
Morteza Kamrani
Department of Plant Production, Moghan College of Agriculture and Natural Resource, University of Mohaghegh Ardabili, Ardabil, Iran
Journal of Agronomy
Year: 2015 | Volume: 14 | Issue: 4 | Page No.: 254-263







ABSTRACT
Drought is the most devastating abiotic stress factor worldwide. It severely limits plant growth and development as well as plant performance. In order to study the effect of drought stress on the relationship among agro-morphological traits, 34 bread wheat genotypes were tested in a randomized complete block design with three replications under irrigated and rainfed conditions during 2012-13 and 2013-14 cropping seasons. The results of combined analysis of variance showed that the effect of environment, genotype and genotype×environment interaction were significant (p<0.01) for all studied traits. Biplot analysis revealed that Harvest Index (HI), Kernel Weight per Spike (KWS), Thousand Kernel Weight (TKW), Flag Leaf Weight (FLW), Flag Leaf Area (FLA), Days to Heading (DTH), Days to Maturity (DTM), Peduncle Length (PL), Plant Height (PH) and Biological Yield (BY) in irrigated and the number of kernel per spike (NKS), chlorophyll content (CHL), FLA, FLW, BY and DTM in rainfed condition had positive effects on Grain Yield (GY). The results of correlation and biplot analyses showed that STI, GMP and MP indices were positively correlated with grain yield under both irrigated and rainfed conditions. Based on the results, the superior wheat genotypes identified were G13 and G7. These genotypes thus may serve as parents for drought tolerance improvement.
PDF Abstract XML References Citation
Received: September 16, 2015;
Accepted: October 22, 2015;
Published: November 11, 2015
How to cite this article
Morteza Kamrani, 2015. Relationship Among Agro-Morphological Traits in Bread Wheat (Triticum aestivum) Genotypes Under Irrigated and Rainfed Conditions. Journal of Agronomy, 14: 254-263.
DOI: 10.3923/ja.2015.254.263
URL: https://scialert.net/abstract/?doi=ja.2015.254.263
DOI: 10.3923/ja.2015.254.263
URL: https://scialert.net/abstract/?doi=ja.2015.254.263
INTRODUCTION
Bread wheat is one of the most important cereal crops in the world, which is grown both in arid and semi arid regions of the world (Tunio et al., 2006). World’s wheat production was about 704 million tons in 2011 (FAO., 2011). Iran is ranked as 14th in world wheat production. According to the recent reports, wheat was cultivated more than seven million ha and its total production was about 14.3 million tons in Iran, during 2010-2011 cropping season (FAO., 2011). Wheat is mainly grown on rainfed land and about 37% of the area of developing countries consists of semiarid environments in which available moisture constitutes a primary constraint on wheat production. Climatic variability in these marginal environments causes large annual fluctuations in yield (Rajaram, 2001). Improving drought tolerance is, therefore, a major objective in plant breeding programs for rainfed agriculture in these regions (Ehdaie and Waines, 1993). Evaluations of genotypes for drought tolerance have identified key sources of tolerance. Development of drought tolerant cultivars is hampered by low heritability of related traits and lack of effective selection strategies (Kirigwi et al., 2004). The basis of drought tolerance is complex and driven by diverse drought adaptive mechanisms, which are normally under multigenic control (Blum, 2005; Pinto et al., 2010). Grain yield is frequently used in wheat as a main criterion for drought tolerance. Selection for drought tolerance typically involves evaluating genotypes for either high yield potential or stable performance under varying degrees of water stress (Ahmad et al., 2003). However, grain yield actually is a product of several contributing factors and can be estimated on the basis of performance of various components. Therefore, progress has required combining measurements of yield-related traits associated with yield response and selection based on above yield components would result in yield increases.
In the most of field based experiments, drought tolerance indices, based on yield reduction under drought conditions in comparison to non-stress conditions are commonly used to identify drought tolerant genotypes (Mitra, 2001). The relative yield performance of genotypes in drought stressed and more favorable environments seems to be a common starting point in identification of traits related to drought tolerance and selection of genotypes for use in breeding for dry environments (Clark et al., 1992). The primary goal of many breeding programs is to identify superior genotypes through Multi Environment Trials (MET), considering multiple traits (Yan and Rajcan, 2002). Effective interpretation and utilization of the MET data are very important at all stages of plant breeding. Usually, a number of genotypes are tested across a number of sites and years and data on multiple traits recorded and it is often difficult to determine the pattern of genotypic performance across environments. Relationships among traits impact the choice of selection and breeding strategies. If all objective traits were positively correlated, selection would not be much more difficult than selecting for a single trait. If all objective traits were either positively correlated or independently inherited, selection would not be too difficult either. However, strong negative correlations between objective traits often exist, which make breeding very challenging (Lewis, 2006). Therefore, sufficient attention must be paid to undesirable associations among objective traits when performing independent culling, because selection for the desired levels or culling for the undesired levels of one trait can mean selection against the desired levels of another trait, which can lead to the loss of useful materials or even render the selection useless (Yan and Rajcan, 2002). The Genotype-by-Trait (GT) biplot proposed by Yan and Rajcan (2002) is a statistical tool for evaluating cultivars based on multiple traits and for identifying those that are superior in desired traits and hence could be candidates for use as parents in a breeding program or could be directly released for commercial production.
