
Tewodros Mulualem
Jimma Agricultural Research Center, P.O. Box 192, Jimma, Ethiopia
Mohammed Abate
Afar Pastoralist Agricultural Research Institute, P.O. Box 16, Samara, Ethiopia
International Journal of Plant Breeding and Genetics
Year: 2016 | Volume: 10 | Issue: 2 | Page No.: 69-78







ABSTRACT
Heterosis is manifested as improved performance for F1 hybrids generated by crossing two inbred parents. The selection of parents for effective hybridization depends on the nature and magnitude of heterosis over mid parent, heterobeltiosis and economic heterosis present in genetic stocks. The response of heterosis either positive or negative mainly depends on the breeding objectives and the type of the crops used. In cereal and vegetable, positive and negative heterosis desired for yield and maturity. Moreover, the nature of pollination, the floral biology and the natural out crossing rate of in cereal and vegetable crops is important for heterosis. In some self pollinated vegetable crops (egg plant), heterosis became very large, with hybrid yield advantage ranges from 33-97%, due to, flowers are perfect and normally the natural out crossing rate near zero. However, most of these crops, produce few seeds per cross make it difficult to produce hybrids economically. Most cereal crops are cross pollinated, however, the natural out crossing rate of 25% and heterosis yield of 28-47%. Nevertheless, self incompatible inbred lines are common in some cross pollinated vegetable and inbreeds can be self pollinated by bud pollination. In vegetable crops, heterosis developed in to four ways, self pollinated with few seed per cross, self pollinated with many seed per cross, cross pollinated with little out crossing and cross pollinated with much out crossing. Exploitation of heterosis in cereal and vegetable provides enhancing food security and represents a greatest applied achievement in the discipline of plant breeding.
PDF Abstract XML References Citation
Received: January 22, 2016;
Accepted: February 24, 2016;
Published: March 15, 2016
How to cite this article
Tewodros Mulualem and Mohammed Abate, 2016. Heterotic Response in Major Cereals and Vegetable Crops. International Journal of Plant Breeding and Genetics, 10: 69-78.
DOI: 10.3923/ijpbg.2016.69.78
URL: https://scialert.net/abstract/?doi=ijpbg.2016.69.78
DOI: 10.3923/ijpbg.2016.69.78
URL: https://scialert.net/abstract/?doi=ijpbg.2016.69.78
INTRODUCTION
Heterosis is the hybrid vigor manifested in hybrids and represents the superiority in performance of hybrid individuals compared with their parents. Hybrid vigor in crops is manifested in the offspring of inbred lines with high Specific Combining Ability (SCA). Heterosis was first applied by the purposed hybridization of complex hybrid mixtures made by farmers1 in the 1800s. However, public scientists E.M. East and G.H. Shull developed the concept of hybrid vigor or heterosis in maize independently2,3 in the early 1900s. It was realized that genetic divergence of parental crosses was important for hybrid vigor expression in cereal crops4. The range of genetic divergence limited the expression of heterosis5. Heterosis can be inferred from heterotic patterns6. A heterotic pattern is the cross between known cross pollinated cereal genotypes that expresses a high level of heterosis7 and it established by relating the heterosis of crosses with the origin of the parents included in the crosses8. Although, many vegetable crops have significant heterosis, some of the major benefits of hybrids relate to other issues. A primary benefits is the protection of parental inbreeds used in the production of elite hybrids. This has become more important with increased involvement of private companies in the development of vegetable cultivars. Hybrids can be sold in commerce without making the parental inbred lines available, so the parents are protected under the ''Trade secrete'' provision of the laws seed companies. A second important benefit of hybrids is uniformity of traits expression among plants of a cultivar. Uniformity was an especially attractive selling point in broccoli and cabbage when they became available as alternatives to open pollinated cultivars. Uniformity of harvest provided earlier and higher yield and uniformity of the harvested product resulted in higher price per kilogram. Hybrids offer additional advantages in vegetable crops where important traits are controlled by dominant genes. Hybrids make it easier to produce a cultivar with many useful traits by combining dominant alleles from the two parents9,10.
Vegetable can grouped according to how adaptable they are to hybrid production. Based on this, self pollinated crops with few or many seeds produced per cross and cross pollinated crops with a low or high rate of natural out crossing. Important self pollinated vegetable crops that have been researched extensively include Solanacea family. Important cross pollinated vegetable crops include the Cucurbitacea, cole crops and root and bulb crops. The wide range of variability for different crop characters indicates their possible utility in a hybrid development program for vegetable crops11.
