
Leonardo Gomez
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Sigifredo Padilla
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Alejandro Fuentes
Agricultural Research Direction, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Yoslaine Ruiz
Agricultural Research Direction, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Tatiana Gonzalez
Department of Process Control, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Margarita Somoza
Department of Process Control, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Lisette Lopez
Department of Process Control, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Julio Sanchez
Department of Process Control, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
David Gavilan
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Elio Espinosa
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Yenisleydis Avila
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Otto Mendoza
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Yordanka Masforrol
Department of Chemical Synthesis, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Cristina Garcia
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
O. Maylin La
Quality Control Direction, Center for Genetic Engineering and Biotechnology, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba
Rodolfo Valdes
Department of Monoclonal Antibody Production, Ave 31/158 and 190, P.O. Box 6162, Havana 10600, Cuba







ABSTRACT
Transgenic plants are an attractive alternative to produce antibodies for the manufacturing of biologics. The main subject of this study was to assess two non-commercial transgenic tobacco plant varieties (BHmN and Habana 92) cultivated in zeolite and confined conditions for the production of a hepatitis B virus surface antigen (HBsAg)-specific plantibody (PHB-01). The plantibody molecule biochemical characterization and the assessment of the immunopurification capacity of the antigen recognized by PHB-01-immunoadsorbents were other subjects studied in this study to decide which variety would be more suitable for PHB-01 large-scale production. As results, the BHmN variety allowed obtaining 1.18 fold more biomass of leaves and up to 2 fold in the pantibody yield with similar specificity and affinity constant by the HBsAg. The assessment of the HBsAg immunopurification capacity of the PHB-01 immunoadsorbents produced by these two non-commercial transgenic plant varieties did not showed significant differences in terms of adsorption capacity (p = 0.2135), elution capacity (p = 0.1239), recovery (p = 0.2655) and purity during 13 purification cycles. In conclusion, the BHmN variety would be the most suitable variety for the production of the PHB-01.
PDF Abstract XML References Citation
Received: January 22, 2013;
Accepted: April 01, 2013;
Published: June 03, 2013
How to cite this article
Leonardo Gomez, Sigifredo Padilla, Alejandro Fuentes, Yoslaine Ruiz, Tatiana Gonzalez, Margarita Somoza, Lisette Lopez, Julio Sanchez, David Gavilan, Elio Espinosa, Yenisleydis Avila, Otto Mendoza, Yordanka Masforrol, Cristina Garcia, O. Maylin La and Rodolfo Valdes, 2013. Assessment of Two Transgenic Tobacco Plant Varieties for the HBsAg-Specific Plantibody Production. Journal of Agronomy, 12: 11-19.
DOI: 10.3923/ja.2013.11.19
URL: https://scialert.net/abstract/?doi=ja.2013.11.19
DOI: 10.3923/ja.2013.11.19
URL: https://scialert.net/abstract/?doi=ja.2013.11.19
INTRODUCTION
Several procedures have been developed to produce small and large quantities of monoclonal antibodies (Kohler and Milstein, 1975). The history of these procedures has evolved from ascitic fluid method, to transgenic plants (Knazek et al., 1972; Brodeur et al., 1984; Hiatt et al., 1989, Veliz, 2002; Wei et al., 2006; Loos et al., 2011; Srivastava et al., 2011). Regarding to this, the capacity of plant cells to express and correctly process complex proteins such as antibodies, the unlimited biomass that can be achieved, the absence of cross-contaminating adventitious agents and the potentially simple downstream process are seem as advantages of this production alternative (De Jaeger et al., 1999; Stoger et al., 2002; Hood et al., 2002; Arntzen et al., 2005; Malabadi and Nataraja, 2007; Behrooz et al., 2008; Gomez et al., 2010).
For these reasons, the Center for Genetic Engineering and Biotechnology of Havana, Cuba developed an industrial-scale process to produce a plantibody (PHB-01) specific for the hepatitis B virus surface antigen (HBsAg) by transgenic plants. This HBsAg specific plantibody was expressed in tobacco plants to be isolated and employed as immunoreagent (matrix’s ligand) to immunopurify the HBsAg, which is used as the active pharmaceutical ingredient (HBsAg) of the Cuban vaccine against Hepatitis B (Ramirez et al., 2003; Valdes et al., 2003a; Pujol et al., 2005; Padilla et al., 2009).
However, the plantibody yield obtained in the Habana 92 variety has been no enough so far (Ramirez et al., 2003), aspects that seems to be a common issue of this technology (Franklin and Mayfield, 2005; Stoger et al., 2005, 2000; Decker and Reski, 2008; Sainsbury and Lomonossoff, 2008; Vezina et al., 2009; Hensel, 2011; De Muynck et al., 2010).
