
Majid Amini Dehaghi
Department of Agronomy, Faculty of Agriculture, Shahed University, Iran
Shahram Jafari
Department of Computer, Faculty of Engineering, Shiraz University, Shiraz Iran
Reza Tabatabaie
Department of Agronomy, Faculty of Agriculture, Shahed University, Iran
Journal of Agronomy
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 554-559







ABSTRACT
In order to examine the effect of calcium, salt and cold resistances during seed germination stage of Medicago polymorpha, an experiment was conducted in the Laboratory of the Agriculture College at Tarbiat Modarres University. The seeds were sterilized and incubated in 5, 10, 15 and 20 mM CaCl2 at 20°C for 24 h. Exogenous calcium treated seeds were transferred to 0, 50, 100, 150 and 200 mM NaCl combined with air temperature incubation at 5 and 15°C in germination chamber for 14 days. Then the seed germination percentage, number of normal and abnormal seedlings, fresh and dry weight and length of root and shoot and their ratios were determined. Results of the analysis of variance showed that the treatment and their interaction significantly influenced all measured parameters. The results indicated that calcium has a significant effect on the reduction the effect of stress condition during seed germination.
PDF Abstract XML References
How to cite this article
Majid Amini Dehaghi, Shahram Jafari and Reza Tabatabaie, 2007. Effect of Exogenous Calcium Chloride on Sodium Chloride Salt and Cold Resistances During Seed Germination Stage of Medicago polymorpha. Journal of Agronomy, 6: 554-559.
DOI: 10.3923/ja.2007.554.559
URL: https://scialert.net/abstract/?doi=ja.2007.554.559
DOI: 10.3923/ja.2007.554.559
URL: https://scialert.net/abstract/?doi=ja.2007.554.559
INTRODUCTION
The expansion of salty soils in Iran is approximated about 24 million hectares which is about 15% of the whole lands of the country. Although in some areas of Iran, there has been remarkable amount of sulfates, but dominant anion is chloride and dominant cation is Na+. Therefore, the saltiness of the lands in Iran is due to NaCl or Na2SO+4. Total salt in the kinds of land is about 3% (Jafari, 1995; Szabolcs, 1992).
The effect of NaCl salt on medicago germination and embryo developments in term of germination percentage, length of root and shoot and dry weight of embryo is intensively studied. The results indicated that all parameters are significantly decreased relating to NaCl salt treatment (Assadia and Miyamoto, 1987).
Aminpour and Aghaee (1997) after studying the salt effect on germination stage of 3 medicago varieties represented that with the increase of salt, the speed and the percentage of germination and length of root decreases and the length of root decreases more than that of shoot under the salt effect.
Saffari (1997) conducted a study of the salt effect on seeds germination of varieties of medicago species in different levels of salt from 0 to 300 mM. He informed that in 7th day and at distilled water, the average of seed germination was 80%, but in salt solution the percentage of germination decreased, especially when the salt concentration reached to more than 200 mM L-1 (ECe >15 m mhos cm-1 ).
Asgarian (1997), assessed the salt effect on germination of 5 varieties of medicago and indicated that increasing of salt causes an increase of the percentage of abnormal seedlings of the species at different ratios. Totally, the recorded results show that the effect of temperature reduction on physiologic activities of plants was conducted through the three following ways; effect on cell membrane, effect on intra-cell water, effect on enzymes activities and biochemical procedures (Vezina et al., 1997).
The cell membrane has negative charge because of carboxylic and phosphoric groups. Calcium acts as electrical bridge between these charges and makes the membrane stable and firm. In the other hand, the existence of these ions in the space between cells in pectin compounds causes the stability of the tissue (Marschher, 1995). In addition to increasing cytosolic calcium, salt-stress induces ABA accumulation (Ohta et al., 2003).