Once the heterotic patterns are established in cultivated crops, the identification of best performing hybrids is cost-efficient. Moreover, the performance of crosses (in the first-generation) among groups based upon genetically diverse parents is essential to identify promising heterotic patterns12. Consequently, the establishments of heterotic patterns and comparison study on heterotic response among different crops have tremendous impact for selecting inbred lines as potential seed stocks in hybrids13. Therefore, this paper was assessed on the comparative heterotic responses of the most cultivated cereals and vegetable crops under different heterotic breeding.
INBREEDING AND HETEROSIS
Inbreeding (genetic assortive mating), a converse phenomenon of heterosis, is usually defined as the lowered fitness or vigour of inbred individuals compared with their non-inbred counterparts. It is the most powerful of all mating systems in all self pollinated crops to lower the percentage of heterozygosity in the population leading to fixation of alleles and thus the phenotype to the extent that is under genetic control12. Precisely assortive mating or inbreeding reduces the proportion of heterozygous loci by half in each generation and homozygous types are correspondingly increased. In quantitative genetic theory, inbreeding depression and heterosis are due to non-additive gene action and are considered to be two aspects of the same phenomenon. Thus, the most striking observed consequence of inbreeding is the reducing of mean phenotypic value and the phenomenum known as inbreeding depression13.
Heterosis and inbreeding depression indeed are two faces of the same coin and reflect two diametrically opposite action of the same genetic phenomena. Inbreeding depression is likely to be manifested practically for all the characters displaying the heterosis. Inbreeding depression usually is associated with reduction in vigor, fertility and productivity. The progeny includes very weak individuals which are difficult to be maintained14. The effect of inbreeding is very pronounced in first 5-8 generations after, which the general level of performance stabilizes. The homozygosity increases and lines became different from each other but, differences among individuals with in lines are reduced. Like that of heterosis the extent of inbreeding depression also varies with crops as well as the characters. Some crops are completely intolerant to inbreeding alfalfa and carrot, which deteriorate drastically in first few generation of inbreeding.
The inbreeding progeny of such crops includes very weak and lethal individuals, which die even at the seedling stages. Inbreeding depression is quite high in maize where, plants became extremely weak and less productive after 4-5 generation of inbreeding depression. The modern cultivated varieties of cereals particularly in maize however, may not show very high level of depression13. The inbreeding depression is quite low in crops like onion sunflower, rye pear millet, cucurbits and hemp. In general high breeding depression is displayed by cross pollinated crops, whereas self pollinated crops are highly tolerant to inbreeding practically with respect to survival of individuals. This differential behavior of different crops can again be related to type of genetic balance. The heterozygous balance is cross pollinated crops lead to reduce the fitness. The in breeders, on the other hand, have higher 'Homozygous balance' that adapts these species such that optimum fitness emanates from heterozygotes.
In quantitative traits, inbreeding depression is quantified by the reduction in the mean that occurs only under dominant genetic effects, in other words, for traits with no dominance, no inbreeding depression is detected. The deleterious recessive major genes with qualitative inheritance may not only cause strong inbreeding depression but may also act as epistatic genes in that they can block important physiological pathways, precluding the expression of other quantitative genes of minor effects14. The increase of homozygosity or the decrease in the frequency of heterozygous genotypes is a natural consequence of inbreeding and reaches its maximum rate through selfing, the most extreme form of inbreeding. Other less severe forms of inbreeding also lead to increases in homozygosity whose rate depends on the degree of parentage between the mating parents. The inbreeding coefficient (F) is the probability of randomly uniting gametes having alleles identical by descent. So F is the probability of homozygosis where the homozygotes carry alleles that are identical copies of the same allele in a common ancestor15.
Heterosis depends on dominance and dominance types of epistatic genetic effects; in this sense no heterosis can be detected for a quantitative trait in a hybrid, in relation to the average of the parents, if genes controlling the trait act in a strictly additive way (no dominance). The phenomenon of heterosis is the opposite of the inbreeding depression in the sense that the vigor lost during inbreeding is recovered through crossing. However, although inbreeding depression and heterosis depend on dominance genetic effects, they are not merely the same phenomenon in opposite directions. In fact, when dealing with populations the inbreeding depression is an intrapopulation effect, while heterosis is expressed at the inter population level. Earlier studies on heterosis defined the phenomenon as merely the recovery of inbreeding depression caused by major genes visually detected. However, under a quantitative viewpoint, heterosis may occur whenever there is genetic divergence (differences in gene frequencies) between parents and some level of dominance controlling the trait. The recovery of vigor lost as a consequence of the effects of deleterious genes of large effects can be seen as a nullification of the epistatic effects (preventing full expression of other genes of minor effects) of the recessive major genes. If dominance controls a trait, heterosis will be a function of the distribution of allele frequencies.