Therefore, the main subject of this study was to assess two non-commercial transgenic tobacco plant varieties (BhmN and Habana 92) cultivated in zeolite and confined conditions for the production of a immunopurification capacity of the antigen recognized by PHB-01-immunoadsorbents were other subjects studied in this study to decide which variety would be more suitable for PHB-01 large-scale production.
MATERIALS AND METHODS
Transgenic plants: Transgenic tobacco plants expressing the anti-HBsAg CB.Hep-1 monoclonal antibody (mAb) PHB-01 BHmN (clone 26) and PHB-01 H-92 (clone 23) (Ramirez et al., 2003) were used in this study to compare the plantibody yield in BHmN and Habana 92 varieties, respectively. Briefly, a tandem expression vector strategy to produce these molecules in these Nicotiana tabacum varieties was employed. Clones 26 and 23 were chosen among several clones for this study.
Chemical and mineralogical compositions of zeolite used in this study: The chemical composition of the zeolite employed as substrate to cultivate both variety plants was (% in weight): SiO2 (66.62), Al2O3 (12.17), Fe2O3 (2.08), CaO (3.19), MgO (0.77), Na2O (1.53), K2O (1.20), PPI (11.02), for a total of 98.58. The ratio of SiO2/Al2O3 (mol) was 9.29. The total cationic-exchange capacity expressed as TCEC meq/100 g was 138.69 and for each element was Ca2+ 94.48, Mg2+ 4.13, Na2+ 32.49 and for K+ 7.59. Its mineralogical composition was Clinoptilolite (49%), Mordenite (12%), Montmorillonite (Na, Ca) 0.33 (Al, Mg) 2 Si4O10 (OH) 2nH2O (very few); Calcite, CaCO3 (very few); Quarz, SiO2 (very few); Feldespast, Fe-aluminosilicates (undetermined). The hardness was 1.08±2.1 and results of chemical resistance (oxidability) was NaCl (3.13±1.9%), HCl (3.39±0.5%), NaOH (2.50±1.3%), H2O (2.38±1.2%), Cl2 (1.58±2.19%). The characteristics of main granular materials were (Mean±CV): Particle density (2120.0±1.45 kg cm¯3), apparent density (1021.0±1.76 kg cm-3), porosity (0.51±0.4) and shape factor (0.64±1.97) (Marquez et al., 2000).
Generation of seedlings: Nicotiana tabacum L., seeds were cultivated in Turba Rubia (85% organic matter, 15% ashes, 0,26% total nitrogen, 300 mg L-1 phosphorous anhydride, 144 mg L-1 magnesium oxide, pH 5.5-6, 50% maximum humidity, TCEC 100-130 meq/100 g and 500-700 μS cm-1 conductivity) using the floating tray method (Cristanini, 1995). Seedlings were fertilized with 5 g L-1 of the Haifa-Chemical, Israel 19-19-19 NPK-fertilizer solution at days 7th and 21st, respectively and pruned at 28th day after seedtime. Next, the pruning was repeated every 48 h up to the seedling transplant into the green-containment for producing vegetal biomass rich in PHB-01 BHmN and PHB-01 H-92, respectively.
Production of transgenic plants: Both varieties of plants were growth in 5 furrows at 8 plants by square meter in a green house under controlled conditions using the zeolite employed in the seedling generation. Briefly, plants were fertilized, dropping four times per day during 4 min, with the 50 g L-1 of the 19-19-19 NPK-fertilizer solution (Haifa-Chemical, Israel). The vol-vol Magnisal and 19-19-19 NPK-fertilizer solution (Haifa-Chemical, Israel) were used after of 4th week. After 9 weeks, plants were then harvested. The estimation of the biomass production and weight of leaves, roots and stems were done using all cultivated plants. (Pushkina et al., 1996).
Purification of plantibody from transgenic tobacco plants: Nicotiana tabacum plants were processing using a Fitzmill model D6A Comminutor (Fitzpatrick Company, USA). Green fraction was then removed from the juice by centrifugation at 1,500 rpm in a Rina 100F 800 bascket centrifuge (Riera Nadeu S.A, Spain) and supernatants, still containing small amount of suspended green particles, were heated. Next, heated supernatants were centrifuged in a Wetsfalia centrifuge and submitted to an acid precipitation with 100 mM HCl. After that, the precipitated supernatants were equilibrated with 2M Tris and centrifuged again to be applied in a chromatography column loaded with ProSepvA Ultra. Elution fractions were submitted to saline precipitation using Ammonium sulfate. This material was centrifuged again and pellets were dissolved in 20 mM Tris/150 mM NaCl, pH 7.0. Subsequently, a gel filtration chromatography using Sephadex G-25 coarse (Amersham-Biosciences, Uppsala, Sweden) was performed for desalting the material before the plantibody preparation sterile filtration.