The membrane firmness has a direct effect on semi-osmosis parameter of membrane. Therefore, the insufficient amount of calcium around the cell (because of non-transferring or non-osmosis) destroys the semi-osmosis parameter, the permeation of intra-cell material to out and the penetration of useless material into the cell and it causes finally the death of cell. Then the concentration of 1 to 5 mM of this ion around the cell is necessary for surviving of membrane (Poovaiah, 1988).
The SOS pathway regulates ion homeostasis under salt stress. An unknown salt-stress sensor induces cytosolic calcium signals, which are transduced by the SOS3-SOS2 kinase complex (Chinnusamy et al., 2005 ).
The stresses such as lack of water, saltiness, Aluminum poisonous effect and low pH which make disorder in permeation and transfer of calcium results in reduction of out-cell concentration of this ion and distraction of membrane firmness. Consequently, release of out-cell material such as potassium has been seen. Some research results show that under stress conditions, the calcium ion prevents potassium from emission and causes the cell resistance (Kuiper, 1985).
Palta et al. (1977) and his colleagues recorded that in cold conditions some amount of calcium is separated from cell membrane. Zsoldos and Karvaly (1978), recorded that calcium at cold conditions can play an important role in supporting the cell membrane, Palta and Li (1978 and 1980), indicated that the main reason of emission of potassium from cell at the primary stage of cold stress is membrane weakness and lack of its semi-osmosis parameters. This is resulted from separating calcium from out-surface of membrane. Therefore, the increase of out-cell concentration of calcium in cold conditions have an important role in improving the membrane resistance and consequently the plant withstanding toward low temperature stress. The aforementioned issue about wheat cell was proved by (Pomeroy and Andrews, 1985).
Dhidsa et al. (1993), proved that temperature reduction from 25 to 10°C has no significant effect on transferring of calcium into medicago cells. But after its reduction to 4°C, the amount of transferring increases about 15 times.
The first defense line against the entrance of Na+ into the plant is the membrane of root cells which has low penetrability toward Na+, but the same cells penetrate potassium against concentration gradient. In the less compatible plants against salt, because of high amount of Na+ concentration in root area, their potassium penetration decreases. In adverse, their Na+ penetration and transferring to the aboveground organs increase. The main reason is disappearing of semi-osmosis parameter of membrane. It is recognized that the existence of calcium is necessary for maintenance of membrane heath whereas Na+ results in disappearing of semi-osmosis parameter of membrane by separating calcium from membrane and replacing it (Glenn et al., 1997).
The recovering role of Ca+2 is probably related to the maintenance of cell firmness and plasma membrane activities in root and aboveground organs (Lauchli, 1990). Adding calcium neutralizes the effects of Na+ i.e., disordering of calcium homeostasis (Rengal, 1992). Besides, calcium addition results in reduction of replaced Na+ in cell walls (Stassart et al., 1981) and plasma membrane (Lynch et al., 1987). Therefore, it prevents from membrane leakage (Picchioni et al., 1991). Finally, it prevents from growth reduction resulted from production decrease and cell lengthening (Zidan et al., 1990). Salt affects significantly on the rate of calcium penetration and transfer. Therefore, plants clearly showed the signals of lack of calcium especially in sensitive genotypes (Caramer et al., 1994).
At salty conditions, adding calcium into root area has no significant effect on biomass of imposed types of medicago. On the contrary, the function of sensitive types increases by adding calcium (Khan et al., 1998).
The current study was conducted to assess the effect of calcium on the cold and salt resistances of Medicago polymorpha cv. Santiago seeds in germination stage and recognition of morphological and physiological changes resulted from cold and salt stresses and the effective factors in increase of the resistance of Medicago polymorpha cv. Santiago to stress conditions.
MATERIALS AND METHODS
The experiment was conducted in the Laboratory by using the germinator on Medicago polymorpha cv. Santiago species at Agriculture College of Tarbiat Modarres University for two months in 2006. The seeds were sterilized in sodium hypochlorite during 7 min. Then, they were washed three times by sterilized distilled water.
