
A.M. Abdel-Moneim
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
N.M. Abou Shabana
National Institute of Oceanography and Fisheries, Alexandria, Egypt
S.E.M. Khadre
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
H.H. Abdel-Kader
Department of Zoology, Faculty of Science, Alexandria University, Alexandria, Egypt
International Journal of Zoological Research
Year: 2008 | Volume: 4 | Issue: 4 | Page No.: 189-202







ABSTRACT
This study investigated the toxicity of chemical dye production industry wastewaters and emphasized the importance of toxicity tests in wastewater discharge regulations. The dyestuff samples were obtained from an untreated effluent at Kafr El-Dawar (Egypt) during 5 months from December 2005 to April 2006. The effluent was near-neutral, strongly colored and highly ecotoxic with a COD content of 240 mg L-1. Catfish (Clarias lazera) were used as test organisms. The 96 h LC50 of C. lazera exposed to the effluent was found to be 35%. Twenty eight-day exposures to 3.12, 6.25 and 12.5% doses were conducted and several toxicological endpoints were evaluated. The predominant haematological change was a severe anaemia. Leucocytes count increased in stressed fish. Moderate neutrophilia and lymphopenia were detected in fish exposed to effluent concentrations of 6.25 and 12.5%. Monocytosis was quite evident at 12.5% concentration. Overall increases over control were observed in serum transaminases (GPT and GOT). However, these increases were not dose dependent. Blood urea insignificantly changed in treated fish while serum creatinine significantly elevated after exposure to 3.12 and 12.5% dose levels. In addition, free amino acids in flesh exhibited a decreasing trend at all concentrations tested. Moreover, histopathological alterations in the gills, liver and kidney occurred exclusively after treatment. Histological analysis of gill samples revealed a range of lesions including lamellar fusion due to hyperplasia and hypertophy of epithelial cells, subepithelial cell edema, collapsed pillar cell system and extensive lamellar aneurism. Liver pathologies included extensive necrosis of hepatocytes, cytoplasmic vacuolation, distended sinusoids with massive congestion and infiltration of inflammatory cells through out the liver parenchyma. In the kidney, the glomeruli appeared shrunken. The tubular epithelium was desquamated, vacuolated and often destroyed, which render the tubular system of the mesonephros incapable of functioning properly. From the present findings, it is assumed that under field conditions, the dyestuff effluent could induce general toxic effects and possibly might disturb ecologically relevant processes such as fish reproduction.
PDF Abstract XML References Citation
How to cite this article
A.M. Abdel-Moneim, N.M. Abou Shabana, S.E.M. Khadre and H.H. Abdel-Kader, 2008. Physiological and Histopathological Effects in Catfish (Clarias lazera) Exposed to Dyestuff and Chemical Wastewater. International Journal of Zoological Research, 4: 189-202.
DOI: 10.3923/ijzr.2008.189.202
URL: https://scialert.net/abstract/?doi=ijzr.2008.189.202
DOI: 10.3923/ijzr.2008.189.202
URL: https://scialert.net/abstract/?doi=ijzr.2008.189.202
INTRODUCTION
In urban aquatic environments fish may be exposed to a myriad of substances at the same time produced by different kinds of anthropogenic activities. Biological changes in fish that are related to the exposure or to the effects of contaminants are called biomarkers and their use has led to good results in environmental risk assessment (Van der Oost et al., 2003). The use of physiological and biochemical parameters as indicators of water quality has recently been developed to detect sublethal impacts of pollutants. Prominent among these biomarkers are haematological data (Lohner et al., 2001;Cazenave et al., 2005) and physiological variables, such as plasma levels of metabolites (DiGiulio et al., 1995) and ions (Engelhardt et al., 1981; Alkindi et al., 1996; Martinez and Souza, 2002), levels of hormones like cortisol (Hontela et al., 1996; Barton et al., 1998; Hontela, 1998; Benguira and Hontela, 2000) and biochemical variables such as detoxifying enzyme activities (Paris-Palacios et al., 2000; Teles et al., 2003). Similarly, histopathological changes have been widely used as biomarkers in the evaluation of the health of fish exposed to contaminants, both in the laboratory (Wester and Canton, 1991; Thophon et al., 2003) and field studies (Schwaiger et al., 1997; The et al., 1997). One of the great advantages of using histopathological biomarkers in environmental monitoring is that this category of biomarkers allows examining specific target organs, including gills, kidney and liver, that are responsible for vital functions, such as respiration, excretion and the accumulation and biotransformation of xenobiotics in the fish (Gernhofer et al., 2001). Furthermore, the alterations found in these organs are normally easier to identify than functional ones (Fanta et al., 2003) and serve as warning signs of damage to animal health (Hinton and Lauren, 1990).
Color is one of the most obvious indicators of water pollution and the discharge of highly colored effluents containing dyes can be damaging to the receiving bodies (Nigam et al., 1996). Al-Sabti (2000) proved the genotoxicity of Chlorotriazine Reactive Azo Red 120 textile dye using micronuclei induction in Prussian carp (Carassius auratus gibelio) erythrocyte cells under laboratory conditions. Recently, Sharma et al. (2006) reported higher sensitivity of fish RBCs (their counts and morphological abnormalities) to an azo dye methyl red in comparison to their mortality. However, pathophysiological alterations in fish after in vivo exposure to complex dyestuff effluent have not been thoroughly documented. With this background, the present study focused on the subchronic exposure of sexually mature Clarias lazera to wastewater from a chemical dye production industry, emphasizing the incorporation of toxicity parameters into Egyptian Environmental Regulations in order to protect receiving ecosystems. Along with physiological perturbations, histopathological changes in gills, liver and kidneys were investigated.