Estimation of plantibody concentration by enzyme-linked immunosorbent assay: A polystyrene microplate (Costar, Cambridge, USA) was coated with 10 μg per well of HBsAg in 100 mM NaHCO3 buffer for 20 min at 5°C. After this step, samples were added to the plate in 150 mM saline buffered solution (PBS)/0.05% Tween 20 and incubated for 1 h at 37°C. Several washings were done with PBS/0.05% Tween 20. The plate was incubated for 1 h at 37°C with an IgG-horseradish peroxidase conjugate (Sigma Chemical Co., St. Louis, USA). The reaction was then revealed using 100 μL well-1 of 0.05% O-phenylendiamine (OPD) and 0.015% H2O2 in citrate buffer (pH 5.0) and stopped with 50 μL well-1 of 1.25 M H2SO4. The absorbance was measured in a Multiskan ELISA reader (Labsystems, Helsinki, Finland) using a 492 nm filter (Leyva et al., 2007).
Protein determination: Protein concentration was determined following the method described by (Lowry et al., 1951), using bovine serum albumin as standard material. The curve range was from 10 to 100 μg mL-1 and the absorbance of samples was measured at 280 nm in a UV/Visible Ultrospec 2000 Spectrophotometer (Pharmacia Biotech, Cambridge, England).
SDS-PAGE and western-blott: The purity of the plantibody samples was analyzed carrying out an electrophoresis on a 12.5% (w/v) SDS-polyacrylamide gel (Laemmli, 1970) followed by a Coomassie blue staining.
Determination of plantibody affinity constant: The affinity constant of plantibody molecules was determined by the method described by Beatty et al. (1987). Microtiter plates were coated with the HBsAg and incubated with plantibodies. Plates were sequentially incubated with a horseradish peroxidase-antibody conjugate and the reaction was revealed using OPD as substrate and 0.015% H2O2 in citrate buffer; pH 5.0. Reaction was stopped by adding 50 mL of 2M H2SO4. The plantibody amount adherent to the HBsAg was reflected by the enzyme product measured by optical density at 492 nm using an ELISA reader (Labsystem, Helsinki, Finland).
Specificity assay: The peptide recognition assay was performed using 40 mg of peptides extracts Tenta Gel resin humid. This resin was washed with purified water (3 times x 5 min) and washed again with 150 mM PBS (3 times x 5 min). Then, it was blocked with BSA 1% in 150 mM PBS for 1h. After that, the resin was incubated with respective PHB-01 molecules in 400 μL (1 mg mL-1) in BSA's dissolution to the 1% in 150 mM PBS (1 h). Next, a wash with 0.05% Tween 20/150 mM PBS (5 times x 5 min) was done. Incubation with 400 μL with an anti mouse IgG-phosphatase alkaline was performed (1:2000) for 1h, followed by washings with 0.05% Tween 20/150 mM (5 times x 5 min). The resin was then incubated with BCIP solution (5-bromo-4-cloro-3-indolil-fosfato) in 400 μL of substrate solution for 30 min (until color appeared). Reaction was stopped when pearls colored were washed with 150 mM PBS (5x5 min). The 37 peptide sequence was TTSTGPCKTCTT. This corresponds with the peptide 20 of “a” determinant of the HBsAg and is a positive control. Besides, RFSWLSLLVPFV (P36) corresponds with the peptide 29 of HBsAg but no with a determinant and is a negative control. On the other hand, the mAb anti CEA (anti-carcinogenic embrionary antigen) was used as negative control of antibody.
Purification of the CB.Hep-1 mAb used as experimental control: Hybridoma 48/1/5/4 was grown as ascites tumor in specific pathogen free Balb/c mice. Animals received 0.5 mL of mineral oil and 1x106 cells per animal were inoculated into the animal peritoneal cavity in cell culture medium RPMI (Gibco, Grand Island, USA). Abdominal taps were performed when marked abdominal distention was observed. The purification process included clarification and filtration of the ascites, affinity chromatography, using n-protein A SepharoseTM 4 fast flow loaded in a column BPG100/500 operated at 100 cm h-1. Then, a gel-filtration chromatography, using Sephadex G-25 coarse (Amersham-Biosciences, Uppsala, Sweden), was performed in a BPG200/750 column at 130 cm h-1 to exchange the buffer for allowing the antibody concentration in a Sartocon alpha (SARTOCON Silice Cassettte) and the sterile filtration using capsules of 0.45-0.2 μm SARTOBRAN P Sterile MidiCap of Sartorius Stedim Biotech, Germany.