PRAGMATIC EVIDENCE OF HETEROSIS
Heterosis can be grouped into two broad categories, true heterosis and pseudo-heterosis. In case of true heterosis, there is an increase in general vigor, yield and adaptation. In case of pseudo-heterosis, the F1 hybrid exhibits increase in vegetative growth only. It refers to the superiority of F1 over the standard commercial check variety, it is also called economic heterosis or superiority over checks16. Recently it has been divulged that the utilization of heterosis is extremely effective for the genetic improvement of different traits and that the concepts of combining ability are the fundamental tools for enhancing hybrids17. Heterosis occurred in the F1 productivity of different crops in the form of F1 hybrids for all traits of interest, but their values varied among crosses and characters. However, genotypes harboring desirable attributes and stable performance are vital, both as cultivars and as source of desirable germplasm for further improvement. The concept of heterosis is wide and different criteria are being adopted for the evaluation of hybrid vigour. Among the methods of evaluation of hybrid vigour, the following three aspects of comparative evaluations are worth considering.
Mid-parent heterosis (MPH): It is the performance of a hybrid relative to the average performance of its parents expressed in percentage:
![]()
where, mp is mid-parent value obtained as (P1+P2)/2.
High-parent heterosis (HPH): It is the performance of a hybrid relative to the performance of its best parent expressed in percentage:
![]()
Heterosis over check (HCH):
![]()
The high parent heterosis method has been less used but it provides better and more accurate information.
GENETIC MODELS OF HETEROSIS
Heterosis can be controlled by multiple linked or unlinked genes, represented by ‘A’, ‘B’, etc. These types of genes are described by dominance, over-dominance and pseudo-over dominance genetic model of heterosis.
Dominance: This model proposes that independent sets of slightly deleterious alleles by ‘a’ in parent 1 (P1) and ‘b’ in P2 accumulate throughout the genome during inbreeding of parental lines (inbreeding depression). Hybridization causes complementation in F1 progeny by the superior alleles ‘A’ in P2 and ‘B’ in P1, resulting in phenotypic superiority over both parents. Dominance is most easily conceptualized in the context of monogenic recessive mutations that affect plant fertility. For example, heterosis can be artificially created by crossing two genotypes carrying mutations in different genes affecting fertility. Reciprocal complementation following hybridization fully restores fertility leading to superiority over the two parents.
Over-dominance: This model proposes that allelic interactions at a single heterozygous locus result in a synergistic effect on vigor that surpasses both homozygous parents17,18. In the example shown, P1 contributes allele ‘B* and P2 allele ‘B’, with no assumption of a dominant-recessive relationship18. The F1 hybrid carries both alleles, which function together to promote greater vigor. Although, evidence for over-dominance is debated, examples of over-dominance genes are known. Plants heterozygous show greater fertility than homozygous type siblings, but no molecular explanation is known19.
Pseudo-over-dominance: The genetic intermediate of dominance and over-dominance is ‘Pseudo-over-dominance’, which is actually a simple case of dominance complementation, in which the two recessive mutations (‘a’ from P1 and ‘b’ from P2) are linked in trans, or ‘In repulsion’.
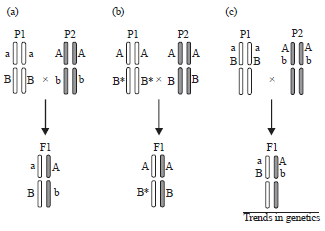 | |
| Fig. 1(a-c): | Genetic models of heterosis20, (a) Dominance, (b) Overdominance and (c) Pseudo-overdominance |
This type of complementation in the hybrid resembles over-dominance, because of the tight chromosomal linkage (Fig. 1).
COMPARATIVE HETEROTIC RESPONSE IN CEREAL AND VEGETABLE OF CROPS
The use of heterosis has become one of the major strategies for increasing crop production. This review focused the most important cultivated cereal and vegetable crops have contributed a great deal towards increasing the yield potential.