MATERIALS AND METHODS
Animals
C. lazera, a common freshwater teleost (length 22-27 cm and body weight 100-120 g), was selected as the test species owing to its hardy nature to survive under laboratory conditions and to its being sensitive to small environmental changes.
Maintenance of Test Species
C. lazera were collected from a relatively unpolluted source. A batch of 10 adult, healthy, disease free specimens were stocked in 10 glass aquaria measuring 60x30x30 cm each containing 30 L of water and acclimatized in the tap water for 15 days before they were subjected to dyestuff treatment. During the acclimatization process, the fish were fed ad libitum and the water was changed daily to discard the metabolic wastes. Fitness of the test animals is of prime importance in toxicity bioassay studies. Therefore diseased fish or fish showing any abnormal behavior were removed from the aquaria as soon as possible. Unnecessary handling of fish was also strictly avoided.
Dye and Chemical Wastewater
The dyestuff samples used during the present study were obtained from an untreated effluent at Kafr El-Dawar (Egypt) during 5 months from December 2005 to April 2006. The wastewater was a combination discharges from different processes in the industry producing dyestuffs of all groups including azo reactive dyes used in textile and paper industries, pesticides such as dimethoate and malathione and the active matter necessary for manufacture of detergents such as linear alkyl benzene sulphonic acid. The sample was delivered to laboratory at 0-6 °C within a few hours. The effluent was filtered through a 60-Am nylon plankton net to remove indigenous organisms. Then the effluent was kept frozen and thawed before use.
| Table 1: | Composition of dyestuff and chemical effluent at Kafr El-Dawar (Egypt) |
 | |
| *: Effluent was analyzed following standard methods of the examination of water and wastewater (APHA/AWWA/WEF, 1989), values are the averages of triplicate analyses of the samples, COD: Chemical Oxygen Demand, BOD: Biological Oxygen Demand, LSA: Linear Alkyl Sulphonate, ND: Not Determined | |
The effluent used in the present study was a complex mixture of several metals and hydrocarbons (Table 1). The effluent was near-neutral, strongly colored and highly ecotoxic with a COD content of 240 mg L-1.
Determination of 96 h LC50 of Dyestuff Effluent Complex and Selection of Exposure Concentrations
The 96 h LC50 of C. lazera exposed to the effluent was determined according to the graphical interpolation method (Verma et al., 2005). For the determination of median tolerance limits or LC50, fish were exposed to different concentrations of the effluent (25, 50, 75 and 100% v/v) in different aquaria. The test water was renewed and fresh pollutant added daily to remove the fecal waste and to maintain the concentration of effluent at a constant level. A concurrent control of 30 fish in three aquaria was maintained throughout the experimental duration under identical conditions. The median value was derived by plotting the experimental data on semilogarithmic coordinate paper, with test concentration on the logarithmic scale and the percentage of survival on the arithmetic scale. A straight line was drawn between the 2 points, which are above and below the 50% survival line. The point of intersection with the 50% survival line was considered LC50 which was found to be 35%. In the present study, adult catfish were exposed to dyestuff and chemical effluent, solubilized in dilute fresh water, at concentrations of 3.12% (9/100 LC50), 6.25% (18/100 LC50) and 12.5% (36/100 LC50).
Experimental Design
Twenty eight-day exposures to 3.12, 6.25 and 12.5% doses were conducted. Blood and tissues were collected from both control and experimental fish. After the exposure period, 10 fish per test group were anaesthetised to carry out both physiological and histopathological examinations.
Haematological Examinations
Blood samples were taken from the caudal vein and processed for haematological investigations according to Dacie and Lewis (1975). Haematological parameters include the total number of erythrocytes, the packed-cell volume (%) and the haemoglobin concentration (g dL-1). From the results obtained, the erythrocyte indices including the mean corpuscular volume (MCV), the mean corpuscular haemoglobin (MCH) and the mean corpuscular haemoglobin concentration (MCHC) were calculated. Furthermore, the total number of leukocytes was counted and a differential white blood cell count was performed.
Determination of Serum Liver Function Markers
Serum liver function enzymes, glutamate oxaloacetate transaminase (GOT) (aspartate transaminase; AST) and glutamate pyruvate transaminase (GPT) (alanine transaminase; ALT) were determined according to the reported procedures (Reitman and Frankel, 1957). The enzyme activity was expressed in terms of unit enzyme mL-1 of serum.
Renal Function Tests
Creatinine was determined according to Henry et al. (1974). In this method, creatinine reacts with picrate to form a coloured complex. The rate of formation of the complex is measured photometrically at 492 nm. Urea was determined by using the coupled urease/glutamate dehydrogenase (GLDH) enzyme system according to Tietz (1995).
Quantitative Determination of Free Amino Acids (FAAs) in Muscles
Samples of muscles of fish were homogenized in exactly three times their weight in ice-cold distilled water. Crude homogenates (3 g) were deproteinized by the addition of 10 mL g-1 sulfosalicylic acid (3.5%). The supernatant was collected after centrifugation at 3500 rpm for 15 min and stored at -16 °C for amino acid analysis. Determinations of FAAs were preformed using Beckman amino acid analyzer 119CL.
Histopathological Examinations
After complete necropsy of ten fish per group, tissue specimens of the gills, liver and kidney were removed, fixed in Bouin`s fluid and routinely processed for paraffin embedding. Sections were cut at 3 mm and stained with haematoxylin and eosin (H and E).
Statistical Analysis
Data are expressed as means ± SE. The results were computed statistically (SPSS software package, version 15) using one-way analysis of variance (ANOVA). Post-hoc test was performed for inter-group comparison using the LSD. Values of p<0.05 were considered statistically significant.