The objectives of the present study were to determine the relationships among agro-morphological traits of bread wheat under irrigated and drought conditions and to identify agro-morphological traits related to drought tolerance.
MATERIALS AND METHODS
Plant material and experimental conditions: Thirty four bread wheat (Triticum aestivum) genotypes (Table 1) from the wheat breeding project of International Maize and Wheat Improvement Center (CIMMYT) were evaluated during 2012-2013 and 2013-2014 growing seasons.
| Table 1: | Pedigree of bread wheat genotypes used in the experiment |
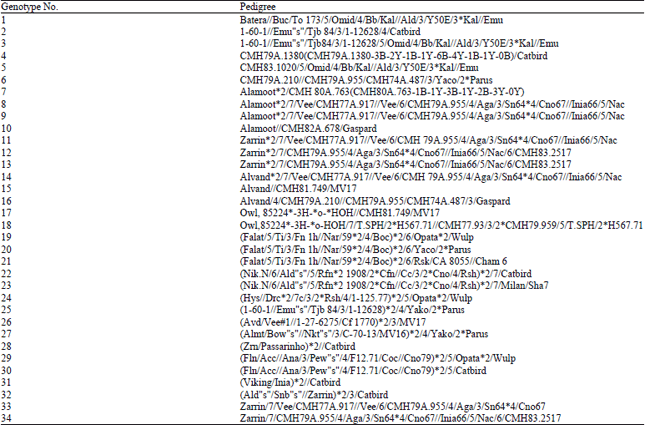 | |
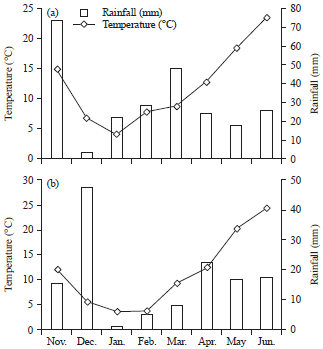 | |
| Fig. 1(a-b): | Rainfall and mean temperature for Moghan during (a) 2012-13 and (b) 2013-14 cropping seasons |
The field experimental design was a Randomized Complete Block Design (RCBD) with three replications under two contrasting irrigation (irrigated and rainfed conditions) regimes conducted at the research farm of Mohaghegh Ardabili University located at Moghan, Iran (39°39' N, 48°16' E and 32 masl). The genotypes were planted on 6 and 1 Nov in the 2011-2012 and 2012-2013 growing seasons, respectively, in 1.2×3 m plots, consisting of six rows 20 cm apart at 350 seeds m–2 for each condition. Based on a soil test before planting, 41 and 46 kg ha–1 of N and P2O5 were applied, respectively. The first irrigation was given at the time of late tillering. The subsequent irrigations were applied every 14 days. Irrigated plants received irrigation water and rainfall water, while non-irrigated plants only received rainfall water during the growing season (Nov.-Jun.). Rainfall and mean temperature of the experimental site is given in Fig. 1. Weed control in both growing seasons was conducted with an application of the herbicides clodinafop-propargyl at 0.7 L ha–1 and tribenuron-methyl at rate of 15 g ha–1.
Agronomic traits: Biological Yield (BY), Grain Yield (GY), Harvest Index (HI) and Thousand Kernel Weight (TKW) were measured by harvesting 1 m2 of the central part of each plot at crop maturity. Ten plants were randomly chosen from each plot to measure the Number of Kernel per Spike (NKS), Kernel Weight per Spike (KWS), Peduncle Length (PL), Plant Height (PH), chlorophyll content (CHL), Flag Leaf Weight (FLW), Flag Leaf Area (FLA), Days to Heading (DTH) and Days to Maturity (DTM).
