthetic insecticides. There have been few researches on the causes of these differences in sensitivity but they are probably due to the insecticide absorption differences, degradation inside the insect and mode of action. Therefore it is not surprising that there are large differences in the sensitivity to pea flour rich in protein between different pests of stored grains. The toxicity of PA1b protein and homologous of different species of legumes (bean, pea, chickpea and broad bean) was tested on granary weevil. It notes, however, differences in toxicity between these food legumes, probably because of differences between the homologous of PA1b and the presence of secondary substances. In anticipation of likely changes in legislation, insecticides should be replaced by physical methods such as silos cooling, storage under CO2 or nitrogen. These methods are difficult and expensive, they require highly technical and are not applicable everywhere. Another alternative is to make the seeds resistant to insects through genetic modification. This vegetable protein is everywhere but at varying levels. She comes from a plant and a tissue regularly consumed by man, which minimizes the risk of toxicity, even if it exists, it must be very small. Therefore, it should be valued in the context of use with plant transgenesis which could also protect against many insects. Given that peas are toxic to cereal weevil, we investigated the repellency of homologous pea against some stored-product insects. In sum, biopesticides represent one of the best alternatives to chemicals for the development of environmental friendly and safe strategies for pest management of stored cereal.
ACKNOWLEDGMENTS
I would like to thank the officials of the various stations of the Technical Institute of Field Crops (TIFC) of Algeria for legume seeds, Cooperative of Cereals and Dry Legumes (CCDL) of Setif for providing us the wheat grains and the all team of functional biology laboratory INRA-INSA of Lyon, France for their contribution to achievement this work.
REFERENCES
- Ali, A., F. Ahmad, A. Biondi, Y. Wang and N. Desneux, 2012. Potential for using Datura alba leaf extracts against two major stored grain pests, the khapra beetle Trogoderma granarium and the rice weevil Sitophillus oryzae. J. Pest Sci., 85: 359-366.
CrossRefDirect Link - Abumhadi, N., E. Todorovska, B. Assenov, S. Tsonev and D. Vulcheva et al., 2012. Agricultural research in 21st century: Challenges facing the food security under the impacts of climate change. Bulgarian J. Agric. Sci., 18: 801-818.
Direct Link - Aliakbarpour, H., M.R.C. Salmah and O. Dzolkhifli, 2011. Efficacy of neem oil against thrips (Thysanoptera) on mango panicles and its compatibility with mango pollinators. J. Pest Sci., 84: 503-512.
CrossRefDirect Link - Khambay, B.P.S., D. Batty, P.J. Jewess, G.L. Bateman and D.W. Hollomon, 2003. Mode of action and pesticidal activity of the natural product dunnione and of some analogues. Pest Manage. Sci., 59: 174-182.
CrossRefDirect Link - Cantrell, C.L., F.E. Dayan and S.O. Duke, 2012. Natural products as sources for new pesticides. J. Nat. Prod., 75: 1231-1242.
CrossRefDirect Link - Daglish, G.J., 2004. Effect of exposure period on degree of dominance of phosphine resistance in adults of Rhyzopertha dominica (Coleoptera: Bostrychidae) and Sitophilus oryzae (Coleoptera: Curculionidae). Pest Manage. Sci., 60: 822-826.
CrossRefDirect Link - Delobel, B. and A.M. Grenier, 1993. Effect of non-cereal food on cereal weevils and tamarind pod weevil (Coleoptera: Curculionidae). J. Stored Prod. Res., 29: 7-14.
CrossRefDirect Link - Desneux, N., A. Decourtye and J.M. Delpuech, 2007. The sublethal effects of pesticides on beneficial arthropods. Annu. Rev. Entomol., 52: 81-106.
CrossRefPubMedDirect Link - Fields, P.G., Y.S. Xie and X. Hou, 2001. Repellent effect of pea (Pisum sativum) fractions against stored-product insects. J. Stored Prod. Res., 37: 359-370.
CrossRefDirect Link - Fields, P.G., 2006. Effect of Pisum sativum fractions on the mortality and progeny production of nine stored-grain beetles. J. Stored Prod. Res., 42: 86-96.
CrossRefDirect Link - Golob, P., C. Moss, M. Dales, A. Fidgen, J. Evans and I. Gudrups, 1999. The Use of Spices and Medicinals as Bioactive Protectants for Grains (FAO Agricultural Services Bulletin 137). Food and Agriculture Organization, Rome, Italy, ISBN-13: 9789251042946, Pages: 239.
Direct Link - Gressent, F., I. Rahioui and Y. Rahbe, 2003. Characterization of a high-affinity binding site for the pea albumin 1b entomotoxin in the weevil Sitophilus. Eur. J. Biochem., 270: 2429-2435.
CrossRefDirect Link - Gressent, F., P. Da Silva, V. Eyraud, L. Karaki and C. Royer, 2011. Pea albumin 1 subunit b (PA1b), a promising bioinsecticide of plant origin. Toxins, 3: 1502-1517.
CrossRefDirect Link - Grewal, K.K., G.S. Sandhu, R. Kaur, R.S. Brar and H.S. Sandhu, 2010. Toxic impacts of cypermethrin on behavior and histology of certain tissues of albino rats. Toxicol. Int., 17: 94-98.
PubMedDirect Link - Hou, X. and P. Fields, 2003. Granary trial of protein-enriched pea flour for the control of three stored-product insects in barley. J. Econ. Entomol., 96: 1005-1015.
CrossRefDirect Link - Hou, X., P. Fields, P. Flinn, J. Perez-Mendoza and J. Baker, 2004. Control of stored-product beetles with combinations of protein-rich pea flour and parasitoids. Environ. Entomol., 33: 671-680.