HETEROTIC RESPONSE IN CEREALS
Heterosis results in the phenotypic superiority of a hybrid over its parents with respect to traits, such as growth rate, reproductive success and yield. This hybrid vigor is determined by non-mutually exclusive mechanisms, including dominance complementation, over-dominance and epistasis in cereals crop. Its magnitude depends on the choice of appropriate parental lines. To increase the efficiency of heterosis breeding in cross pollinated crops, there is a need for a simple and dependable criterion, to select the parental lines which would result in heterotic combinations without making all possible crosses among the potential parents and evaluating them. Several methods (per se performance, combining ability, genetic diversity, etc.) have been used to choose parents a priori with variable success21. However, most traits of economic importance are qualitative and controlled by many major genes. This study assessed the heterotic response in major economical important cereal crops as follow:
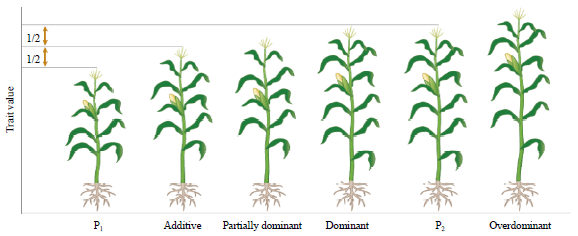 | |
| Fig. 2: | Phenotypic level of trait in a hybrid can be described using several terms in maize20 |
Heterosis in maize: Heterosis is important in maize breeding and depends up on the level of dominance and diversity in gene frequencies. The manifestation of heterosis depends on genetic divergence of two parental varieties8,22. Heterosis is the phenomenon, wherein the performance of an F1 derived by crossing two genetically different individuals is superior to that of the mean of the parents or the better parent. Maize has great potential for heterotic manifestation and its exploitation. This could be the reason that number of hybrid varieties in maize is much higher than any other varietal types i.e., open pollinated, double cross, synthetics or three way crosses. It is endowed with significant amounts of heterosis for grain yield and other agronomic traits. With the introduction of heterosis concepts in maize there has been a breakthrough in yield of this crop. Hence, it may be viewed that there is a wider scope for further improvement of yield through appropriate genetic manipulations. One of the top priorities in breeding of this crop is to exploit the heterosis partially or fully by developing synthetics/composites or hybrids22.
The performance of hybrid depends on the genetic makeup of the parents used. Therefore, selection of appropriate parents is the first step in maize hybrid breeding program. Based on inbred lines abilities to produce superior hybrids, maize parental lines have been grouped into heterotic groups. Further, certain cross combinations produce desirable off springs, whereas, other involving equally promising parents produce poor progeny12. Therefore, selection should be based on the sound consideration of combining ability of parents is the most desirable and high yielding cross combinations could be combed out from a large number of combinations. The estimate of heterosis determined by heterosis over mid parent (average heterosis), better parent (heterobeltiosis) and best check hybrid (standard heterosis) for all the characters. The scope for exploitation of hybrid vigour is dependent upon the magnitude of heterosis, biological feasibility, genetic divergence and the type of gene action i.e., additive, dominance, additive×additive, dominance×dominance effect, which are the components that determines heterosis (Fig. 2).
Recently, it has been divulged that the utilization of heterosis is extremely effective for the genetic improvement of different traits and that the concepts of combining ability are the fundamental tools for enhancing productivity of different cereal crops in the form of F1 hybrids17. Heterosis occurred in the F1 hybrids for all traits of interest, but their values varied among crosses and characters. However, genotypes harboring desirable attributes and stable performance are vital, both as cultivars and as source of desirable germplasm for further improvement.
Heterotic response in wheat: Grain yield is clearly the economic trait of interest in hybrid wheat and the desired expression of economic heterosis can be achieved by matching yield and yield components from genetically diverse parents23. Based on Cytoplasmic Male Sterility (CMS), Nuclear Male Sterility (NMS) as the sterility system for commercial hybrid production and reported maximum better parent heterosis ranging from 3-41% and standard heterosis, the comparison of a hybrid to a commercial cultivar, ranging from -4 to 32%. In a compilation of studies investigating several yield components, grain number per spike, spikes per plant, 1000 grain weight, harvest index, dwarf plant height and spike length were more frequently associated with the significant heterosis for grain yield, suggesting that there could be no separate gene system for yield per se as yield is an end product of the multiplicative interactions between its various component characters24. This indicated that, on the average, a gene causing an increase in one character will also result in an increase in the other character or vise versa. So selecting for one character will help in selecting for the other character24.
Although, several yield components appeared to be important in determining grain yields in high yielding hybrids, the crosses showing heterosis for grain yield per plant were not heterotic for all the characters. Hybrid vigor of even small magnitude for individual yield components may have additive or synergetic effects on the end product25. Evidently, manifestation of heterosis for yield and its attributes may be due to non-additive gene effect in the parents. It was observed that high yielding pure lines did not produce high heterotic response frequently in number of crosses for grain yield24. The first major advantage of hybrid wheat is the consistency of yield and higher yield performance. The consistency of performance of hybrids and comparable conventional varieties tested across range of environments showed that hybrids were more consistent in performance than varieties and cope much better in harsh environments26.
Heterotic response in rice: The utilization of population improvement methods in rice only became feasible after the discovery of the male sterile gene through induced mutation of the rice. The initial breeding strategy to produce hybrids relied on three breeding lines known as A line (the male sterile line), B line (responsible to maintain the genetic male sterility of the A line) and R line (used to restore the fertility of the A line and to produce the hybrid seed). The ideal system for these and other cross-pollinated crops would be the one-line method utilizing the apomixis system that allows preserving the right cultivar. Identi cation and evaluation of male-sterile lines and their restorers, test cross phase to select heterotic combinations and to initiate conversion of maintainer lines into male-sterile lines, back crosses to transfer the cytoplasmic male-sterility to elite maintainer lines, trials to study the combining ability (general and specific) of the parental lines and foundation seed production of all three lines. Production of breeding lines for the three- or two-line methods is still a difficult task for most of the rice breeding programs26.
