
Ali Kemal Ayan
The High School of Profession of Bafra, University of Ondokuz Mayls, Samsun, Turkey
Ertan Sait Kurtar
The High School of Profession of Bafra, University of Ondokuz Mayls, Samsun, Turkey
Cuneyt Cirak
Department of Agronomy, Faculty of Agriculture, University of Ondokuz Mayls, Samsun, Turkey
Kudret Kevseroglu
Department of Agronomy, Faculty of Agriculture, University of Ondokuz Mayls, Samsun, Turkey
Journal of Agronomy
Year: 2004 | Volume: 3 | Issue: 4 | Page No.: 296-300







ABSTRACT
The research was conducted out to evaluate the effects of shading and plant growth regulators on bulb yield and some plant characters of summer snowflake in Samsun, Turkey ecological conditions for three years. Experimental design was a factorial randomized block arrangement with three replications. First two year, planted bulbs were left to growth and in the third year, at the beginning of vegetation, selected plots were shaded by 0.635 cm polyethylene mesh and plant growth regulators GA3 (50, 100, 250 and 500 ppm) and NAA (500, 1000, 2500 and 5000 ppm) were sprayed to both shaded and unshaded plots. Four applications were made with a one week interval for PGRs. According to the results, both shading and PGR treatments had a significant effects on the parameters evaluated.
PDF Abstract XML References Citation
How to cite this article
Ali Kemal Ayan, Ertan Sait Kurtar, Cuneyt Cirak and Kudret Kevseroglu, 2004. Bulb Yield and Some Plant Characters of Summer Snowflake (Leucojum aestivum L.) Under Shading as Affected by GA3 and NAA at Different Concentrations. Journal of Agronomy, 3: 296-300.
DOI: 10.3923/ja.2004.296.300
URL: https://scialert.net/abstract/?doi=ja.2004.296.300
DOI: 10.3923/ja.2004.296.300
URL: https://scialert.net/abstract/?doi=ja.2004.296.300
INTRODUCTION
Summer snowflake (Leucojum aestivum L.) called as akçabardak, kabalak, sarıklı kökü by Turkish folk is a perennial bulbous living in high and moist pastures of North Anatolia[1]. It is an important export material for our country. Likewise, 6,480,080 bulbs were exported in 1999[2]. But the plant is face to face with extinction, because its exported bulbs are obtained rather than by collecting in pastures[3]. This plant has recently attracted scientific concern due to its various secondary metabolite contents[4]. Its ethanol extract exhibits antiviral effect[5] and have some secondary metabolites, namely galanthamine, lectins and chelidonic acid, having multifunctional pharmacological effects[6]. Galanthamine found in whole plant and especially in bulbs is a dibenzofuran-type alkaloit[7] and its anticholineesterase[8,9], anaesthetic[10], analeptic and analgesic[11] and antimalarial[12] effects were demonstrated by relevant studies. Besides, the alkaloid has been used in treatment of Alzheimer[13], infantile paralysis and some kind of neurological disorders[14].
Light has been recognized a critical factor for production of high quality crop and light availability is believed to be one of major factors controlling main physiological functions in plants[15]. The desire to manipulate plant growth, while maximizing yield, has led to interest in Plant Growth Regulators (PGR)[16]. PGRs encompass a broad category of compounds that promote, inhibit, or otherwise modify plant physiological or morphological processes and they are have been widely used in crop production in an attempt to adjust plant growth and to improve yield and quality[17,18]. To author’s knowledge, there is no report on the effects of light reduction and plant growth regulators on summer snowflake. Here, we report our investigation of shading and plant growth regulator treatments on this plant.
MATERIALS AND METHODS
Materials: Summer snowflake bulbs used as material were obtained from Bilge Incorporated Company, Trabzon, Turkey. Plant growth regulators GA3 and NAA were purchased from Sigma Chemical Co.
The experimental area: This Study was conducted at Black Sea Agricultural Research Institute, Samsun located on The Black Sea coast of Turkey between 2000 and 2002. During the experimental period, annual precipitation was 698, 637 and 729 mm for 2000, 2001 and 2002, respectively.
Some major soil characteristics were found to be as follows; the soil texture is clay-loam, pH is 6.45, organic matter is 3.03%; extractable P by 0,5 N NaHCO3 extraction is 4.2 mg kg-1; exchangeable K by 1 N NH4OAc extraction is 61 mg kg-1 and EC is 1.42 mS cm-1 in soil saturation extract.