Hepatitis B surface antigen production: The HBsAg was produced by fermentation of a recombinant strain of Pichia pastoris (C-226) in saline medium supplemented with glycerol and its expression was induced by methanol. Cells were centrifuged and disrupted with 20 mM Tris-HCl (pH 8.0)/3 mM EDTA/300 mM NaCl/3M KSCN/10 g L-1 sucrose on a bed mill (KDL type: WAB, Basel, Switzerland). The homogenate was precipitated by adding 1M HCl and centrifuged at 10000 xg for 30 min. Supernatant was adsorbed on Hyflo Super Cell (a flux-calcined grade of Celite filter-aid) and equilibrate to pH 4.0 under continuous stirring during 2 h. After washing step, HBsAg was eluted with 20 mM Tris/1M HCl/3 mM EDTA/100 g L-1 sucrose (pH 8.2) and semipurified material of 10-25% purity was used as starting material for the immunoaffinity chromatography experiments (Lucila et al., 1992).
Immunoaffinity matrix: Sepharose CL-4B (Amersham-Bioscences, Uppsala, Sweden) was activated by the cianogen bromide (CNBr) method (Kohn and Wilchek, 1984). The CB.Hep-1 mAb and plantibodies were coupled as recommended by Amersham-Biosciences, Uppsala, Sweden (Gomez et al., 2002) (Wimalasena and Wilson, 1991). The amount of coupled antibody was determined by measuring the total protein before and after the coupling reaction (about 4 mg mL-1 of gel for each immunogel).
Immunoaffinity chromatography: Gels were packed into PD-10 chromatography columns with 7 mL of gel. (Amersham-Bioscences, Uppsala, Sweden) and equilibrated with 20 mM Tris-HCl/3 mM EDTA, pH 7.8. Adsorption and elution flow rates were 20 and 35 cm h-1, respectively. Columns were loaded with a partially purified HBsAg preparation in the equilibrium buffer containing 1M NaCl. After washing, the bound antigen was eluted with 20 mM Tris/3M KSCN/3 mM EDTA, pH 7.0 and monitored at 280 nm.
Mathematical analysis: ANOVA tests were carried out using the Statgraphic program (version 5.0) and p-values less than 0.05 were considered to be of statistical significance.
RESULTS AND DISCUSSION
Since Hiatt et al. (1989), pioneered the expression of immunoglobulin chains in tobacco plants, various portions of immunoglobulin chains have been expressed heterologously, including single chain molecules, Fab fragments, small immune proteins, IgGs and chimeric secretory IgAs (Hensel, 2011). In view of the growing demand for recombinant antibodies, production capacity has been saturated, so there is a need for the development of attractive alternative means of production to the classical mammalian expression systems such as transgenic plants (Morrison et al., 1984; Jones et al., 1986; Jain et al., 2007; De Muynck et al., 2010).
In regard to the production of antibodies in transgenic plants, different levels of plantibody yield have been obtained in different host species and organs. However, the huge majority of examples showed an insufficient level of expression. For instances, the maximum expression level obtained in leaves has been 325 μg g-1 fresh weight (FW) of Nicotiana bethamiana, 757 μg g-1 FW in Nicotiana tabacum (Sainsbury and Lomonossoff, 2008; Vezina et al., 2009), 1.1 μg g-1 FW (H10 antibody) in Nicotiana tabacum (Villani et al., 2009), 0.9 μg g-1 FW (CO17-1A) in Nicotiana Bethamiana (Ko et al., 2005), 1 μg g-1 FW (cT84.66) in Nicotiana tabacum (Vaquero et al., 1999), 8.5 μg g-1 FW (rAb29) in Nicotiana tabaccum (Schillberg et al., 1999), 60 μg g-1 FW (MGR48) in Nicotiana tabacum (Stevens et al., 2000) and 31 μg g-1 FW (BR55-2) in Nicotiana tabacum (Brodzik et al., 2006). Similar situation was described by (Ramirez et al., 2003) for the expression of the PHB-01 in Nicotiana tabacum. That is why; authors sought to assess the yield of the PHB-01 in two different tobacco plant varieties (BHmN and Habana 92), to decide the suitable condition for large-scale production.
The comparison of these two varieties of transgenic tobacco plant cultivated in zeolite under confined conditions in terms of biomass yield and weight of leaves, stems and plants demonstrated that the biomass yield (kg of leaves) measured in the BHmN variety was 1.18 fold higher than in Habana 92 variety (p = 0.0023).
 | |
| Fig. 1: | Picture of the BHmN (left) and Habana 92 (right) varieties cultivated in zeolite under confined conditions |
Similar increase was observed when the weight of stems (1.14 fold, p = 0.0470) was measured. In consequence, the comparison of the whole weight of the plants evidenced better results for the BHmN variety (304.02 g) against Habana 92 (258.65 g). The increase in this parameter was 1.17 fold (p = 0.0089) (Table 1). Other agronomic values estimated at 7 weeks of cultivation were (BHmN:) foliar area 1666.1 cm2, height growth rate 0.085 cm day-1 and average height 80.6 cm. (Habana 92:) foliar area 1174.2 cm2, height growth rate 0.028 cm day-1 and average height 72.4 cm. In addition, BHmN plants were smaller than Habana 92 plants but had more and bigger leaves than Habana 92 plants (Fig. 1).