Drought tolerance indices: Drought tolerance indices were calculated using the following relationships:
| • | Stress Tolerance Index (STI) = (Yp×ys)/( p)2 (Fernandez, 1993) |
| • | Mean Productivity (MP) = (Yp+Ys)/2 (Rosielle and Hamblin, 1981) |
| • | Geometric Mean Productivity (GMP) = (Yp×Ys)0.5 (Fernandez, 1993) |
| • | Stress Tolerance (TOL) = Yp-Ys (Rosielle and Hamblin, 1981) |
| • | Stress Susceptibility Index (SSI) = 1-(Ys/ p)/SI (Stress Intensity) SI = 1-( s/ p) (Fischer and Maurer, 1978) |
| • | Yield index (YI) = Ys/ s (Gavuzzi et al., 1997) |
| • | Yield Stability Index (YSI) = Ys/Yp (Bouslama and Schapaugh, 1984) |
In the above formulas, Ys, Yp, s and p represent grain yield under stress, grain yield under non-stress for each genotype, grain yield mean in stress and non-stress conditions for all genotypes, respectively.
Statistical analysis: Data was analyzed using SAS version 9.1 and GGEbiplot (Yan, 2001) statistical software. Analysis of variance (ANOVA) was performed to determine the effect of year, environment, genotype and their interactions on the traits. Prior to the combined ANOVA analysis, homoscedasticity of variance among environments was tested by the Levene test. Significant differences among genotypes were determined using Fisher’s protected LSD at 5% probability level. The genotype-by-trait biplot was used for studying relationships among traits and genotypes. Pearsons’correlation was conducted between drought tolerance indices and traits.
RESULTS AND DISCUSSION
Analysis of variance: The results of combined analysis of variance revealed that there were no significant differences between the two studied years for all measured traits (Table 2), hence, data averaged across the two seasons was used for means comparisons. The effect of environment, genotype and genotype×environment interaction were significant (p<0.01) for all measured traits. Traits were not affected by year×environment interaction. These results indicate that studied genotypes responded differently to the different environmental conditions suggesting the importance of the assessment of genotypes under different environments in order to identify the best genetic make up for a particular environment. The traits NKS, KWS, TKW, HI and CHL were affected by genotype×year interaction (p<0.01). The effect of genotype×year×environment interaction was significant (p<0.01) for BY, GY, TKW and CHL traits (Table 2).
| Table 2: | Combined analysis of variance for agronomic traits and grain yield of wheat genotypes |
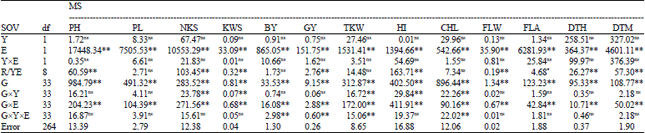 | |
| nsNot significant, **Significant at 0.01 probability levels, BY: Biological yield, GY: Grain yield, HI: Harvest index, TKW: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: Flag leaf area, DTH: Days to heading, DTM: Days to maturity | |
| Table 3: | Mean of measured agronomic traits for 34 wheat genotypes under irrigated condition (averaged over 2 years) |
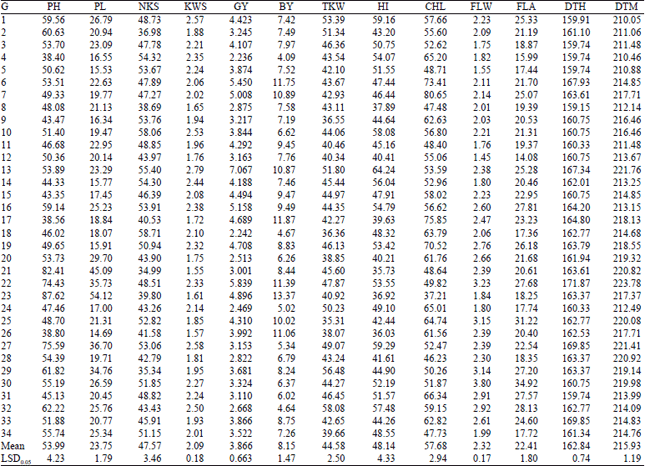 | |
| BY: Biological yield, GY: Grain yield, HI: Harvest index, TWK: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: Flag leaf area, DTH: Days to heading, DTM: Days to maturity, G: Genotypes | |
Comparisons of mean for traits: Means of grain yield and agronomic traits of genotypes in irrigated and rainfed conditions are given in Table 3 and 4, respectively. Results for grain yield depict that the highest and lowest grain yield belonged to the genotypes G13 (7.066 t ha–1) and G4 (2.236 t ha–1) under irrigated condition (Table 3). Under rainfed condition, genotypes G7 (4.996 t ha–1) and G4 (1.228 t ha–1) had the highest and lowest grain yield, respectively. Drought stress reduced biological yield of all genotypes significantly. Genotypes G23 (13.369 t ha–1), G17 (11.865 t ha–1) and G6 (11.748 t ha–1) had high biological yields and genotypes G4 (4.090 t ha–1), G32 (4.6 t35 ha–1) and G18 (4.666 t ha–1) had low biological yields under irrigated condition.