The experimental procedures: Experimental design was a factorial randomized block arrangement with three replications. Each treatment plot was 3x7 m with a distance of 0.5 m each plot. Bulbs were planted in the experimental area fertilized by 8 ton ha-1 farm manure with 40x10 cm plant-row spacing in September of 2000. Shading was performed as following manner: light was reduced at the rate of 50% of sunlight-exposed control by 0.635 cm polyethylene mesh tied on the tope and south and east sides of the selected plots in March, 2002. PGRs, GA3 and NAA at the concentrations 50, 100, 250 and 500 ppm and 500, 1000, 2500 and 5000 ppm, respectively were sprayed to both shaded and unshaded plots. Four applications were made with a one week interval by a CO2 backpack with a single nozzle wand and a length of hose to place the PGRs between rows and on the canopy. Pressure and nozzle size were adjusted to deliver the appropriate rate. PGR concentrations used in the experiment were the average rates for commercial uses suggested by chemical company. Mature bulbs were harvested at the beginning of June, 2002 after ignoring 0.5 m area from all sides of the plots. During vegetation, plant height and leaf number per plant; after harvesting, bulb diameter, bulb height, lateral bulb number per harvested bulb and bulb yield were determined as plant characters for each plot. The data were objected to ANOVA and differences among treatments were tested Duncan Multiple Range Test (Level of significance p<0.05).
RESULTS
According to the results of the variance analysis, plant height and bulb yield increased with both shading and PGR treatments (Table 1); while bulb height was affected by only PGR treatments and bulb diameter by shading (p<0.05). Besides, an interaction was found to be significant between shading and PGR treatments in term of leaf number per plant (p<0.05). On the contrary of other parameters evaluated here, lateral bulb number per bulb was not affected by treatments tested.
As mean of shaded and light conditions all treatments increased plant height when compared to control. GA3 at 500 ppm gave the highest plant height with 46.30 cm and it was followed other GA3 doses found in the same statistical group (43.90, 43.85 and 43.72 cm, respectively). In general, higher values for plant height were observed in shaded plots. Both PGR and shading treatments resulted in an increase in bulb yield (Table 1). All PGR treatments increased bulb yield when compared to control (419.17, 404.17, 390.00, 386.17, 381.67, 354.17, 340.50 and 320.00 kg da-1 for 50 ppm GA3, 5000, 500 and 1000 ppm NAA, 500 and 100 ppm GA3, 2500 ppm NAA and 250 ppm GA3, respectively). Also, shading increased bulb yield significantly (342.78 kg da-1 for sunlight exposed plots and 384. 44 kg da-1 for shaded plots). In term of bulb height, PGR were the only treatments affecting it and all plots treated with PGR gave higher values when compared to untreated control. Similarly, bulb diameter was affected by only shading. As mean of PGR, the higher value with 27.58 mm was obtained from shaded plots. It is interesting to note that there was an interaction between shading and PGR treatments for leaf number per plant since no interaction was observed between them for other parameters evaluated. In term of this interaction, plots treated with 1000 ppm NAA under shading gave the highest leaf number per plant (6.63) and the lowest values were obtained from untreated but shaded control (4.70).
DISCUSSION
The experiment was conducted out to investigate the effects of shading, combined with aforesaid doses of GA3 and NAA, on bulb yield and some plant characters of summer snowflake. Previous reports clearly demonstrate that the effect of shading on plant growth and development varied with greatly depending on species. Experimental shading reduced the light reaching a shallow seagrass meadow to less than 10% of incident light. Shaded seagrass Posidonia australis had significantly lower leaf growth rate, shoot density, shoot weight and epiphyte weight than seagrass in control plots[19]. Reduction of irradiance to 35 and 25% of control levels resulted in a 25-50% decrease in leaf cluster density in Heterozostera tasmanica[20]. Leaf elongation rates in Thalassia testudinum plants decreased relative to unshaded controls after one month of shading treatment[21]. Total grain yield was 5% lower after shading (60% ambient radiation) in Hordeum vulgare[22]. Similarly, yield was reduced by all levels of shade, owing to the combined effect of reduced bunch weight and a lower stand in banana[23]. On the contrary of the results from above mentioned study, plants subjected to 92% light reduction showed the most pronounced effects in chlorophyll a concentration, above- and below-ground biomass and leaf dimensions in Vallisneria americana[24]. Higher stomatal conductance and higher CO2 assimilation rates were observed in shaded plants, so that integrated daily net CO2 uptake was approximately 20% higher than in exposed plants in citrus[25]. Low irradiance (mean total shortwave radiation of 2.1 or 6.3 MJ m-2 day-1 compared to 10.5, 14.6 or 20.9 MJ m-2 day-1) increased shoot elongation and reduced leaf area, plant dry weight and floral bud and open flower number in the passion fruit[26].