In order to perform calcium treatment, the seeds were incubated in 0, 5, 10, 15 and 20 mM CaCl2 for 24 h. Then, 25 healthy seeds were selected and placed into the Petri dishes between 2 filter papers (Petri dishes were sterilized in ethanol 98%). In order to perform salt treatment, the 0, 50, 100, 150 and 200 mM. NaCl salt concentrations were added to the dishes contained exogenous calcium treated seeds. So, the seeds were completely soaked in solutions. The Petri dishes were placed into the germinator for 14 days at 5 and 15°C.
The experiment was conducted at the factorial plan based on completely randomized plots with 3 replications. On the 14th day, the number of germinated seeds, normal and abnormal seedlings, length of root and shoot, fresh and dry weight of plants and the ratio of shoot to root were determined. The analysis of data was carried out by SAS software and the comparison of means by Duncan’s Multiple Range Test (DMRT) at 5% level.
RESULTS AND DISCUSSION
The main effects of salt, Ca, temperature, germination and their interactions on length of root and shoot and their ratio, dry weight of embryos, the number of germination and its percentage were significant at stage 1% (Table 1).
The maximum of dry weight, number and percentage of germinated seeds, length of root and shoot and their ratio was at 15°C and their minimum was at 5°C (Table 2). Regarding to the recorded results, it can be stated that at 15°C, the enzymes activities and their effects on penetrability of seed membrane increase. Therefore, the germination and the growth of medicago embryos increase.
The highest reducing effect of salt on germination was received at 200 mM concentration of NaCl (Table 3) which is because of the negative osmosis and poisonous effect of salt concentration on germination.
The maximum root and shoot length was at 20 mM concentration of calcium, the greatest number and percentage of germinated seeds at 10 mM and the highest ratio of shoot to root and amount of dry material at 5 mM (Table 4) show the positive effect of calcium on germination and growth of Medicago polymorpha seedling.
The effect of calcium on membrane firmness and its fluidity has been seen and recorded through various experiments (Paliyath et al., 1984).
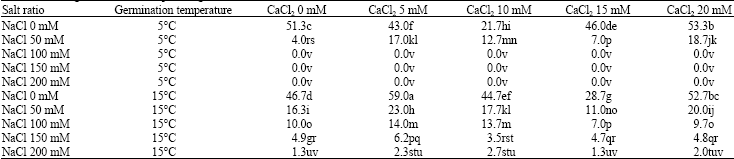
The interaction of temperature with calcium and salt showed that the longest shoot (59 mm) was produced at 15°C and 5 mM of calcium and zero concentration of salt. The shortest shoot was produced at 5°C and 100, 150 and 200 mM of salt and all levels of Ca. The recorded results also showed that the seed treatment with calcium before germination has a great effect on increasing the salt resistance at germination stage of Medicago polymorpha cv Santiago which is related to the significant effect of calcium on membrane (Table 5).
The longest root (33.3 mm) was perceived at 15°C, zero concentration of salt and 20 mM of calcium and the shortest one at 5°C in 100, 150 and 200 mM of salt and all levels of calcium.