Heterosis is the base of the great success in hybrid rice. Currently, hybrid rice accounts for 55% of the total planting acreage of paddy rice in Far East Asia and the annual increased rice production resulting from planting hybrid rice amounts to 20 million metric tons, which can provide a main staple food for 70 million people27. Hybrid rice varieties have a yield advantage of 10-20% over the best conventional inbred varieties using similar cultivation conditions27. In rice breeding, most agronomic and grain quality traits are controlled by many genes each of, which has a relatively small effect on the overall phenotype. These traits do not show discrete phenotypes, consequently they are often measured and given a quantitative value and are referred to as quantitative traits. Quantitative traits are difficult to study because the phenotypes do not give an insight into the genotype. The expression of genes controlling quantitative traits can be greatly influenced by the environment. Consequently, the improvement of polygenic traits by traditional breeding methods is time consuming and the gains are harder to realize. Breeders usually overcome this problem by multi environmental evaluation of replicated trials to capture the effect of the environment.
Due to the intractable problem encountered while trying to improve quantitative traits by conventional means, breeders and geneticists have considered the potential use of DNA markers to identify chromosomal regions harboring genes that influence the quantitative traits. Morphological markers were the first generation of markers to be used for identification and selection for quantitative traits loci. Very slow progress was made in the use of these markers due to the undesirable effects of many of the markers on the target phenotype. The effect of the marker genes on the quantitative traits was often larger than that of the linked Quantitative Trait Loci (QTL), hence it was difficult to effectively and extensively use these markers to study quantitatively inherited traits28. Moreover, there was a dearth of available segregating genetic markers26.
Several efforts have been made to introgress useful genes into elite rice varieties through interspecific hybridization with varying amount of success. Recent breakthroughs in another culture and molecular biology provide greater opportunities for rice breeders to develop a new generation of rice varieties that are better adapted and high yielding. It is now possible to select for molecular markers linked to traits of interest (marker assisted selection) rather than selecting for the traits themselves. Chromosomal regions controlling various agronomic traits have been identified in rice. The QTL have been revealed through their association with molecular markers and by the year 2000, more than 1000 QTL have been documented in rice29. However, the use of these QTLs by breeding programs is still in its infancy. Only a few of the reported QTLs have been used for crop improvement.
HETEROSIS IN VEGETABLE CROPS
Some of the earliest study on heterosis was in the vegetable crops. The discovery cytoplasmic male sterility in onion, is the base for making hybrid production for economical. Growers adopted hybrid cultivars rapidly when they were made available by the seed industry. However, hybrid cultivars have not been exploited commercially as much in self-pollinated vegetable crops as in cross-pollinated ones. Major self-pollinated vegetable crops include the Solanaceae (egg plant, Solanum melongena, pepper, Capsicum annuum; tomato, Lycopersicon esculentum) and lettuce (Lactuca sativa). Cross-pollinated vegetable crops include the cucurbits (cucumber, Cucumis sativus, melon, Cuwmis meld, squash, Cucurbita spp., watermelon, Citrullus lanatus), the cole crops (broccoli, cabbage, cauliflower, Brassica oleracea), root and bulb crops (carrot, Daucus carota, onion, Allium cepa), asparagus (Asparagus officianalis) and spinach (Spinacia oleracea).
The advantages of hybrid cultivars in the vegetable crops include expression of heterosis for yield and other traits, protection of proprietary parental lines, combined expression of dominant alleles from the parents and production of unique traits from the parents. In this study, the vegetable crops classified in to four groups according to their use of heterosis: (1) Self-pollinated with few seeds per cross, (2) Self- pollinated with many seeds per cross, (3) Cross-pollinated with little out crossing and (4) Cross-pollinated with much out crossing.
SELF-POLLINATED WITH FEW SEEDS PER CROSS
In some of the self-pollinated vegetable crops, such as bean, pea and lettuce, there has been no use of heterosis or hybrid cultivars. In those crops, self-pollination occurs naturally, cultivars are inbred lines, there is no inbreeding depression, cross-pollination is difficult (other than controlled crosses in a breeding program), little heterosis is expressed for yield and no economical mechanism exists for mass production of hybrid seeds. In this type of plants, hybrids are difficult to produce, because of perfect flowers that enclose the stamens and style. In bean, efforts have been unsuccessful to incorporate the extrorsely stigma (stigma facing away from pollen) trait from the (naturally outcrossing) scarlet runner bean (Phaseolus coccineus). Extrorse stigma might provide a way to make hybrids economically to protect proprietary inbred lines developed by seed companies. Opportunities for hybrid production are even worse in lettuce, where only 10-20 seeds are produced per pollination and 250,000 seeds per hectare are required to plant the crop. The pollen is sticky and is not carried by insects or wind. Also, some of the important traits (lettuce mosaic and corky root rot) are controlled by recessive genes, making it necessary to incorporate them into both parents of the hybrid.