The plantibody expression level measured was 0.28% for BHmN and 0.11% for Habana 92, which represented 2.54 fold of increase. In this regards, authors must point out that Total Soluble Protein (TSP) concentration was 10.98 and 13.07 mg mL-1 in the leaf extract of BHmN and Habana 92 varieties, respectively, which was coincident with those values reporter by (Floss et al., 2008). As consequence, results of the plantibody yield in the extract of leaves was 67.62 mg kg-1 for BHmN and 33.67 mg kg-1 for Habana 92, representing thus a net increase of 2.00 fold (Table 1), which was analogous with those reported by (Stevens et al., 2000) and controversial with results reported by (Vaquero et al., 1999; Villani et al., 2009).
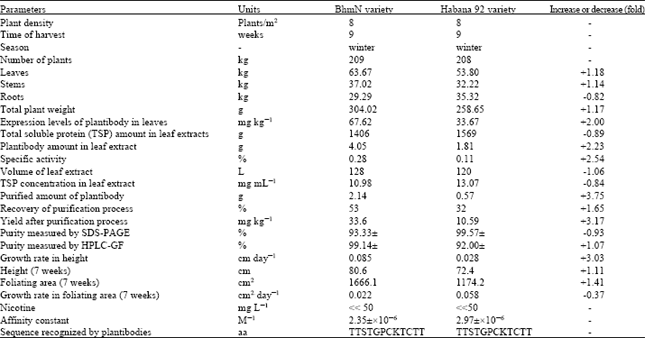
In respect to results obtained after the purification of the plantibody molecules, the yield of plantibodies after the purification process, expressed as mg of plantibody per kg of processed leaves, was 33.6 (BHmN) and 10.59 (Habana 92). That corresponded to a 3.17 fold increase (Table 1).
| Table 1: | Results of the harvests of transgenic plants and purification of the plantibody molecules |
 | |
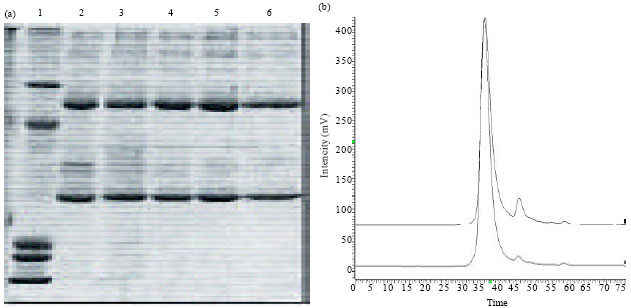 | |
| Fig. 2(a-b): | Purity of the plantibody molecules measured by SDS-PAGE under reduction conditions (a) and HPLC-GF (b). (a). Line 1: Molecular Weight Marker-091201: Bovine serum albumin 60 kDa, Ovoalbumin 41.07 kDa, Trypsin inhibitor 20.6 kDa, Lisozime 12 kDa, Aprotinin 6,4kDa. Line 2: PHB-01. Line 3: PHB-01 BHmN (experiment 1). Line 4: PHB-01 BHmN (experiment 2). Line 5: PHB-01 BHmN (experiment 3). Line 6: Reference Material-020907 (CB.Hep-1 monoclonal antibody used as experimental control). (B). a) PHB-01 BHmN, b) PHB-01 H-92 |
The purity of the purified molecules estimated by SDS-PAGE under reducing conditions was 93% for BhmN and 92% for Habana 92, whereas 99.57 and 99.14% were values of purity measured by gel-filtration-HPLC in both varieties, respectively (Fig. 2). Therefore, no differences were observed in this important parameter between both varieties. In that sense, similar results were obtained by Valdes et al. (2003b).
Another remarkable issue that has to be considered is the nicotine level in the purified plantibody preparation.