RESULTS
Haematological Results
As regard Hb content and RBC count, highly significant (p<0.05) decreases were detected in fish exposed to 3.12 and 6.25% concentrations of the dyestuff and chemical effluent for 28 days when compared to control values. Insignificant (p > 0.05) decreases of these parameters were evident at 12.5% concentration. Exposure to the effluent also caused a significant (p<0.05) decrease in haematocrit value in the treated fish. The results showed that MCV and MCHC did not change in fish after exposure to dyestuff and chemical effluent at concentrations of 3.12 and 6.25% whereas significant (p<0.05) decrease in MCV as well as significant (p<0.05) increase in MCHC were recorded in fish exposed to 12.5% of the effluent. MCH was not affected after treatment (Table 2).
Table 3 provides an overview on the calculated leucocyte values. Leucocytes count increased in stressed fish. The total number of leucocytes increased significantly (p<0.05) in fish exposed to 6.25 and 12.5% concentrations of dyestuff and chemical effluent. Fish exposed to 6.25% treatment showed highly significant (p<0.05) elevation in the relative percentage of the circulating neutrophils. The percentage of stab forms increased significantly (p<0.05) after exposure to 12.5% dose level. Furthermore, reading of the differential white blood cell count demonstrated a significant (p<0.05) lower percentage of lymphocytes in fish intoxicated with effluent concentrations of 6.25 and 12.5%. Moreover, monocytosis was quite evident at 12.5% concentration. In the exposed fish, the eosinophils percentages were only slightly affected in all treated groups. 3.12% effluent exposed fish did not reveal any significant (p > 0.05) changes in leucocytic parameters.
| Table 2: | Changes in erythrocyte values of Clarias lazera after exposure to different concentrations of dyestuff and chemical effluent for 28 days |
 | |
| Values are expressed as mean ± SE, for five animals in each group, means in a row with no common superscripts are significantly different (p<0.05) | |
| Table 3: | Changes in leucocyte values of Clarias lazera after exposure to different concentrations of dyestuff and chemical effluent for 28 days |
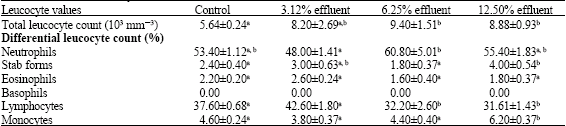 | |
| Values are expressed as mean ± SE, for five animals in each group, means in a row with no common superscripts are significantly different (p<0.05) | |
| Table 4: | Changes in serum biochemical parameters indicative of liver and kidney functions of Clarias lazera after exposure to different concentrations of dyestuff and chemical effluent for 28 days |
 | |
| Values are expressed as mean ± SE, for five animals in each group, means in a row with no common superscripts are significantly different (p<0.05), GPT: Glutamic Pyruvic Transaminase, GOT: Glutamic Oxaloacetic Transaminase | |
Changes in Liver and Kidney Functions
The liver function test was assessed by measuring the serum GOT (AST) and serum GPT (ALT) levels in control and treated fish (Table 4). Overall increases over control were observed in both parameters. However, these increases were not dose dependent. Pronounced increase of GPT activity was noted at 3.12% treatment (more than 4 folds higher as compared to control). However, a fall in the rate of increase in GPT levels was evident at 6.25% treatment. At 12.5% treatment, GPT activity increased approximately 5.5 folds over control. The GOT level increased maximally at 3.12% concentration (approximately 9 folds higher than control). This stimulation was further decreased at higher doses.
Blood urea insignificantly (p > 0.05) changed in treated fish as compared to control animals (Table 4). However, creatinine significantly (p<0.05) elevated after 28 days of exposure to 3.12 and 12.5% dose levels (approx. 50% higher as compared to control). Creatinine was observed to be insignificantly (p > 0.05) increased (34% over the control value) after exposure to 6.25% of the effluent.
Changes in Muscle Free Amino Acids
The free amino acids in fish muscles exhibited a decreasing trend at all concentrations tested (Table 5). Essential and non-essential amino acids were significantly reduced by elevated concentration of the effluent (ANOVA p<0.05).