SELF-POLLINATED WITH MANY SEEDS PER CROSS
In the Solanaceae, important crops, such as eggplant and tomato have been easily adapted to hybrid production. In those crops, hand pollination is easy, many seeds are produced from each cross and significant heterosis is expressed for important traits like yield (estimates average 60% for egg plant and tomato and 35% for pepper). In those crops, hybrid cultivars are being produced widely.
CROSS POLLINATED WITH LITTLE OUT CROSSING
In the Cucurbitaceae, crops, such as cucumber, melon, squash and watermelon have been adapted to hybrid production. In these crops, cross-pollination is easily enforced, often there are methods for mass production of hybrid seeds, many seeds are produced from each cross and heterosis is usually expressed for important traits like yield. In cucumber, there is no inbreeding depression and little heterosis. However, hybrids are used to provide gynoecious sex expression and for protection of proprietary inbreds. Hybrids are made by crossing gynoecious with monoecious inbreds in isolation using bees as pollinators. A major advantage of this system is that the hybrid is gynoecious, producing more pistillate nodes than normal cultivars. The extra pistillate nodes give gynoecious cultivars and advantage in early yield and are useful for once-over harvest production systems. Gynoecious hybrids have been produced in melon, but inheritance of sex expression is different than in cucumber. It has been difficult to stabilize sex expression in melon for reliable production of hybrids. Also, pistillate flowers produce oval fruits and the market demands the round fruits produced by perfect flowers. Since, there is only 8% heterosis and no inbreeding depression and because of the difficulties with hybrid production, most cultivars are and romonoecious inbreds. Summer squash (Cucurbita pepo) hybrids are produced using two monoecious inbreds. One is converted to gynoecious sex expression by spraying it with ether at the 2 and 4 leaf stages. That system does not work well for winter squash, so most cultivars are open-pollinated. Watermelon hybrids are made by hand pollination, although male sterility is being researched for possible reduction in labor requirement. Hybrids provide greater uniformity in crop production than open-pollinated cultivars and protect proprietary parental inbreds. However, much of the industry is now using hybrids between tetraploid and diploid inbreds as a method for producing seedless (triploid) cultivars.
CROSS POLLINATED WITH MUCH OUT CROSSING
The cole crops (broccoli, cabbage and cauliflower), root and bulb crops (carrot, onion) and other crops (asparagus, spinach) in this group have been adapted to hybrid production using diverse methods. In these crops, pollination control is difficult, but methods have been developed for mass production of hybrid seeds. However, hybrids are often earlier and more uniform than open-pollinated cultivars and heterosis is expressed for important traits like yield. Heterosis of 12-15% occurs in broccoli, cabbage and cauliflower and they express some inbreeding depression. Those crops have sporophytic self-incompatibility to enforce outcrossing, but bud pollination can be used to self-pollinate plants. Hybrids are produced by planting two self-incompatible inbreds in isolation. Seeds harvested from the two lines are mostly hybrid. Hybrids were adopted widely by growers, because of increased uniformity and earliness, permitting fewer harvests of the crop. Unfortunately, self-incompatibility does not always prevent self-pollination, so the parent plants can be found in production fields. Because of that, breeders have begun using cytoplasmic male sterility for hybrid production.
COLE CROPS
Broccoli, cabbage and cauliflower are hermaphroditic annals with a high degree of out crossing enforced by sporophytic self incompatibility. Significant inbreeding depression and heterosis is present. The exception is summer cauliflower where there is no inbreeding depression and no self incompatibility. Heterosis of hybrids over open pollinated cultivars may be only 10%. Cabbage hybrids were 12-15% better yielding than open pollinated cultivars30. In broccoli, hybrid cultivars yielded 40-90% better than open pollinated ones, but that comparison was no entirely due to heterosis.
High rate of natural out crossing and heterosis in the cole crops has made them ideal for hybrid production, with 84% of cabbage and 100% of broccoli and in hybrid cultivars. Another significant factor in the adoption of hybrids has been their greater uniformity in yield and quality relative to open pollinated cultivars. Single cross hybrids are produced by crossing two self incompatible inbred lines in an isolation block. Self incompatible inbreeds can be self pollinated using bud pollination, since flowers are self compatible before they open. The system permits economical production of hybrid seed, but it ''Leaky''. Occasional self or sib-pollination results in seeds of the parental inbred being sold in the hybrid cultivar. One solution has been to use four way hybrids (with occasional two way hybrids mixed in). Cytoplasmic male sterility is being phased in for hybrid production since it is more reliable in protecting the proprietary inbred lines used as parents.