CrossRefDirect Link - Hou, X., W. Taylor and P. Fields, 2006. Effect of pea flour and pea flour extracts on Sitophilus oryzae. Can. Entomol., 138: 95-103.
CrossRefDirect Link - Islam, M.S., M. Hasan, C.L. Lei, T. Mucha-Pelzer, I. Mewis and C. Ulrichs, 2010. Direct and admixture toxicity of diatomaceous earth and monoterpenoids against the storage pests Callosobruchus maculatus (F.) and Sitophilus oryzae (L.). J. Pest Sci., 83: 105-112.
CrossRefDirect Link - Jilani, G. and H.C.F. Su, 1983. Laboratory studies on several plant materials as insect repellants for protection of cereal grains. J. Econ. Entomol., 76: 154-157.
CrossRefDirect Link - Kumar, P.P., S. Mohan and G. Balasubramanian, 2004. Effect of whole-pea flour and a protein-rich fraction as repellents against stored-product insects. J. Stored Prod. Res., 40: 547-552.
CrossRefDirect Link - Louis, S., B. Delobel, F. Gressent, G. Duport and O. Diol et al., 2007. Broad screening of the legume family for variability in seed insecticidal activities and for the occurrence of the A1b-like knottin peptide entomotoxins. Phytochemistry, 68: 521-535.
CrossRefDirect Link - Louis, S., B. Delobel, F. Gressent, I. Rahioui, L. Quillien, A. Vallier and Y. Rahbe, 2004. Molecular and biological screening for insect-toxic seed albumins from four legume species. Plant Sci., 167: 705-714.
CrossRefDirect Link - Malik, M.M. and S.H.M. Naqvi, 1984. Screening of some indigenous plants as repellents or antifeedants for stored grain insects. J. Stored Prod. Res., 20: 41-44.
CrossRefDirect Link - McLaren, J.S., 1986. Biologically active substances from higher plants: Status and future potential. Pestic. Sci., 17: 559-578.
CrossRefDirect Link - Nathan, S.S., M.Y. Choi, C.H. Paik, H.Y. Seo, J.D. Kim and S.M. Kang, 2007. The toxic effects of neem extract and azadirachtin on the brown planthopper, Nilaparvata lugens (Stal) (BPH) (Homoptera: Delphacidae). Chemosphere, 67: 80-88.
CrossRefPubMedDirect Link - Petit, J., G. Duport, Y. Rahbe, E. Guiderdoni and J.C. Breitler, 2005. Pea albumin 1b (PA1b), an entomotoxic cystein-rich peptide protects transgenic rice seeds against the stored product pest Sitophilus oryzae L. Proceedings of the 5th International Rice Genetics Symposium and 3rd International Rice Functional Genomics Symposium, November 19-23, 2005, Manila, Philippines.
- Pimentel, M.A.G., L.R.D.A. Faroni, R.N.C. Guedes, A.H. Sousa and M.R. Totola, 2009. Phosphine resistance in Brazilian populations of Sitophilus zeamais Motschulsky (Coleoptera: Curculionidae). J. Stored Prod. Res., 45: 71-74.
CrossRefDirect Link - Prakash, A., J. Rao and V. Nandagopal, 2008. Future of botanical pesticides in rice, wheat, pulses and vegetables pest management. J. Biopesticides, 1: 154-169.
Direct Link - Pretheep-Kumar, P., S. Mohan and K. Ramaraju, 2004. Protein-enriched pea flour extract protects stored milled rice against the rice weevil, Sitophilus oryzae. J. Insect. Sci., 4: 26-32.
CrossRefDirect Link - Raj, J., M. Chandra, T.D. Dogra, M. Pahuja and A. Raina, 2013. Determination of median lethal dose of combination of endosulfan and cypermethrin in wistar rat. Toxicol. Int., 20: 1-5.
CrossRefPubMedDirect Link - Rajendran, S. and V. Sriranjini, 2008. Plant products as fumigants for stored-product insect control. J. Stored Prod. Res., 44: 126-135.
CrossRefDirect Link - Randhawa, M.A., 2009. Calculation of LD50 values from the method of Miller and Tainter, 1944. J. Ayub. Med. Coll. Abbottabad, 21: 184-185.
PubMedDirect Link - Regnault-Roger, C., C. Vincent and J.T. Arnason, 2012. Essential oils in insect control: Low-risk products in a high-stakes world. Annu. Rev. Entomol., 57: 405-424.
CrossRefDirect Link - Saxena, R.C., O.P. Dixit and V. Harshan, 1992. Insecticidal action of Lantana camara against Callosobruchus chinensis (Coleoptera: Bruchidae). J. Stored Prod. Res., 28: 279-281.
CrossRefDirect Link - Shaaya, E., M. Kostjukovski, J. Eilberg and C. Sukprakarn, 1997. Plant oils as fumigants and contact insecticides for the control of stored-product insects. J. Stored Prod. Res., 33: 7-15.
CrossRefDirect Link - Su, H.C.F., R.D. Speirs and P.G. Mahany, 1972. Citrus oils as protectants of black-eyed peas against cowpea weevils: Laboratory evaluation. J. Econ. Entomol., 65: 1433-1436.
CrossRefDirect Link - Taylor, W.G., P.G. Fields and J.L. Elder, 2004. Insecticidal components from field pea extracts: Isolation and separation of peptide mixtures related to pea albumin 1b. J. Agric. Food Chem., 52: 7491-7498.
CrossRefDirect Link - De Kouassi, M., 2001. Les possibilites de la lutte microbiologique: Emphase sur le champignon entomopathogene B. bassiana. Revue Electronique Sciences l'Environnement, Vol. 2, No. 2, (In French).
CrossRefDirect Link