| Table 5: | Changes in muscle free amino acids profile (μg g-1 wet tissue) of Clarias lazera after exposure to different concentrations of dyestuff and chemical effluent for 28 days |
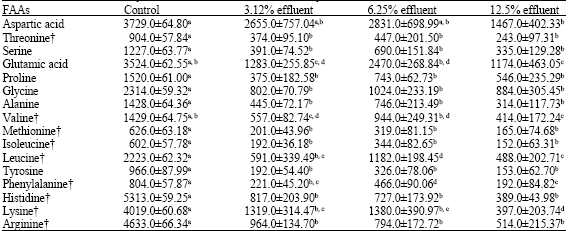 | |
| Values are expressed as mean ± SE, for 3 animals in each group, means in a row with no common superscripts are significantly different (p<0.05), †: Essential Amino Acids | |
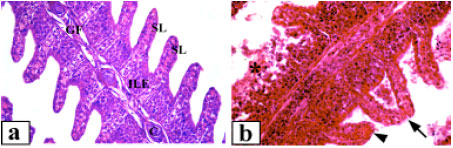 | |
| Fig. 1: | Gill tissue of Clarias lazera, (a) normal gill, gill filament (GF), secondary lamellae (SL), interlamellar epithelium (ILE) and hyaline cartilage (C) surrounded with darkly stained blood vessels and sinusoidal space, (b) after exposure to 3.12% of the effluent, lamellar tip fusion (arrow) with hypertrophied epithelium, lamellar aneurism (arrowhead), shortening of secondary lamellar and degenerated necrotic tissues (*) can be seen, (c) after exposure to 6.25% of the effluent, subepithelial edema (arrows) with hyperplasia of interlamellar epithelia and dilated blood capillaries (arrowheads), (d) after exposure to 12.5% of the effluent, hyperplasia (arrows) and deformed covering of the respiratory lamellae. Arrowheads point to deformed capillary channels. Magnification: 400x (a-d) |
Catfish Histopathology
Gills
Control gill tissues consisted of a primary filament and secondary lamellae. The secondary lamellae composed of a single layer of epithelial cells that were supported by pillar cells separating the capillary channels. One or two erythrocytes were usually recognized within each capillary lumen (Fig. 1a). Chloride cells were visible along the interlamellar epithelium. In fish exposed to 3.12% of the effluent, the pillar cell system displayed hemorrhagic foci. There was extensive dilation of blood capillaries and swollen lamellae packed with erythrocytes (i.e., aneurism) were quite evident (Fig. 1b). Furthermore, fusion at the tips of lamellae was observed, with edematous hypertrophy of epithelial cells. Necrotic changes in secondary lamellae were commonly noted with rupture of lamellar epithelium. Higher concentration of the effluent (6.25%) resulted in more pathological changes, including lamellar fusion due to simple apposition as well as hypertrophy and hyperplasia of epithelial cells (Fig. 1c). The fusion commonly occurred along the entire length of secondary lamellae. Moreover, subepithelial cell edema was also noted with collapsed pillar cell system. At subchronic exposure to 12.5% of the effluent, the quick death of fish observed at this concentration may be due to degenerative and necrotic changes of the pillar cells as well as extensive dilation of the capillary wall (Fig. 1d). There was extensive lamellar aneurism. Furthermore, hyperplasia of epithelial cells caused diffuse thickening of secondary lamellae. In some incidences, there was respiratory epithelial necrosis.
Liver
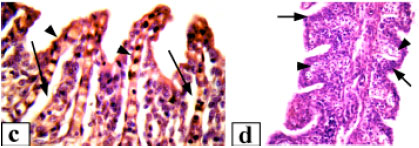
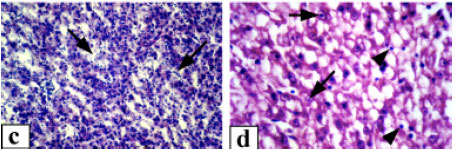
Control fish liver generally exhibited a normal architecture with hepatocytes presenting a homogenous cytoplasm and a large spherical nucleus containing one nucleolus and variable amounts of dispersed and peripheral heterochromatin (Fig. 2a). Hepatocytes were located among blood capillaries called sinusoids forming cord-like structures. Kupffer cells were found on the luminal surface of the sinusoid endothelium. With exposure to 3.12% of the effluent, the liver showed severe disorganization of the hepatic cords, damaged cell membrane and extensive necrosis of hepatocytes were observed (Fig. 2b). Hepatocyte cytoplasmic vacuolation was the most evident pathological alteration observed thus the cellular structure was totally obscured. Associated with these pathological changes, the nuclei of hepatocytes frequently exhibited karyolysis. Exposure to 6.25% of the effluent resulted in more notable detrimental changes. There was marked swelling of the hepatocytes.Sinusoids in most cases were distended with massive congestion (Fig. 2c). The liver cells were degenerated with necrosis which appeared in focal areas in association with mononuclear leucocyte infiltration. A large number of hepatocytes showed extensive pyknosis and Kupffer cells were hypertrophied. At exposure to 12.5% of the effluent complex, hepatic tissue of all fish showed degenerative changes such as cloudy swelling and necrosis (Fig. 2d). The liver displayed a spongy appearance due to severe lesions. In addition, the nuclei were karyolysed and the nucleoli were commonly terminalized. Massive infiltration of inflammatory cells through out the liver parenchyma was also observed.
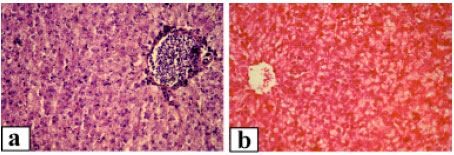 | |
| Fig. 2: | Hepatic tissue of Clarias lazera, (a) control, (b) after exposure to 3.12% of the effluent, massive necrosis of hepatocytes is evident, (c) after exposure to 6.25% of the effluent, necrotic tissue with dilation and congestion of sinusoids (arrows), (d) after exposure to 12.5% of the effluent, the cytoplasm of hepatic cells is cloudy, arrows point to degenerative nuclei and arrowheads indicate scattered leukocyte infiltration. Magnification: 400x (a-c), 1000x (d) |
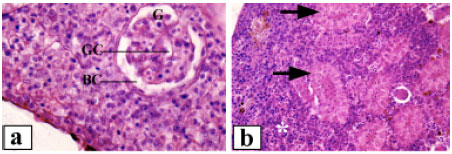 | |
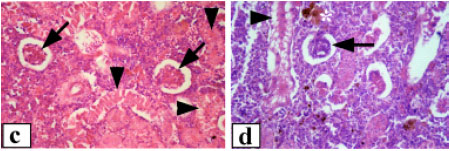