| Table 1: | Bulb yield and some plant characters of summer snowflake under shading as affected by GA3 and NAA |
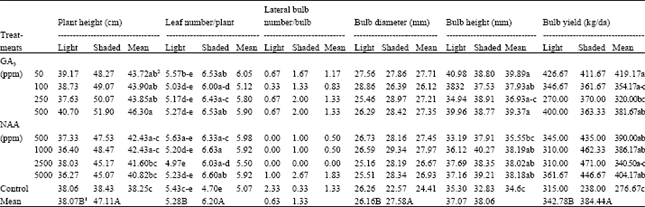 | |
| 1Values followed by different capital letter in bottom row and 2small letter(s) in each column are significantly different (p<0.05) according to Duncan Multiple Range test | |
The most important plant characters for fruit production were best in pepper plants grown under 30% shading. This shade increased stem and root dry weight which favored fruit production. Also, 30% shading increased branch number which enhanced fruit setting and favored early flowering[27]. According to our results, shading treatment resulted in significant increase in bulb yield and diameter, leaf number per plant and plant height. Even though light is the ultimate substrate for photosynthetic energy conversion, it can also harm plants[28,29]. Maximum level of photosynthetic light saturation varied with each plant species greatly and to author’s knowledge, it is unknown for summer snowflake. At light levels above the saturation point, carbon metabolism may limit the consumption of photosynthetic energy which results in excess photon absorption. As a consequence, non-utilized excitation energy can accumulate, promoting reductions in photosynthetic efficiency, along with consequent yield reduction[30]. The frequency of this damage is relatively low under normal conditions, but becomes a significant problem for the plant with increasing light intensity. This is true for, especially, geophytes like summer snowflake. Yield and bulb height increase in shaded plots were probably due to this reason. In term of the increase in plant height, the decreased light intensity due to increased shading should promoted greater cell elongation and enlargement resulting in taller plants[31].
PGRs are now becoming very important as an effective and often relatively low-cost, means of improving crop production. There are now a range of products to regulate plant growth, harvest period and plant physiology. There are a number of studies related to field using of PGR. In a field study, The effects of three concentrations of BA, GA4+7 and Promalin (a mixture of BA and GA4+7) on branching and flower production of three jojoba clones were studied. The growth regulators had a significant effect on both branching and flower bud production and clones differed in their response. For the most responsive clone, the most effective treatments were 100 ppm GA4+7 and 100 ppm Promalin and these resulted in a 133 and 110% increase in flower buds. The results of these studies indicate that growth regulators can be used to significantly increase branching and flower bud production on jojoba[32]. Bean plants were sprayed with the growth regulators uniconazole (25 and 50 ppm), paclobutrazol (100, 200, 400 and 800 ppm) and cytogen (5 and 10 ppm). Cytogen affected plant height positively and in addition to uniconazole at 25 ppm significantly increased plant fresh weight, number and weight of pods per plant as well as total yield[33]. Soil application of plant growth regulator PGR-4 increased root length, branching and yield in soybean[34]. These results are in accordance with these reported by us. But it is interesting to note that there were no difference among GA3 and NAA doses in term of their growth and yield enhancing effects whereas the difference was significant when compared to control under both shaded and sun light exposed plots. It may be due to that GA3 and NAA doses tested were up to the concentrations, probably sufficient for supplying maximum yield and growth of summer snowflake.
Consequently, results from the present study indicated that irradiance is one of the major environmental factors limiting plant growth and bulb yield of summer snowflake. If this medicinal plant is thought to cultivate under field conditions, shading will be a very useful practice to enhance bulb yield. PGRs GA3 and NAA can be used to obtain bigger bulbs, but further studies will be needed to determine the most suitable concentrations for this plant.