ROOT AND BULB CROPS
Beet and carrot express much inbreeding depression, so lines usually are not advanced beyond S5. When cytoplasmic male sterility was discovered, hybrids began to replace open-pollinated cultivars. Hybrids are produced using bees in isolation to move pollen from normal to male sterile inbred lines. Onion was one of the first crops to make use of cytoplasmic male sterility for hybrid production. Onion hybrids are 14-67% better than open-pollinated cultivars and are widely used in onion producing country. Onion is a hermaphroditic biennial outcrosser. There is significant breeding depression, making the development of inbred lines difficult. Heterosis is large, with hybrids 14-67% higher yielding than the best open pollinated cultivars. Hybrid production was made economically by the discovery of cytoplasmic male sterility.
Carrot (Daucus carota L.) is a monocious biennial outcrosser with much inbreeding depression and essentially no naturally self pollination. Heterosis for marketable yield was measured at 25-30% over open pollinated cultivars. Most cultivars are two or three way hybrid and are produced by cytoplasmic malesterelity. Inbreeds are difficult to develope because of inbreeding depression, but can be advanced to S3-S6 by alternating each generations of self pollination with open pollination. Hybrids are produced in isolation blocks with eight rows of the female parent alternating with two rows of the male parent. Hybrids are produced on 50% of the carrot land.
Cultivated okra is an introduced vegetable crop. Although, it is a multipurpose and multifarious crop, it is extensively grown for its tender pods, which are used as a very popular, tasty and gelatinous vegetable. It is a power house of valuable nutrients. Despite its recognized potential and significant area and consumption, it is being neglected because of non-availability of high yielding open-pollinated varieties. Yield plateau seems to have been reached in open-pollinated varieties of okra. To break the yield barriers in existing open-pollinated varieties of okra, a hybridization-based breeding strategy would be desirable. Heterosis breeding has been the most successful approach in increasing the productivity in cross-pollinated vegetable crops.
Okra is one often-cross pollinated vegetable crop. The ease in emasculation and very high percentage of fruit setting indicates the possibilities of exploitation of hybrid vigour in okra. The presence of sufficient hybrid vigour is an important prerequisite for successful production of hybrid varieties. Therefore, the heterotic studies can provide the basis for the exploitation of valuable hybrid combinations in the future breeding programs and their commercial utilization. The initial selection of parents for effective hybridization depends upon the nature and magnitude of relative heterosis (heterosis over mid parent), heterobeltiosis (heterosis over better parent) and economic heterosis (heterosis over check) present in genetic stocks. Exploitation of heterosis is primarily dependent on the screening and selection of available germplasm that could be produced by better combinations of important agronomical and horticultural traits. Heterosis breeding based on the identification of the parents and their cross combinations is capable of producing the highest level of transgressive segregates31. The choice of the best parental matings is crucial for the development of superior hybrids and because combinations of hybrids grow exponentially with the potential number of parents to be used, this is one of the most expensive and time-consuming steps in hybrid development programs32. The magnitude of heterosis provides a guide for the choice of desirable parents for developing superior F1 hybrids, so as to exploit hybrid vigour. It also helps in choosing suitable crosses to be used for commercial exploitation as well as in component breeding program.
OTHER VEGETABLE CROPS
Asparagus is a dioecious perennial outcrosser having a single gene for control of sex expression, with male (Mm) plants producing higher yield than female (mm) plants, because they do not use photosynthate in seed production each year33. Since, male plants occasionally have perfect flowers, it is possible to self-pollinate males to do progeny testing to develop super males (MM). Hybrids produced by crossing a female line with a super male line will be all male. Males yield 25-38% more than females33. Also, hybrids can yield 64-149% more than open pollinated cultivars.
Spanach (Spinacea pleracea L.) is normally dioecious, but monocious flowering also exists. It is an annual outcrosser that is wind pollinated with a natural outcrossing rate of 60% (near 100% in dioecious population). Inbreeding deppression occurs with a 2-6% reduction in yield relative to open pollinated cultivars. Hybrid cultvars out yield open pollinated cultivars by 16-20%. Hybrids are produced by planting a monocious inbred with a high percentage of pistillate flowers (gynoecious inbred) aternating with rows of a monoecious or androecious inbred. The gynoecious inbred is maintained in an isolation block, using the late-appearing staminate flowers for natural self pollination.
CONCLUSION
Hybrid cultivars have been adopted rapidly since their introduction by occupying 100% of production area.