| Table 4: | Mean of measured agronomic traits for 34 wheat genotypes under rainfed condition (averaged over 2 years) |
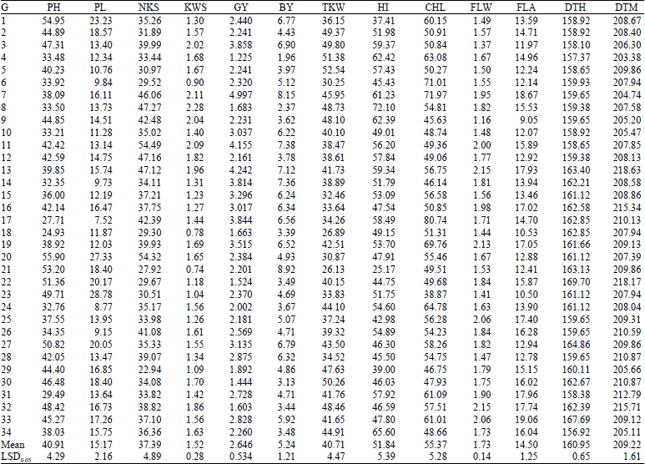 | |
| BY: Biological yield, GY: Grain yield, HI: Harvest index, TWK: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: Flag leaf area, DTH: Days to heading, DTM: Days to maturity, G: Genotypes | |
Under rainfed condition, genotypes G21 (8.921 t ha–1), G7 (8.149 t ha–1) and G11 (7.379 t ha–1) had high and genotypes G4 (4.090 t ha–1), G32 (4.6 t35 ha–1) and G18 (4.666 t ha–1) had low biological yields. The TKW, as an important yield grain component, also decreased significantly with drought stress (from 44.58 g under irrigated condition to 40.71 g under rainfed condition). Under irrigated condition the highest TKW value was observed for genotype G32 with 58.08 g and the lowest value for genotype G25 with 35.31 g while under rainfed condition highest TKW was assigned to genotype G5 with 52.53 g and the lowest TKW was observed in genotype G21 with 26.12 g. Under irrigated condition, the highest HI belonged to genotype G13 (64.24%) followed by genotypes G27 (59.28%) and G1 (59.15%) and the lowest value belonged to genotype G21 (35.73%) followed by genotypes G26 (36.03%) and G23 (36.91%). Under rainfed condition, the highest HI was assigned to genotype G8 (72.10%) followed by genotypes G34 (65.59%) and G4 (62.41%) and the lowest HI was observed in genotype G21 (25.16%) followed by genotypes G1 (37.41%) and G29 (38.99%). Warrington et al. (1977) explained that drought stress at anthesis may lead to yield loss by reducing TKW and if accompanied with high temperatures after anthesis, accelerates whole plant senescence and reduces biological yield and HI. Bayoumi et al. (2008) and Dorostkar et al. (2015) observed that grain yield, biological yield, HI and TKW decreased under stress condition.
The number of the grains per spike is an important grain yield component. It has been reported that high yield in the durum wheat varieties are associated with the increasing number of grain per spike (Calderini et al., 2006). The highest NKS under irrigated condition was recorded for G18 (58.71) followed by G10 (58.05) and G13 (55.40), while the lowest NKS was observed in G21 (34.98) followed by G29 (35.33) and G2 (36.98). Under rainfed condition, the highest NKS belonged to G11 (54.48) followed by G20 (54.32) and G8 (47.27). The lowest NKS belonged to G29 (22.94) followed by G21 (27.92) and G18 (29.30). Severe water stress from the seedling stage to maturity reportedly reduced all grain yield components, particularly the number of grain per spike, dry matter and harvest index (Garcia del Moral et al., 2005).