| Fig. 3: | Renal tissue of Clarias lazera, (a) control, glomerulus (G), glomerular capillaries (GC) and Bowman`s capsule (BC), (b) control, arrows point to renal tubules and (*) indicates lymphoid tissue, (c) after exposure to 3.12% of the effluent, arrows point to shrunken glomeruli and arrowheads indicate degenerated renal tubules, (d) after exposure to 6.25% of the effluent, arrow points to damaged glomerulus with increased mesangial matrix and arrowhead indicates severe vacuolation of tubular epithelium. Shrinkage of renal tubules can also be seen. (*) indicates hyaline droplets degeneration, (e) after exposure to 12.5% of the effluent, break down of glomerular capillaries (arrow) and lifting of tubular epithelium from its original position (*), (f) after exposure to 12.5% of the effluent, note the great sloughing of renal tubular epithelium with coagulative necrosis (arrow) and syncytial tubules (arrowheads). Magnification: 400x (a, c, d), 1000x (b, e, f) |
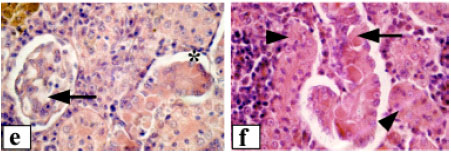
Kidney
The normal trunk kidney possessed a number of nephrons and interstitial lymphoid tissue. The nephron was made up of renal corpuscles and renal tubules. The renal corpuscles consisted of a cluster of capillaries (glomerulus) surrounded by Bowman`s capsule. The microscopic picture of the glomerulus showed capillary spaces covered by endothelial cells on the inner side. The space between capillaries was filled with mesangial cells. On the inner side of the Bowman`s capsule, epithelial cells were found (Fig. 3a). The convoluted renal tubules were covered by tall columnar cells with a weak eosinophilic cytoplasm and apical microvilli or brush border towards the lumen (Fig. 3b). The proximal tubules were characterized by well developed brush border along the apices of the cells. Alteration of renal corpuscles of fish exposed to 3.12% of the effluent included extensive damage of the glomeruli and renal tubules, which led to the gradual disappearance and occasional replacement of the tubules by interstitial lymphoid tissue (Fig. 3c). The glomeruli appeared shrunken with densely basophilic nuclei and there was an increased space within the Bowman`s capsule. The renal tubules revealed various degrees of degenerations. Extensive fibrosis was evident. Necrotic tubular cells with complete lysis of the cytoplasm and karyolysis of the nuclei were also noted. On the other hand, the intensity of glomerular and tubular damage was more severe in fish exposed to 6.25% of the effluent. The glomerular network showed frequent shrinkage and destruction, with increase of mesangial tissue. Consequently, the Bowman`s space was enlarged. The tubular epithelium was desquamated, vacuolated and often destroyed (Fig. 3d), which render the tubular system of the mesonephros incapable of functioning properly. The tubular lumen was obliterated in the majority of tubules. The nuclei were enlarged and vesicular, others were degenerated. In addition, hyaline droplet degeneration was occasionally observed. Higher concentration of the effluent (12.5%) resulted in massive pathological changes (Fig. 3e-f). The renal corpuscles exhibited severe damage including breakdown of glomerular blood capillaries. The tubules had lost their regular shape. Tubular shrinkage and syncytial tubules were common features. In addition, necrotic cells with complete lysis of cytoplasm were also noted.
DISCUSSION
The development of Whole Effluent Toxicity (WET) methodologies has become an important part of applied ecotoxicology in recent years (Smolders et al., 2002). Because effluents are a main source of direct and continuous input of pollutants in aquatic ecosystems, the study of the effects of effluent exposure on organisms has a high ecological relevance (Chapman, 2000). However due to the unknown and complex nature of effluents it is virtually impossible to relate observed effects to specific pollutants or even a class of pollutants (Guerra, 2000). The traditional WET test has always been mortality based but the inherent characteristics of effluents imply that WET should provide a holistic and integrative overview of the effects of toxic exposure on different biological processes at different levels of biological organization within the test organism(s) (Munkittrick and McCarthy, 1995; Wepener et al., 2005).
The haematological results documented that exposure to dyestuff and chemical effluent could result in severe anemia in catfish. Decreases in RBC count, Ht level and Hb content are symptoms of anemia (Koprucu et al., 2006). There are various factors to cause anemia in animals, such as hemorrhage, hemolysins and renal diseases (Roberts, 1978). As kidney is the most important hematogenic organ for fish (Zuasti and Ferrer, 1989), it is quite possible that effluent exposure resulted in hypofunction ofhemopoiesis, leading to anemia. Our histolopathological observations of kidney lesions in treated fish support this assumption. There were no prominent changes in red blood cell indices (MCV, MCH and MCHC) of catfish exposed to different effluent concentrations. Normocytic anaemia was evident at low and moderate doses while the high concentration resulted in microcytic anaemia. Furthermore, the results of the leucogram analysis suggest that initial leucocytosis was stress-related. As regards the differential count, effluent exposure at 6.25 and 12.5% concentrations induced lymphopenia. The reduction in percentage of lymphocytes contributes to immunosuppression of stressed fish. Thus antibody titer might be reduced as also observed by Saxena et al. (1992) in catfish (Heteropneustis fossilis) exposed to cadmium and hexachlorocyclohexane. The concurrent stimulation of neutrophils and monocytes might be a response to the destruction of phagocytic cells by contaminants such as metals (Witeska, 2001). In agreement with the present findings, Wojtaszek et al. (2002) reported profound lymphopenia and an increased number of circulating promyelocytes and myelocytes as well as metamyelocytes and mature polymorphonuclear neutrophilic granulocytesin Cyprinus carpio after a single intraperitoneal injection of a high dose of cortisol.