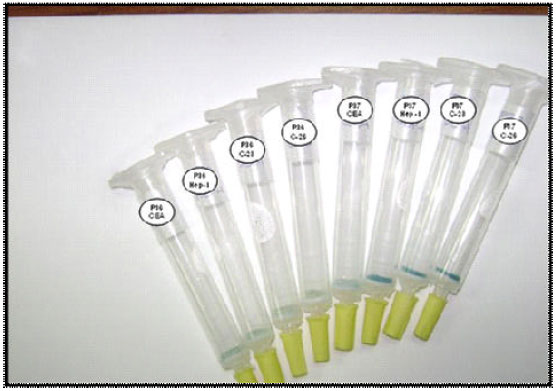 | |
| Fig. 3: | Results of the assay performed to demonstrate the specificity of the plantibody molecules by the HB |
This is an important aspect for human health because after the nicotine absorption, Adrenals glands activation is produced and a discharge of adrenaline produces corporal stimulation and sudden discharge of glucose, blood pressure increase, respiration and heart rate. The addictive potential is due to the dopamine secretion produced on the regions of the brain that control placer sensations. The nicotine decreases basal metabolism. The interaction with other substances including drugs is possible using the same metabolic way of C-P450 in liver. In the brain, part of the metabolites is transformed in intermediates (as nornicotine) that could be neurotoxic, which acts on the nicotine cholinergic receptors of the central nervous system (Andersson et al., 2003). In such sense, the nicotine values measured in both purified plantibody preparations were always lower than the detection limit of the technique (0.05 mg L-1) and lower than 50 mg. This is of the minimum toxic doses of nicotine permitted in humans (Andersson et al., 2003; Tuduri, 2010).
Results of assay designed to detect the epitope recognized by plantibody molecules corroborated that both molecules recognized the same epitope into the “a” determinant of the HBsAg molecule. This can be appreciated with the blue color displayed in samples of PHB-01 BHmN, PHB-01 H-92 and CB.Hep-1 mAb corresponding with the peptide 37 case. The absence of color in the anti-CEA mAb sample in the peptide 37 validated the specificity of the assay. Results of the assays with the peptide 36 revealed that PHB-01 BHmN, PHB-01 H-92 and CB.Hep-1 mAb did not recognized an aminoacid sequence non-related with the “a” determinant of the HBsAg (Fig. 3).
Table 1 shows the affinity constant of plantibody molecules purified from both varieties. The affinity constant values obtained were 2.35x10-8 and 2.97x10-8, which no also corroborate the specificity of the molecules but the strength of association with HBsAg.
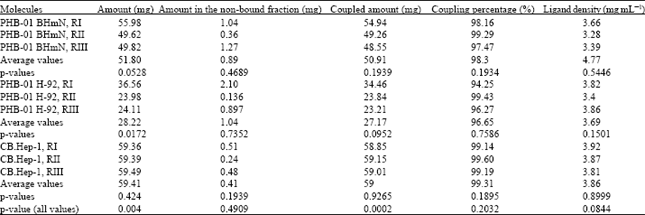
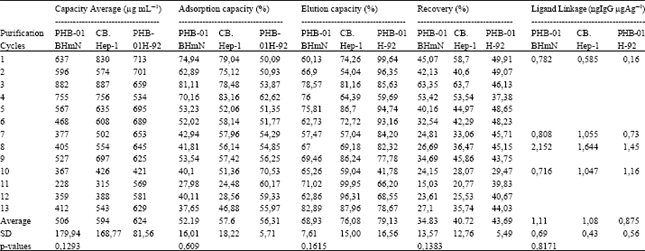
Finally, immunoadsorbents of both plantibodies were performed at a ligand density ranged 3-5 mg mL-1 of gel, because we have previously demonstrated that there were not differences in the behavior of the CB.Hep-1 mAb immunoadsorbents during the HBsAg purification in this range of ligand density (Gomez et al., 2002). As Table 2 shows, there were not statistical differences in the plantibody coupling efficiency (p = 0.2032), immunoadsorbent ligand density (p = 0.0844), HBsAg adsorption capacity (p = 0.609), HBsAg elution capacity (p = 0.1615) and HBsAg recovery (p = 0.1383). In addition, the ligand linkage determined in the purification cycles (1, 7, 8 and 10) was always below the limit established for this parameters (3 ng IgG mg-1 HBsAg) and without significant differences (p = 0.8171) between them (Table 3).
| Table 2: | Results of the comparison of the plantibody inmovilization experiments |
 | |
| Table 3: | Results of the evaluation of the HBsAg immunopurification experiments |
 | |
Similar results were obtained when results of the plantibody-immunoadsorbents where compared with those obtained with the immunoadsorbent prepared with the CB.Hep-1 mAb used as positive control.