In irrigated condition, genotypes G13 (2.79 g), G27 (2.58 g) and G1 (2.57 g) had the highest KWS and genotypes G21 (1.55 g), G26 (1.56 g) and G23 (1.60 g) showed the lowest KWS. In rainfed condition, genotypes G8 (2.28 g), G7 (2.10 g) and G11 (2.08 g) had the highest KWS and genotypes G21 (1.55 g), G26 (1.56 g) and G23 (1.60 g) showed the lowest KWS. Drought stress reduced plant height and peduncle length of genotypes significantly. Plant height ranged from 38.39 cm (G4) to 87.61 cm (G23) under irrigated condition and from 24.93 cm (G18) to 55.89 cm (G20) under rainfed condition. Generally, drought stress caused a decrease in PH, up to 8.80 cm for the means of genotypes. Richards et al. (2001) have reported that one of the major effects of drought stress is to decrease plant height, which also caused a reduction in dry matter accumulation and subsequently plant production.
Peduncle length ranged from 14.68 cm (G26) to 54.12 cm (G23) under irrigated condition and from 7.52 cm (G17) to 28.78 cm (G23) under rainfed condition. Drought stress decreased the mean PL for 34 genotypes by 13.60 cm. Plaut et al. (2004) reported that the peduncle length of wheat was reduced in the drought stress.
Drought stress caused an obvious decrease in FLW across all 34 genotypes. The mean FLW of 34 genotypes under irrigated and rainfed condition was 2.32 and 1.73 g, respectively. Drought stress resulted in decrease in FLA. Under irrigated condition, genotypes G30, G25 and G32 had bigger FLA, while G12, G4 and G18 smaller FLA. Under rainfed condition, genotypes G33, G7 and G31 had bigger FLA, while G9, G23 and G18 had smaller FLA. Blum (2005) suggested that a small flag leaf area is beneficial under drought stress due to being dehydration avoidant.
Chlorophyll content (CHL) was decreased significantly under drought stress. Under irrigated condition the highest CHL value was observed for genotype G7 followed by G17 and G6 and the lowest value for genotype G23 followed by G28 and G8, while under rainfed condition highest CHL was assigned to genotype G17 followed by G7 and G6 and the lowest CHL was observed in genotype G23 followed by G9 and G14. Similar results were observed by Sayar et al. (2008) in wheat and Lonbani and Arzani (2011) in wheat and triticale. Drought stress accelerated days to heading for all genotypes. The extent of acceleration was different among genotypes. Under irrigated and rainfed condition, the mean DTH of all genotypes was 162.84 and 160.95 days, respectively. Under irrigated condition, genotypes G8, G4 and G5 had earlier DTH, while G22, G33 and G27 had later DTH. Genotypes G34, G4 and G3 showed the earliest DTH, while G22, G33 and G27 were the latest DTH under rainfed condition. Drought stress affected days to maturity among the 34 genotypes. The DTM under rainfed condition was earlier than that under irrigated condition. Drought stress caused 7 days acceleration in the mean DTM of 34 genotypes. Under irrigated condition, genotypes G1, G4 and G5 had earlier DTM, while G22, G13 and G27 had later DTM. Genotypes G4, G7 and G34 had earlier DTM, while G13, G22 and G32 were the latest DTM under rainfed condition. Kilic and Yagbasanlar (2010) reported that the number of days to heading and the number of days to maturity were reduced under drought stress condition.
Visualization of relationships among traits: To visualize the interrelationships among studied traits of 34 genotypes in each environments, genotype by trait biplots are presented in Fig. 2. The biplots were based on the first two principal components derived from subjecting the standardized genotype-by-trait table. In the biplots, a vector is drawn from the biplot origin to each marker of the traits to facilitate visualization of the interrelationship among the studied traits. The biplot explained 53 and 48% of the total variation of standardized data for irrigated and rainfed conditions, respectively. A relatively low proportion of the total variation explained by the biplots indicates the complexity of the interrelationship among the traits (Yan and Rajcan, 2002).
Under irrigated condition, HI, KWS, TKW, FLA, FLW, DTH, DTM, PH, PL and BY constituted a group of traits with positive effects on GY (indicated by acute angles between vectors for those traits and GY), while CHL had a negative effect on GY, as indicated by the obtuse angles between their vectors. A near zero correlation was between NKS and GY as indicated by the near perpendicular vectors. Strong positive correlations were observed among NKS, KWS and HI and among PH, PL and BY traits. Chlorophyll content (CHL), NKS, KWS and HI were negatively correlated with PH, PL and BY.