| Table 1: | Variance analysis of temperature and calcium effect on germination and growth of Medicago polymorpha cv. Santiago under different salt concentrations |
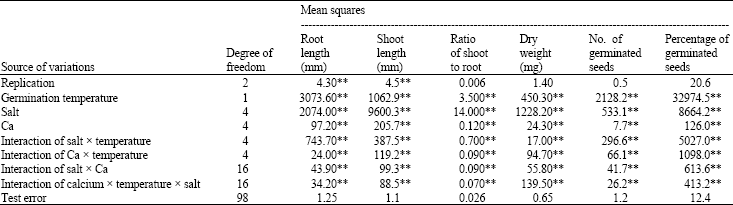 | |
| No star: Not significant; **Significant at level 1% | |
| Table 2: | The comparison of means of temperature effect on Medicago polymorpha cv. Santiago by Duncan’s Multiple Range Test (DMRT) (at level 5%) |
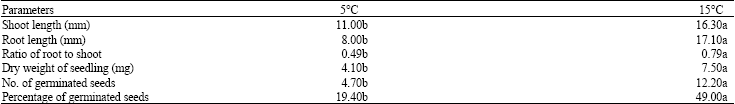 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 3: | The comparison of means of salt effect on effective parameters in growth and germination of Medicago polymorpha cv. Santiago through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
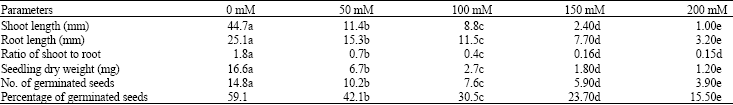 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 4: | The comparison of means of calcium effect on effective parameters in growth and germination of Medicago polymorpha cv. Santiago through Duncan’s Multiple Range Test (DMRT) (at stage 5%) |
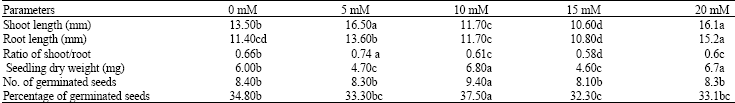 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 5: | The comparison of means of calcium and salt interactions on shoot length in Medicago polymorpa cv. Santiago at different germinating temperature through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 6: | Comparisons of mean interactions of calcium and salt effect on root length of Medicago polymorpha cv. Santiago at different germinating temperature through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
The results showed that calcium treatment has a great influence on reduction of negative effect of salt on root growth so that at 5 and 10 mM of calcium and different salt levels, the root growth increased (Table 6).
Dhidsa et al. (1993) proved that temperature reduction from 25 to 10°C has no significant effect on calcium transmission into Medicago polymorpha cells. However, its reduction to 4°C increases the transmission amount 15 times.
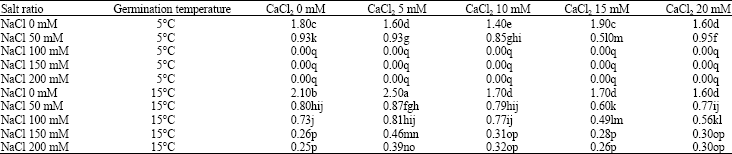
The maximum of ratio of shoot and root (2.5) was produced at 15°C germination, zero concentration of salt and 5 mM of calcium and its minimum at 5°C, in 100, 150 and 200 mM of salt and all levels of calcium. The results showed that salt resistance of seeds, treated in 5 mM of calcium before germinating stage, increases. Therefore, they have bigger ratio of shoot and root than other treatments (Table 7).
The highest amount of seedling dry matter (30.7 mg) was produced at 15°C, in zero concentration of salt and 20 mM calcium and the lowest one at 5°C in 100, 150 and 200 mM salt and all levels of Ca. The result showed the seeds treated before germination, have more salt resistance and dry matter than the ones without using calcium (Table 8).
It was perceived that adding calcium to environment in cold conditions has no meaningful effect on biomass of studied types of medicago. However, the function of sensitive types increases by adding calcium (Khan et al., 1998).
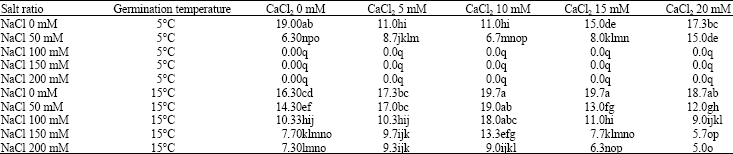
The highest number of germinated seeds was recorded as 19.7 and maximum percentage of germinated seeds was produced as 78.7% at 15°C, zero concentration of salt, 10 and 15 mM calcium. Their lowest values were observed at 5°C in 100, 150 and 200 mM salt and all concentration levels of Ca.
The results showed that pre-treatment of seeds with calcium causes the increase of number and percentage of germinated seeds at different salt levels and zero level of calcium (Table 9).