CONCLUSION
The BHmN variety allowed obtaining 1.18 fold more biomass of leaves that Habana 92 variety. The plantibody amount measured in the leaf extracts of the BHmN variety plants was 2.54 fold higher than in the Habana 92 variety. The total soluble concentration was similar between both varieties. The plantibody yield measured in the leaf extracts of the BHmN variety plants was 2.0 fold higher than in the Habana 92 variety. The recovery of the plantibody process purification was 1.65 fold higher in BHmN experiments than in Habana 92. The plantibody yield measured after the purification process was 3.17 fold higher in BHmN variety than in the Habana 92 variety experiments. The purity of the plantibody molecules measured by SDS-PAGE and HPLC-GF was always higher than 90% and statistically similar between molecules purified from both varieties. Both plantibody molecules recognized the same epitope on the HBsAg and with similar affinity constant. The behaviour of immunoadsorbents of both plantibody molecules was statistical equal in terms of plantibody coupling efficiency, plantibody ligand density, HBsAg adsorption capacity, HBsAg elution capacity, HBsAg recovery and plantibody leakage. Therefore, the replacement of the Habana 92 variety by BHmN variety for the production of the HBsAg-specific plantibody for vaccine purpose has been scientifically argued.
ACKNOWLEDGMENTS
This study was supported by the Center for Genetic Engeneering and Biotechnology of Havana and authors gratefully acknowledge to: Danay Callard of the Direction of Agropecuary Research for his assistant on the obtaining of transgenic BHmN variety and Yordan Issac, Yarusenky Lescaille, Adelma Perez and Yandiesky Lowery for his assistant in the cultivation of two N. tabacum varieties.
REFERENCES
- Loos, A., B. Van Droogenbroeck, S. Hillmer, J. Grass and R. Kunert et al., 2011. Production of monoclonal antibodies with a controlled N-glycosylation pattern in seeds of Arabidopsis thaliana. Plant Biotechnol. J., 9: 179-192.
CrossRef - Arntzen, C., S. Plotkin and B. Dodet, 2005. Plant-derived vaccines and antibodies: Potential and limitations. Vaccine, 23: 1753-1756.
PubMed - Behrooz, D., S. Farajnia, M. Toorchi, S. Zakerbostanabad, S. Noeparvar and C.N. Stewart Jr., 2008. DNA-delivery methods to produce transgenic plants. Biotechnology, 7: 385-402.
CrossRefDirect Link - Beatty, D.J., G.B. Beatty and W.G. Vlahos, 1987. Measurement of monoclonal antibody affinity by non-competitive enzyme immunoassay. J. Immunol. Methods, 100: 173-179.
PubMed - Brodeur, B.R., P. Tsang and Y. Larose, 1984. Parameters affecting ascites tumour formation in mice and monoclonal antibody production. J. Immunol. Methods, 71: 265-272.
CrossRef - Brodzik, R., M. Glogowska, K. Bandurska, M. Okulicz and D. Deka et al., 2006. Plant-derived anti-Lewis Y mAb exhibits biological activities for efficient immunotherapy against human cancer cells. Proc. Natl. Acad. Sci. USA., 103: 8804-8809.
CrossRefDirect Link - De Muynck, B., C. Navarre and M. Boutry, 2010. Production of antibodies in plants: Status after twenty years. Plant Biotechnol. J., 8: 529-563.
CrossRefDirect Link - Decker, E.L. and R. Reski, 2008. Current achievements in the production of complex biopharmaceuticals with moss bioreactors. Bioprocess Biosyst. Eng., 31: 3-9.
CrossRefPubMedDirect Link - Floss, D.M., M. Sack, J. Stadlmann, T. Rademacher and J. Scheller et al., 2008. Biochemical and functional characterization of anti-HIV antibody-ELP fusion proteins from transgenic plants. Plant Biotechnol. J., 6: 379-391.
CrossRefPubMedDirect Link - Franklin, S.E. and S.P. Mayfield, 2005. Recent developments in the production of human therapeutic proteins in eukaryotic algae. Expert Opin. Biol. Ther., 5: 225-235.
CrossRefPubMedDirect Link - Gomez, L., R. Valdes, M. Pujol, S. Padilla and T. Gonzalez et al., 2010. Kinetic of expression of a plantibody in (Nicotiana tabacum) plants cultivated in different substrates (Zeolite and Soil). J. Agron., 10: 20-26.
CrossRef - Hiatt, A., R. Cafferkey and K. Bowdish, 1989. Production of antibodies in transgenic plants. Nature, 342: 76-78.
CrossRef - Jain, M., N. Kamal and S.K. Batra, 2007. Engineering antibodies for clinical applications. Trends Biotechnol., 25: 307-316.
CrossRefPubMedDirect Link - Knazek, R.A., P.M. Gullino, P.O. Kohler and R.L. Dedrick, 1972. Cell culture on artificial capillaries: An approach to tissue in vitro. Science, 178: 65-66.
CrossRefPubMedDirect Link - Kohler, G. and C. Milstein, 1975. Continuous cultures of fused cells secreting antibody of predefined specificity. Nature, 256: 495-497.
CrossRef - Laemmli, U.K., 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227: 680-685.
CrossRefDirect Link - Leyva, A., A. Franco, T. Gonzalez, J.C. Sanchez and I. Lopez et al., 2007. A rapid and sensitive ELISA to quantify an HBsAg specific monoclonal antibody and a plant-derived antibody during their downstream purification process. Biologicals, 35: 19-25.