Relationships among traits under irrigated condition were different from those under rainfed condition, which suggested that there was differential response of genotypes to the two environments. Under rainfed condition, NKS, CHL, FLA, FLW, BY and DTM had positive effects on GY (indicated by acute angles between vectors for those traits and GY). KWS and DTH were less positively correlated with GY. The TKW, HI, PH and PL were not correlated with GY as indicated by the right angle between their vectors. NKS, KWS, HI and TKW were negatively correlated with PH and PL. Other relations revealed from the genotype by trait biplot include positive associations among KWS, HI and TKW, between PH and PL and among DTH, DTM and BY. Genotype by trait biplot provides more information on interrelationship among traits than other commonly used methods, such as path coefficient analysis (Rubio et al., 2004). Fernandez-Aparicio et al. (2009) demonstrated that the genotype by trait biplots were an excellent tool for visualizing genotype-by-trait data and revealing the interrelationships among traits.
The polygon view of GT biplot allows visualization of the which-won-where pattern that identifies genotypes that are best for certain traits and it is also used as independent selection criteria based on several traits (Yan and Rajcan, 2002).
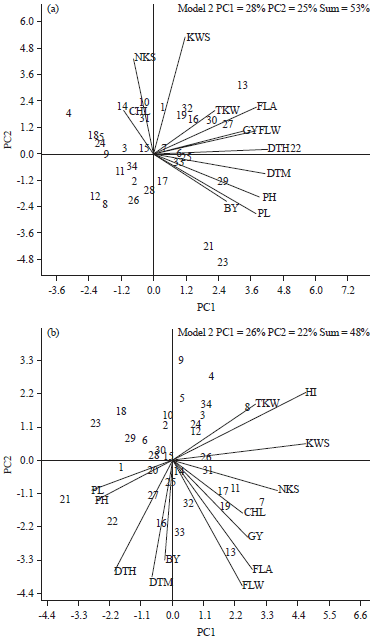 | |
| Fig. 2(a-b): | Vector view of genotype by trait biplot which showing relationship among the traits under (a) Irrigated and (b) Rainfed conditions. Numbers inside the figure are genotypes number. BY: Biological yield, GY: Grain yield, HI: Harvest index TKW: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: flag leaf area, DTH: Days to heading, DTM: Days to maturity |
Under irrigated conditions (Fig. 3a), genotype G22 had the highest values for FLA, FLW, GY, DTH, DTM and PH, whereas genotypes G21 and G23 had the highest values for PL and BY. Genotype G13 had the highest value for TKW. Under rainfed conditions (Fig. 3b), genotype G13 had the highest values for GY, FLA, FLW, BY and DTM. Genotype G8 had the highest value for HI and TKW. Genotype G7 had the highest value for KWS, NKS and CHL. Genotype G21 had the highest value for PH and PL.
Relationships between drought tolerance indices and traits: The result of correlation analysis (Table 5) showed that STI, GMP and MP indices were positively correlated with grain yield under both irrigated and rainfed conditions. Therefore, they can be used as suitable criteria for identifying drought tolerance in genotypes. The capability of the GMP and STI indices to identify genotypes satisfactorily under both conditions observed in this study is consistent with the results reported by Fernandez (1993) in mung bean and Nouri et al. (2011) in durum wheat. The TOL and SSI indices were positively correlated with grain yield under irrigated condition, while they were negatively correlated with grain yield under rainfed condition.
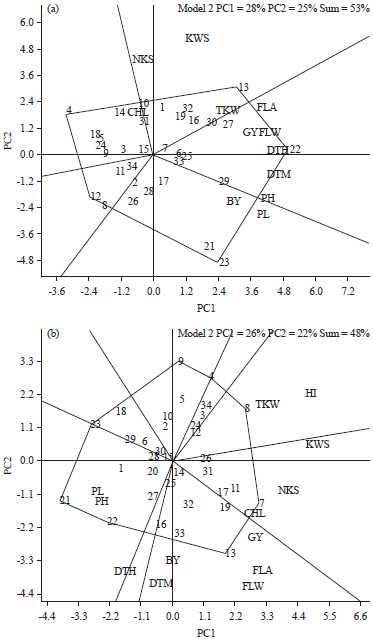 | |
| Fig. 3(a-b): | Polygon view of genotype by trait biplot of 34 wheat genotypes under (a) Irrigated and (b) Rainfed conditions. Numbers inside the figure are genotypes number (Table 1). BY: Biological yield, GY: Grain yield, HI: Harvest index, TKW: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: Flag leaf area, DTH: Days to heading, DTM: Days to maturity |
The observed relations were in agreement with those reported by Bahrami et al. (2014) in safflower and Nouri et al. (2011) in durum wheat. Fernandez (1993) reported that selection based on STI, GMP and MP would identify genotypes with higher levels of yield potential and stress tolerance, whereas, SSI and TOL indices had a significant negative correlation with seed yield under stress conditions but a positive correlation with seed yield under non-stress conditions. Therefore, selection based on SSI and TOL decreases grain yield under normal conditions but increases it under drought-stress conditions. The YSI was positively correlated with grain yield under rainfed condition, but it was negatively correlated with grain yield under irrigated condition. A positive correlation was found between YI and grain yield under both irrigated and rainfed conditions. In rainfed condition, NKS, KWS, BY, CHL, FLW, FLA and DTM traits were positively correlated with STI, GMP and MP indices, while TKW was negatively correlated with them. These results indicate that genotypes with high NKS, KWS, BY, CHL, FLW, FLA and DTM have high drought tolerance under rainfed condition. NKS, KWS and BY were positively correlated with STI, GMP and MP indices, while they were negatively correlated with TOL and SSI indices.