As the main organ of various key metabolic pathways, the effects of a chemical usually appear primarily in the liver. This in turn provides toxicologists with a definitive site for the investigation of the hepatotoxic potential of a chemical. The teleost liver is one of the most sensitive organs with regard to showing alterations in biochemistry, physiology and histoarchitecture following exposure to various types of environmental pollutants. In the present study, increases in GOT and GPT levels after 28 days of exposure to dyestuff and chemical effluent indicate liver damage, since increases in the activities of blood transaminase have been attributed to tissue damage, particularly the liver (Palanivelu et al., 2005). Sastry and Sharma (1980) observed an increase in GOT and GPT in the blood of Channa punctatus following the treatment of mercuric chloride. Similar results were also reported following chronic exposure of carbofuran (Ram and Singh, 1988). It is generally accepted that an increase of these enzyme activities in the extracellular fluid or plasma is a sensitive indicator of even minor cellular damage (Palanivelu et al., 2005). Thus, the measurement of transaminase activities in blood plasma of fish can be used as indicator for toxic exposure. In this context, The et al. (1997) and Pacheco and Santos (2002) have reported liver lesions in fish as evidence of the exposure to multiple environmental stressors. The liver lesions observed in our study were dose-dependent, with the highest effluent concentration inducing the highest damages. Similar results have been reported for Gambusia affinis and Oncorhynchus mykiss exposed to endosulfan (Cengiz et al., 2001; Capkin et al., 2006). Mosquitofish exposed to 1.0, 2.5 and 5.0 μg L-1 for 7, 14 and 30 days showed liver morphological alterations as cellular hypertrophy, sinusoids dilatation, cell vacuolization, disorganization of areas around blood vessels, hemorrhage, lymphocyte infiltration, cell disconnection and cellular degeneration (Cengiz et al., 2001) and trout exposed to 2.6 μg L-1 for 24 and 96 h showed liver edema and necroses (Capkin et al., 2006).
Another finding of this study was the increase of serum creatinine after exposure to the dyestuff and chemical wastewater indicating kidney failure. The kidney of fishes receives the largest proportion of postbranchial blood and therefore renal lesions might be expected to be good indicators of environmental pollution. In the present study, histological changes in the kidney after exposure to different concentrations of the effluent included shrinkage and damage of glomerulus. In addition, the tubular epithelium was desquamated, vacuolated and often destroyed. Dilation of the lumina of the kidney tubules, necrosis of tubules, shrinkage of glomerular tuft and vacuolation of blood cells in the glomerular tuft have been reported in Heteropneustes fossilis exposed to chlorpyrifos (Srivastava et al., 1990). Elsan treatment in Channa punctatus resulted in a significant decrease in the dimension of Bowman`s capsule and glomerulus and the tubules lost their regular shape due to precipitation of cytoplasm and karyolysis (Banerjee and Bhattacharya, 1994). Dass and Mukherjee (2000) reported dilation of tubules, necrotic changes characterized by karyorrhexis and karyolysis at the nuclei of affected cells of Labeo rohita exposed to hexachlorocyclohexane. Ortiz et al. (2003) showed tubular necrosis, desquamation and vacuolization of tubular epithelial cells in kidney of fishes exposed to lindane. Roy and Bhattacharya (2006) also reported similar results in fish exposed to arsenic.
At the end of experimental period, selected free amino acids were significantly lower in the treated groups exposed to dyestuff and chemical wastewater than the respective control group. The depletion of muscle free amino acid profiles in catfish concomitantly exposed to elevated concentrations of dyestuff and chemical effluent indicated use of free amino acids for energy purposes in stressed fish. Muscle free amino acid pool and especially branched chain amino acids, are used for oxidative metabolism and energy production (Milligan, 1997; Waagbø et al., 2008). Positive correlations between muscle free branched chain amino acids and plasma Na+ and negative correlation to plasma Cl-, further suggests that the increased energy may be used for osmoregulation (Waagbø et al., 2008).
Gills are generally considered good indicators of water quality, being used as models for studies of environment impact, such as xenobiotic agents (Fanta et al., 2003). In fish, gills are critical organs for their respiratory, osmoregulatory and excretory functions. Respiratory distress is one of the early symptoms of poisoning (McDonald, 1983). A high rate of absorption of water pollutants through gills also makes fish a vulnerable target of its toxicity (Srivastav et al., 1997). In this study, desquamation, necrosis, the lifting of the lamellar epithelium, oedema, aneurism, hyperplasia of epithelial cells and fusion of the secondary lamellae were observed in the gills after exposure to dyestuff and chemical wastewater. Several other studies have shown the results obtained from several contaminants. Karan et al. (1998) observed lesions such as epithelial hyperplasia and curling of secondary lamellae on the gills, swelling and thrombosis at the tips of several secondary lamellae and clup-shaped secondary lamellae. Erkmen et al. (2000) reported the lifting of epithelial layer from gill lamellae, necrosis and degeneration of secondary lamellae, shortening of secondary lamellae and clup-shaped lamellae in the gills of Lepistes reticulatus exposed to cyphenothrin. Aneurism, epithelial necrosis, secondary lamellae showing fusion, lifting of epithelium have also been showed in other species (Cengiz and Unlu, 2002, 2003). Ortiz et al. (2003) reported fusion of the secondary lamella, increased rising of the branchial epithelium and intraepithelial edema in gills after an accidental discharge of lindane.
The observed epithelial necrosis and desquamation of the gill epithelium are direct responses to the action of water pollutants. The defense responses noticed are lifting up of the epithelium, hyperplasia and lamellar fusion. The lifting up of the epithelium increases the distance through which the toxicant has to travel to reach the blood stream. Gill hyperplasia might serve as a defensive mechanism leading to a decrease in the respiratory surface and an increase in the toxicant-blood diffusion distance. The defense responses will take place at the expense of the respiratory efficiency of the gills and eventually, the respiratory impairment must outweigh any protective effect against pollution uptake. At 3.12% exposure to dyestuff and chemical effluent, the gills of C. lazera exhibited aneurism in many areas of secondary lamellae. This appearance of the secondary lamellae results from the collapse of the pillar cell system and breakdown of vascular integrity with a release of large quantities of blood that push the lamellar epithelium outward (Alazemi et al., 1996).