The improving action of Ca+2 is probably related to the protection of cell firmness and activities of plasma membrane of root and aboveground organs (Rengal, 1992).
| Table 7: | Comparison of mean interaction of calcium and salt effect on the ratio of shoot and root in Medicago polymorpha cv. Santiago at different germinating temperature through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 8: | Comparison of mean interactions of calcium and salt effect on dry material of Medicago polymorpha cv. Santiago embryo at different germinating temperature through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
| Table 9: | Comparison of mean interaction of calcium and salt effect on the number of germinated seeds of Medicago polymorpha cv. Santiago at different germinating temperature through Duncan’s Multiple Range Test (DMRT) (at level 5%) |
 | |
| Dissimilar letter(s) in each row have 5% significant difference | |
Besides, adding calcium to the root area results in reduction of Na+ replaced in cell wall (Stassart et al., 1981) and plasma membrane (Lynch et al., 1987). Therefore, it prevents or decreases the membrane leakage (Picchioni et al., 1991) and finally, it prevents growth decline resulted from production lengthening and reduction (Zidan et al., 1990).
With regards to the recorded results, it can be stated that; firstly, the temperature has a significant role on germinating of Medicago polymorpha cv. Santiago seeds. The increase of temperature causes the increase of enzyme activities and seed metabolic procedures. Consequently, absorption strength of seedlings, number of germinated seeds and weight of seedlings in the cold condition increase. Also, increase of salt concentration causes germination and seedling growth decreased because of harmful osmotic and poisonous effects. Thirdly, pre-treated M. polymorpha seeds, as annual medicago with calcium, results in the increase of seedling growth, percentage and number of germinated seeds.
Finally, the results of this research showed that pre-treatment of M. polymorpha seeds and calcium with 5 and 10 mM concentration causes enhancement of seedling tolerance to salinity, number and percentage of germinated seeds. It will make seedlings grow better. This is a result of protecting role of calcium in cells and cell membrane. It protects cell from denaturizing in high salt concentration. It also prevents from toxicity of Na+ on the membrane and prevents destruction of cell membrane. Decrease of calcium behaves as the second messenger to increase the cell tolerance against the environmental stress. Temperature reduction causes changes in concentration of biochemical regulator enzymes.
Decrease of cell membrane activity results in ions and material transfer. Changes of intercellular enzyme activities (decrease or increase) stimulate production of various compounds. Some are toxic but others are useful and they increase tolerance against coldness (Cattivell and Bartela, 1992). Cell membrane and cell wall have negative charge because of carboxylic and phosphoric groups. Calcium acts as an electric bridge between these charges and strengthens cell membrane and cell wall. On the other hand, existence of such ion in the inter-cellular space strengthens tissue in the pectinic compounds (Marschher, 1995).
REFERENCES
- Cramer, G.R., G.J. Alberico and C. Schmidit, 1994. Salt tolerance is not associated with the sodium accumulation of two maize hybrids. Funct. Plant Biol., 21: 675-692.
CrossRefDirect Link - Khan, M.G., M. Silberbush and S.H. Lips, 1998. Response of Alfalfa to potassium, calcium and nitrogen under stress induced by sodium chloride. Biol. Plant, 40: 251-259.
CrossRef - Kuiper, P.J.C., 1985. Environmental changes and lipid metabolism of higher plants. Physiologia Plantarum, 64: 118-122.
CrossRef - Lauchli, A., 1990. Calcium, salinity and the plasma membrane. Curr. Top. Plant Physiol., 4: 26-35.
Direct Link - Ohta, M., Y. Guo, U. Halfter and J.K. Zhu, 2003. A novel domain in the protein kinase SOS2 mediates interaction with the protein phosphatase 2C ABI2. Proc. Nat. Acad. Sci. USA., 100: 11771-11776.
Direct Link - Rengel, Z., 1992. The role of calcium in salt toxicity. Plant Cell Environ., 15: 625-632.
CrossRefDirect Link - Zidan, I., H. Azaizeh and P.M. Neumann, 1990. Does salinity reduce growth in maize root epidermal cells by inhibiting their capacity for cell wall acidification?. Plant Physiol., 93: 7-11.
Direct Link