| Table 5: | Correlation coefficients between agronomic traits and drought tolerance indices under irrigated and rainfed conditions |
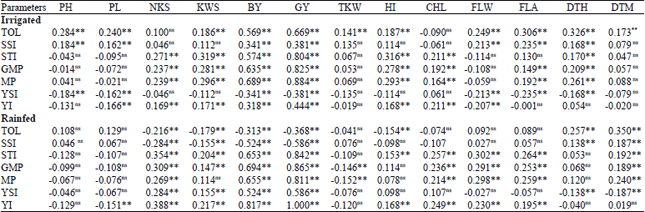 | |
| nsNot significant, **Significant at 0.01 probability levels, BY: Biological yield, GY: Grain yield, HI: Harvest index, TWK: Thousand-kernel weight, NKS: Number of kernel per spike, KWS: Kernel weight per spike, PL: Peduncle length, PH: Plant height, CHL: Chlorophyll content, FLW: Flag leaf weight, FLA: Flag leaf area, DTH: Days to heading, DTM: Days to maturity, TOL: Stress tolerance, SSI: Stress susceptibility index, STI: Stress tolerance index, GMP: Geometric mean productivity, MP: Mean productivity, YSI: Yield stability index, YI: Yield index | |
CONCLUSION
The results showed that drought stress reduced grain yield and agronomic traits except HI. Genotypes number G13 and G7 had higher grain yields under irrigated and rainfed conditions, respectively. The biplots for each condition were significantly different, indicating that the relationships among traits are significantly influenced by drought stress. Biplot analysis revealed that NKS, CHL, FLA, FLW, BY and DTM were positively correlated with GY under rainfed condition and could be reliable traits for drought tolerance and obtaining a high potential yielding in rainfed condition. Correlation analysis showed that STI, GMP and MP indices were positively correlated with grain yield under irrigated and rainfed conditions, NKS, KWS, BY, CHL, FLW, FLA and DTM traits. Therefore, they can be used as suitable criteria for identifying drought tolerance in genotypes.
REFERENCES
- Ahmad, R., S. Qadir, N. Ahmad and K.H. Shah, 2003. Yield potential and stability of nine wheat varieties under water stress conditions. Int. J. Agric. Biol., 5: 7-9.
Direct Link - Bahrami, F., A. Arzani and V. Karimi, 2014. Evaluation of yield-based drought tolerance indices for screening safflower genotypes. Agron. J., 106: 1219-1224.
CrossRefDirect Link - Bayoumi, T.Y., M.H. Eid and E.M. Metwali, 2008. Application of physiological and biochemical indices as a screening technique for drought tolerance in wheat genotypes. Afr. J. Biotechnol., 7: 2341-2352.
Direct Link - Blum, A., 2005. Drought resistance, water-use efficiency and yield potential-are they compatible, dissonant, or mutually exclusive? Aust. J. Agric. Res., 56: 1159-1168.
CrossRefDirect Link - Bouslama, M. and W.T. Schapaugh, 1984. Stress tolerance in soybeans. I. Evaluation of three screening techniques for heat and drought tolerance. Crop Sci., 24: 933-937.
CrossRefDirect Link - Calderini, D.F., M.P. Reynolds and G.A. Slafer, 2006. Source-sink effects on grain weight of bread wheat, durum wheat and triticale at different locations. Aust. J. Agric. Res., 57: 227-233.
CrossRefDirect Link - Clark, J.M., R.M. DePauw and T.F. Townley-Smith, 1992. Evaluation of methods for quantification of drought tolerance in wheat. Crop Sci., 32: 723-728.
CrossRefDirect Link - Dorostkar, S., A. Dadkhodaie and B. Heidari, 2015. Evaluation of grain yield indices in hexaploid wheat genotypes in response to drought stress. Arch. Agron. Soil Sci., 61: 397-413.