ACKNOWLEDGMENT
We greatly acknowledge to Dr. Wasu Mohammed, Haramaya University for sharing his deepest knowledge and skills in the area of heterosis breeding.
REFERENCES
- Anderson, E. and W.L. Brown, 1952. The history of the common maize varieties of the United States corn belt. Agric. History, 26: 2-8.
Direct Link - Moll, R.H., J.H. Lonnquist, J.V. Fortuno and E.C. Johnson, 1965. The relationship of heterosis and genetic divergence in maize. Genetics, 52: 139-144.
Direct Link - Carena, M.J., 2005. Maize commercial hybrids compared to improved population hybrids for grain yield and agronomic performance. Euphytica, 141: 201-208.
CrossRefDirect Link - Stuber, C.W., S.E. Lincoln, D.W. Wolff, T. Helentjaris and E.S. Lander, 1992. Identification of genetic factors contributing to heterosis in a hybrid from two elite maize inbred lines using molecular markers. Genetics, 132: 823-839.
PubMedDirect Link - Melchinger, A.E., J. Boppenmaier, B.S. Dhillon, W.G. Pollmer and R.G. Herrmann, 1992. Genetic diversity for RFLPs in European maize inbreds. II. Relation to performance of hybrids within versus between heterotic groups for forage traits. Theoret. Applied Genet., 84: 672-681.
CrossRefPubMedDirect Link - Falconer, D.S. and T.F.C. Mackay, 1996. Introduction to Quantitative Genetics. 4th Edn., Prentice Hall, Harlow, England, ISBN-13: 9780582243026, Pages: 464.
Direct Link - Allard, R.W., 1960. Principles of Plant Breeding. 1st Edn., John Wiley and Sons Inc., New York pp: 372-372.
CrossRefDirect Link - Charlesworth, D. and J.H. Willis, 2009. The genetics of inbreeding depression. Nat. Rev. Genet., 10: 783-796.
CrossRefDirect Link - Lippman, Z.B. and D. Zamir, 2007. Heterosis: Revisiting the magic. Trends Genet., 23: 60-66.
CrossRefDirect Link - Lu, H., J. Romero-Severson and R. Bernardo, 2003. Genetic basis of heterosis explored by simple sequence repeat markers in a random-mated maize population. Theor. Applied Genet., 107: 494-502.
CrossRefPubMedDirect Link - Sharief, A.E., S.E. El-Kalla, H.E. Gado and H.A.E. Abo-Yousef, 2009. Heterosis in yellow maize. Aust. J. Crop Sci., 3: 146-154.
Direct Link - Flint-Garcia, S.A., E.S. Buckler, P. Tiffin, E. Ersoz and N.M. Springer, 2009. Heterosis is prevalent for multiple traits in diverse maize germplasm. PLoS One, Vol. 4.
Direct Link - Luo, L.J., Z.K. Li, H.W. Mei, Q.Y. Shu and R. Tabien et al., 2001. Overdominant epistatic loci are the primary genetic basis of inbreeding depression and heterosis in rice. II. Grain yield components. Genetics, 158: 1755-1771.
Direct Link - Shull, G.H., 1908. The composition of a field of maize. Rep. Am. Breeders Assoc., 4: 296-301.
CrossRefDirect Link - Frascaroli, E., M.A. Cane, P. Landi, G. Pea and L. Gianfranceschi et al., 2007. Classical genetic and quantitative trait loci analyses of heterosis in a maize hybrid between two elite inbred lines. Genetics, 176: 625-644.
CrossRefPubMedDirect Link - Singh, S.K., R. Chatrath and B. Mishra, 2010. Perspective of hybrid wheat research: A review. Indian J. Agric. Sci., 80: 1013-1027.
Direct Link - Burke, J.M. and M.L. Arnold, 2001. Genetics and the fitness of hybrids. Annu. Rev. Genet., 35: 31-52.
CrossRefDirect Link - Duvick, D.N., 1999. Heterosis: Feeding People and Protecting Natural Resources. In: The Genetics and Exploitation of Heterosis in Crops, Coors, J.G. and S. Pandey (Eds.)., American Society of Agronomy Inc., Madison, Wl. USA., pp: 19-29.
Direct Link - Tanksley, S.D. and J.C. Nelson, 1996. Advanced backcross QTL analysis: A method for the simultaneous discovery and transfer of valuable QTL from unadapted germplasm into elite breeding lines. Theor. Applied Genet., 92: 191-203.
CrossRefDirect Link - Xu, Y., H. Beachell and S.R. McCouch, 2004. A marker-based approach to broadening the genetic base of rice in the USA. Crop Sci., 44: 1947-1959.
CrossRefDirect Link - Franken, A.A., 1970. Sex characteristics and inheritance of sex in asparagus (Asparagus officinalis L.). Euphytica, 19: 277-287.
CrossRefDirect Link