In conclusion, the present study indicates that untreated dye wastewater have severe toxic effects on fish. It is assumed that under field conditions, the complex dyestuff effluent could induce general toxic effects and possibly might disturb ecologically relevant processes such as fish reproduction. Present findings also demonstrate clearly the importance of proper treatment of textile dye wastewaters prior to discharge.
REFERENCES
- Alazemi, B.M., J.W. Lewis and E.B. Andrews, 1996. Gill damage in the freshwater fish Gnathonemus petersii (family: Mormyridae) exposed to selected pollutants: An ultrastructural study. Environ. Technol., 17: 225-238.
CrossRef - Alkindi, A.Y.A., J.A. Brown, C.P. Waring and J.E. Collins, 1996. Endocrine, osmoregulatory, respiratory and haematological parameters in flounder exposed to the water soluble fraction of crude oil. J. Fish. Biol., 49: 1291-1305.
CrossRef - APHA, AWWA and WEF., 1998. Standard Methods for the Examination of Water and Wastewater. 20th Edn., American Public Health Association/American Water Works Association/Water Environment Federation, Washington, DC., USA., ISBN-13: 9780875532356, Pages: 1220.
Direct Link - Barton, B.A., A.B. Rahn, G. Feist, H. Bolling and C.B. Schreck, 1998. Physiological stress response of the freshwater chondrostean paddlefish (Polyodon spathula) to acute physical disturbances. Comp. Biochem. Physiol. A, 120: 355-363.
CrossRef - Benguira, S. and A. Hontela, 2000. Adrenocorticotrophin and cyclic adenosine 3′,5′-monophosphate-stimulated cortisol secretion in interrenal tissue of rainbow trout exposed in vitro to DDT compounds. Environ. Toxicol. Chem., 19: 842-847.
CrossRef - Capkin, E., I. Altinok and S. Karahan, 2006. Water quality and fish size affect toxicity of endosulfan, an organochlorine pesticide, to rainbow trout. Chemosphere, 64: 1793-1800.
CrossRefPubMedDirect Link - Cazenave, J., D.A. Wunderlin, A.C. Hued and M. de los Angeles Bistoni, 2005. Haematological parameters in a neotropical fish, Corydoras paleatus (Jenyns, 1842) (Pisces, Callichthyidae), captured from pristine and polluted water. Hydrobiologia, 537: 25-33.
CrossRefDirect Link - Chapman, P.M., 2000. Whole effluent toxicity testing-usefulness, level of protection and risk assessment. Environ. Toxicol. Chem., 19: 3-13.
CrossRef - Cengiz, E.I. and E. Unlu, 2003. Histopathology of gills in mosquitofish. Gambusia affinis after long-term exposure to sublethal concentrations of malathion. J. Environ. Sci. Health B, 38: 581-589.
PubMed - Das, B.K. and S.C. Mukherjee, 2000. A histopathological study of carp (Labeo rohita) exposed to hexachlorocyclohexane. Veterinarski Arhiv., 70: 169-180.
Direct Link - Di Giulio, R.T., W.H. Benson, B.M. Sanders and P.A. Van Veld, 1995. Biochemical Mechanisms: Metabolism, Adaptation and Toxicity. In: Fundamentals of Aquatic Toxicology: Effects, Environmental Fate and Risk Assessment, Rand, G.M. (Ed.). 2nd Edn., Chapter 17, Taylor and Francis Inc., Boca Raton, FL., USA., ISBN-13: 978-1560320913, pp: 523-562.
- Engelhardt, F.R., M.P. Wong and M.E. Duey, 1981. Hydromineral balance and gill morphology in rainbow trout Salmo gairdneri, acclimated to fresh and sea water and affected by petroleum exposure. Aquat. Toxicol., 1: 175-186.
CrossRef - Erkmen, B., M. Caliskan and S.V. Yerli, 2000. Histopathological effects of cyphenothrin on the gills of Lebistes reticulatus. Vet. Hum. Toxicol., 42: 5-7.
PubMed - Fanta, E., F.S.A. Rios, S. Romao, A.C.C. Vianna and S. Freiberger, 2003. Histopathology of the fish Corydoras paleatus contaminated with sublethal levels of organophosphorus in water and food. Ecotoxicol. Environ. Saf., 54: 119-130.
CrossRefPubMedDirect Link - Gernhofer, M., M. Pawet, M. Schramm, E. Muller and R. Triebskorn, 2001. Ultrastructural biomarkers as tools to characterize the health status of fish in contaminated streams. J. Aquat. Ecosyst. Stress Recovery, 8: 241-260.
CrossRefDirect Link - Guerra, R., 2001. Ecotoxicological and chemical evaluation of phenolic compounds in industrial effluents. Chemosphere, 44: 1737-1747.
CrossRefDirect Link - Laurén, D.J. and D. Wails, 1990. Liver Structural Alterations Accompanying Chronic Toxicity in Fishes: Potential Biomarkers of Exposure. In: Biomarkers of Environmental Contamination, McCarthy, CRC Press, Boca Raton, ISBN: 9781351070263, pp: 17-57.
CrossRefDirect Link - Hontela, A., 1998. Interrenal dysfunction in fish from contaminated sites: In vivo and in vitro assessment. Environ. Toxicol. Chem., 17: 44-48.
CrossRef - Hontela, A., C. Daniel and A.C. Ricard, 1996. Effects of acute and subacute exposures to cadmium on the interrenal and thyroid function in rainbow trout, Oncorhynchus mykiss. Aquat. Toxicol., 35: 171-182.
CrossRef - Karan, V., S. Vitorovic, V. Tutundzic and V. Poleksic, 1998. Functional enzymes activity and gill histology of carp after copper sulfate exposure and recovery. Ecotoxicol. Environ. Saf., 40: 49-55.