PubMed - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Malabadi, R. B. and K. Nataraja, 2007. Stable transformation and recovery of transgenic plants by particle bombardment in Pinus wallichiana A.B. Jacks (Himalayan blue pine). Biotechnology, 6: 105-111.
CrossRefDirect Link - Padilla, S., R. Valdes, L. Gomez, D. Geada and W. Ferro et al., 2009. Assessment of a plantibody HB-01 purification strategy at different scales. Chromatographia, 70: 1673-1678.
CrossRefDirect Link - Pujol, M., N.I. Ramirez, M. Ayala, J.V. Gavilondo and R. Valdes et al., 2005. An integral approach towards a practical application for a plant-made monoclonal antibody in vaccine purification. Vaccine, 23: 1833-1837.
PubMed - Pushkina, G.P., P.M. Lyan, L.M. Bushkovskaya and L.I. Krymova, 1996. The use of zeolites for growing medicinal plants. Pharmaceut. Chem. J., 30: 574-578.
CrossRef - Ramirez, N., M. Rodriguez, M. Ayala, J. Cremata and M. Perez et al., 2003. Expression and characterization of an anti-(hepatitis B surface antigen) glycosylated mouse antibody in transgenic tobacco (Nicotiana tabacum) plants and its use in the immunopurification of its target antigen. Biotechnol. Applied Biochem., 38: 223-230.
CrossRefPubMed - Sainsbury, F. and G.P. Lomonossoff, 2008. Extremely high-level and rapid transient protein production in plants without the use of viral replication. Plant Physiol., 148: 1212-1218.
PubMedDirect Link - Schillberg, S., S. Zimmermann, A. Voss and R. Fischer, 1999. Apoplastic and cytosolic expression of full-size antibodies and antibody fragments in Nicotiana tabacum. Transgenic Res., 8: 255-263.
PubMedDirect Link - Morrison, S.L., M.J. Johnson, L.A. Herzenberg and V.T. Oi, 1984. Chimeric human antibody molecules: Mouse antigen-binding domains with human constant region domains. Inmunology, 81: 6851-6855.
PubMedDirect Link - Srivastava, N., V.K. Gupta, R. Pati and R.K. Gaur, 2011. Genetically by modified crops: An overview. Biotechnology, 10: 136-148.
CrossRefDirect Link - Stevens, L.H., G.M. Stoopern, I.J.W. Elbers, J.W. Molthoff and H.A.C. Bakker et al., 2000. Effect of climate conditions and plant developmental stage on the stability of antibodies expressed in transgenic tobacco. Plant Physiol., 124: 173-182.
CrossRefDirect Link - Stoger, E., J.K.C. Ma, R. Fischer and P. Christou, 2005. Sowing the seeds of success: Pharmaceutical proteins from plants. Curr. Opin. Biotechnol., 16: 167-173.
CrossRefDirect Link - Stoger, E., M. Sack, R. Fisher and P. Cristou, 2002. Plantibodies: Aplications advantages and boottlenecks. Curr. Opin. Biotechnol., 13: 161-166.
CrossRefPubMedDirect Link - Veliz, R.V., 2002. Alternative techniques to obtain monoclonal antibodies at small scale: Current state and future goals. Biotechnol. Appli., 19: 119-131.
Direct Link - Vaquero, C., M. Sack, J. Chandler, J. Drossard and F. Schuster et al., 1999. Transient expression of a tumor-specific single-chain fragment and a chimeric antibody in tobacco leaves. Proc. Natl. Acad. Sci. USA., 96: 11128-11133.
CrossRefDirect Link - Vezina, L.P., L. Faye, P. Lerouge, M.A. D'Aoust and E. Marquet-Blouin et al., 2009. Transient co-expression for fast and high-yield production of antibodies with human-like N-glycans in plants. Plant Biotechnol. J., 7: 442-455.
CrossRefPubMedDirect Link - Villani, M.E., B. Morgun, P. Brunetti, C. Marusic and R. Lombardi et al., 2009. Plant pharming of a full-sized tumour-targeting antibody using different expression strategies. Plant Biotechnol. J., 7: 59-72.
CrossRefPubMedDirect Link - Wei, S., Z. Sun, L. Han and L. Yu, 2006. Transgenic tobacco plants over-expressing arabidopsis transcriptional factor CBF1 show morphological and biochemical characteristics associated with cold tolerance. Asian J. Plant Sci., 5: 932-939.
CrossRefDirect Link - Wimalasena, R.L. and G.S. Wilson, 1991. Factors affecting the specific activity of immobilized antibodies and their biologically active fragments. J. Chromatogr., 572: 85-102.
CrossRef