REFERENCES
- Hudson, J.B., M.K. Lee, B. Sener and N. Erdemoglu, 2000. Antiviral activities in extracts of some Turkish medicinal plants. Pharm. Biol., 38: 171-175.
Direct Link - Shen, Z.W., U. Fisinger, A. Poulev, W. Eisenreich and I. Werner et al., 2001. Tracer studies with 13C-labelled carbohydrates in cultured plant cells. Retrobiosynthetic analysis chelidonic acid biosynthesis. Phytochemistry, 57: 33-42.
CrossRefDirect Link - Jorg, E., T. Takada, Y. Kita and M.H. Zenk, 1998. Biosynthesis of the amary llidaceae alkaloit galanthamine. Phytochrmotography, 49: 1037-1047.
Direct Link - Cirak, C., A.K. Ayan, E.S. Kurtar, K. Kevseroglu and N. Camas, 2004. The Effects of different N doses and harvesting times on bulb yield and some plant characters of summer snowflake (Leucojum aestivum L.). Asian J. Plant Sci., 3: 193-195.
CrossRefDirect Link - Orhan, I. and B. Sener, 2003. Bioactivity-directed fractionation of alkaloids from some amaryllidaceae plants and their anticholinesterase activity. Chem. Natural Comp., 39: 383-386.
CrossRefDirect Link - Heinrich, M. and H.L. Teoh, 2004. Galanthamine from snowdrop-the development of a modern drug against Alzheimer`s disease from local Caucasian knowledge. J. Ethnopharmacol., 92: 147-162.
CrossRefPubMedDirect Link - Schwikkard, S. and F.R. van Heerden, 2002. Antimalarial activity of plant metabolites. Nat. Prod. Rep., 19: 675-692.
CrossRefPubMedDirect Link - Matsumoto, J., M. Hiroyuki and W. Izumi, 2000. Whole plant carbon gain of an endangered Herbaceous species Aster kantoensis and the influence of shading by an alien grass Eragrostis curvula in its gravelly floodplain habitat. Ann. Bot., 86: 787-797.
Direct Link - Patricia, S., L. Sviatoslav, B. Andre and G. Armand, 1999. Morphological and anatomical modifications in winter barley culm after late plant growth regulator treatment. Eur. J. Agron., 11: 45-51.
Direct Link - Fitzpatrick, J. and H. Kirkman, 1995. Effects of prolonged shading stress on growth and survival of seagrass Posidonia australis in Jervis Bay, New South Wales, Australia. Mar. Ecol. Prog. Ser., 127: 279-289.
Direct Link - Czerny, A. and K.H. Dunton, 1995. The effects of in situ light reduction on the growth of two subtropical seagrasses, Thalassia testudinum and Halodule wrightii. Estuaries, 18: 418-427.
Direct Link - Grashoff, C. and L.F. d'Antuono, 1997. Effect of shading and nitrogen application on yield, grain size distribution and concentrations of nitrogen and water soluble carbohydrates in malting spring barley (Hordeum vulgare L.). Eur. J. Agron., 6: 275-293.
Direct Link - Israeli, Y., Z. Plaut and A. Schwartz, 1995. Effect of shade on banana morphology, growth and production. Sci. Hortic., 62: 45-56.
Direct Link - Janis, C.K., F. Diane, J.M. Yates, L.Q. Robert, J. Fred, A. Cynthia and D. Richard, 2003. Effects of light reduction on growth of the submerged macrophyte Vallisneria americana and the community of root-associated heterotrophic bacteria. J. Exp. Mar. Biol. Ecol., 291: 199-218.
Direct Link - Medina, C.L., R.P. Souza, E.C. Machado, R.V. Ribeiro and J.A.B. Silva, 2002. Photosynthetic response of citrus grown under reflective aluminized polypropylene shading nets. Sci. Hortic., 96: 115-125.
CrossRef - Cohen, S., S. Moreshet, L.L. Guillou, J.C. Simon and M. Cohen, 1997. Responses of citrus trees to modified radiation regime in semi-arid conditions. J. Exp. Bot., 48: 35-44.
Direct Link - Barber, J. and B. Andersson, 1992. Too much of a good thing: Light can be bad for photosynthesis. Trends Biochem. Sci., 17: 61-66.
PubMedDirect Link - Oosterhuis, D.M. and D. Zhao, 1994. Increased root length and branching by soil application of the plant growth regulator PGR-IV. Plant Soil, 167: 51-56.
Direct Link