CrossRef - Koprucu, S.S., K. Koprucu, M.S. Ural, U. Ispir and M. Pala, 2006. Acute toxicity of organophosphorous pesticide diazinon and its effects on behavior and some hematological parameters of fingerling European catfish (Silurus glanis L.). Pestic. Biochem. Phys., 86: 99-105.
CrossRefDirect Link - McDonald, D.G., 1983. The effects of H+ upon the gill of fresh water fish. Can. J. Zool., 61: 691-703.
CrossRef - Milligan, C.L., 1997. The role of cortisol in amino acid mobilization and metabolism following exhaustive exercise in rainbow trout (Oncorhynchus mykiss Walbaum). Fish Physiol. Biochem., 16: 119-128.
CrossRef - Munkittrick, L.R. and L.S. McCarthy, 1995. An integrated approach to ecosystem health: Top-down, bottom-up or middle out?. J. Aquat. Ecosyst. Health, 4: 77-90.
CrossRef - Nigam, P., I.M. Banat, D. Singh and R. Marchant, 1996. Microbial process for the decolorization of textile effluent containing azo, diazo and reactive dyes. Process Biochem., 31: 435-442.
CrossRefDirect Link - Ortiz, J.B., M.L.G. De Canales and C. Sarasquete, 2003. Histopathological changes induced by lindane (gamma-HCH) in various organs of fishes. Sci. Mar., 67: 53-61.
Direct Link - Pacheco, M. and M.A. Santos, 2002. Biotransformation, genotoxic and histopathological effects of environmental contaminants in European eel (Anguilla anguilla L.). Ecotoxicol. Environ. Saf., 53: 331-347.
CrossRefPubMedDirect Link - Palanivelu, V., K. Vijayavel, S. Ezhilarasibalasubramanian and M.P. Balasubramanian, 2005. Influence of insecticidal derivative (Cartap Hydrochloride) from the marine polychaete on certain enzyme systems of the freshwater fish Oreochromis mossambicus. J. Environ. Biol., 26: 191-196.
PubMed - Ram, R.N. and S.K. Singh, 1988. Carbofuran-induced histopathological and biochemical changes in liver of the teleost fish, Channa punctatus (bloch). Ecotoxicol. Environ. Saf., 16: 194-201.
CrossRefPubMedDirect Link - Reitman, S. and S. Frankel, 1957. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol., 28: 56-63.
CrossRefPubMedDirect Link - Schwaiger, J., R. Wanke, S. Adam, M. Pawert, W. Honnen and R. Triebskorn, 1997. The use of histopathological indicators to evaluate contaminant-related stress in fish. J. Aquat. Ecosyst. Stress Recovery, 6: 75-86.
CrossRefDirect Link - Sharma, S., S. Sharma and K.P. Sharma, 2006. Identification of a sensitive index during fish bioassay of an azo dye methyl red (untreated and treated). J. Environ. Biol., 27: 551-555.
PubMed - Srivastav, A.K., S.K. Srivastava and S.K. Srivastav, 1997. Impact of deltamethrin on serum calcium and inorganic phosphate of freshwater catfish, Heteropneustes fossilis. Bull. Environ. Contam. Toxicol., 59: 841-846.
CrossRefPubMedDirect Link - Teh, S.J., S.M. Adams and D.E. Hinton, 1997. Histopathologic biomarkers in feral freshwater fish populations exposed to different types of contaminant stress. Aquat. Toxicol., 37: 51-70.
CrossRefDirect Link - Tietz, N.W., 1995. Clinical Guide to Laboratory Tests. 3rd Edn., W.B. Saunders Company, Philadelphia, USA, ISBN: 072165035X, Pages: 1096.
Direct Link - van der Oost, R., J. Beyer and N.P.E. Vermeulen, 2003. Fish bioaccumulation and biomarkers in environmental risk assessment: A review. Environ. Toxicol. Pharmacol., 13: 57-149.
CrossRefPubMedDirect Link - Verma, S.R., A.K. Tyagi and R.C. Dalela, 2005. Toxicity of textile waste to some teleost fishes. Water Air Soil Pollut., 10: 351-357.
CrossRef - Waagbø, R., C.D. Hosfeld, S. Fivelstad, P.A. Olsvik and O. Breck, 2008. The impact of different water gas levels on cataract formation, muscle and lens free amino acids, and lens antioxidant enzymes and heat shock protein mRNA abundance in smolting Atlantic salmon, Salmo salar L. Comp. Biochem. Physiol. A, 149: 396-404.
CrossRefPubMed - Wepener, V., J.H.J. Van Vuren, F.B. Chatiza, Z. Mbizi, L. Slabbert and B. Masola, 2005. Active biomonitoring in freshwater environments: Early warning signals from biomarkers in assessing biological effects of diffuse sources of pollutants. Phys. Chem. Earth, 30: 751-761.
CrossRef - Wester, P.W. and J.H. Canton, 1991. The usefulness of histopathology in aquatic toxicity studies. Compa. Biochem. Physiol. Part C: Compa. Pharmacol., 100: 115-117.
CrossRefPubMedDirect Link - Witeska, M., 2001. Changes in the common carp blood cell picture after acute exposure to cadmium. Acta Zool. Lituanica, 11: 366-371.
Direct Link - Thophon, S., M. Kruatrachue, E.S. Upatham, P. Pokethitiyook, S. Sahaphong and S. Jaritkhuan, 2003. Histopathological alterations of white seabass, Lates calcarifer, in acute and subchronic cadmium exposure. Environ. Pollut., 121: 307-320.
CrossRefDirect Link