CrossRefDirect Link - Ehdaie, B. and J.G. Waines, 1993. Variation in water-use efficiency and its components in wheat: I. Well-watered pot experiment. Crop Sci., 33: 294-299.
CrossRefDirect Link - Fernandez-Aparicio, M., F. Flores and D. Rubiales, 2009. Field response of Lathyrus cicera germplasm to crenate broomrape (Orobanche crenata). Field Crops Res., 113: 321-327.
CrossRefDirect Link - Fischer, R.A. and R. Maurer, 1978. Drought resistance in spring wheat cultivars. I. Grain yield responses. Aust. J. Agric. Res., 29: 897-912.
CrossRefDirect Link - Garcia del Moral, L.F., Y. Rharrabti, S. Elhani, V. Martos and C. Royo, 2005. Yield formation in Mediterranean durum wheats under two contrasting water regimes based on path-coefficient analysis. Euphytica, 146: 203-212.
CrossRefDirect Link - Gavuzzi, P., F. Rizza, M. Palumbo, R.G. Campanile, G.L. Ricciardi and B. Borghi, 1997. Evaluation of field and laboratory predictors of drought and heat tolerance in winter cereals. Can. J. Plant Sci., 77: 523-531.
CrossRefDirect Link - Kilic, H. and T. Yagbasanlar, 2010. The effect of drought stress on grain yield, yield components and some quality traits of durum wheat (Triticum turgidum ssp. durum) cultivars. Notulae Botanicae Horti Agrobotanici Cluj-Napoca, 38: 164-170.
Direct Link - Kirigwi, F.M., M. van Ginkel, R. Trethowan, R.G. Sears, S. Rajaram and G.M. Paulsen, 2004. Evaluation of selection strategies for wheat adaptation across water regimes. Euphytica, 135: 361-371.
CrossRefDirect Link - Lewis, R.S., 2006. Identification of germplasm of possible value for confronting an unfavorable inverse genetic correlation in tobacco. Crop Sci., 46: 1764-1771.
CrossRefDirect Link - Lonbani, M. and A. Arzani, 2011. Morpho-physiological traits associated with terminal drought-stress tolerance in triticale and wheat. Agron. Res., 9: 315-329.
Direct Link - Mitra, J., 2001. Genetics and genetic improvement of drought resistance in crop plants. Curr. Sci., 80: 758-763.
Direct Link - Nouri, A., A. Etminan, J.A. Teixeira da Silva and R. Mohammadi, 2011. Assessment of yield, yield-related traits and drought tolerance of durum wheat genotypes (Triticum turjidum var. durum Desf.). Aust. J. Crop Sci., 5: 8-16.
Direct Link - Pinto, R.S., M.P. Reynolds, K.L. Mathews, C.L. McIntyre, J.J. Olivares-Villegas and S.C. Chapman, 2010. Heat and drought adaptive QTL in a wheat population designed to minimize confounding agronomic effects. Theoret. Applied Genet., 121: 1001-1021.
CrossRefDirect Link - Plaut, Z., B.J. Butow, C.S. Blumenthal and C.W. Wrigley, 2004. Transport of dry matter into developing wheat kernels and its contribution to grain yield under post-anthesis water deficit and elevated temperature. Field Crops Res., 86: 185-198.
CrossRefDirect Link - Rajaram, S., 2001. Prospects and promise of wheat breeding in the 21st century. Euphytica, 119: 3-15.
CrossRefDirect Link - Rosielle, A.A. and J. Hamblin, 1981. Theoretical aspects of selection for yield in stress and non-stress environment. Crop Sci., 21: 943-946.
CrossRefDirect Link - Rubio, J., J.I. Cubero, L.M. Martin, M.J. Suso and F. Flores, 2004. Biplot analysis of trait relations of white lupin in Spain. Euphytica, 135: 217-224.
CrossRefDirect Link - Sayar, R., H. Khemira, A. Kameli and M. Mosbahi, 2008. Physiological tests as predictive appreciation for drought tolerance in durum wheat (Triticum durum Desf.). Agron. Res., 6: 79-90.
Direct Link - Warrington, I.J., R.L. Dunstone and L.M. Green, 1977. Temperature effects at three development stages on the yield of the wheat ear. Aust. J. Agric. Res., 28: 11-27.
CrossRefDirect Link - Yan, W., 2001. GGEbiplot-A windows application for graphical analysis of multi environment trial data and other types of two-way data. Agron. J., 93: 1111-1118.
CrossRefDirect Link - Yan, W. and I. Rajcan, 2002. Biplot analysis of test sites and trait relations of soybean in Ontario. Crop Sci., 42: 11-20.
CrossRefDirect Link