
E.C. Titgemeyer
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
K.S. Spivey
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
L.K. Mamedova
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
B.J. Bradford
Department of Animal Sciences and Industry, Kansas State University, Manhattan, KS, USA
International Journal of Dairy Science
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 134-141







ABSTRACT
The objective of the study was to determine if pharmacological supplies of nicotinic acid could reduce lipolysis in cattle. Six ruminally cannulated steers (225 kg) were used. In an initial study, steers received abomasal infusion of nicotinic acid at 0, 8, or 16 g day-1 then were challenged with a pulse dose of isoproterenol. Nicotinic acid at 16 g day-1 inhibited isoproterenol-stimulated increases in plasma free fatty acid concentrations, whereas 8 g day-1 did not. All 6 steers were then fed 60 mg day-1 zilpaterol-HCl and 3 were continuously abomasally infused with water and 3 with 16 g day-1 nicotinic acid. Steers receiving 16 g day-1 nicotinic acid demonstrated reductions in feed intake and nicotinic acid infusions were terminated after 3.2 days. Plasma glucose and insulin were elevated in response to the nicotinic acid infusion but glucagon was largely unaffected. Elevations in plasma free fatty acids in response to nicotinic acid were observed when feed intake was reduced, suggesting that 16 g day-1 nicotinic acid did not completely block mobilization of fatty acids. Temporal patterns for free fatty acids and insulin did not suggest that elevated free fatty acids were causatively related to insulin resistance during nicotinic acid treatment. Reductions in feed intake of cattle given pharmacological amounts of nicotinic acid indicate there may be risks associated with over-consumption of nicotinic acid.
PDF Abstract XML References Citation
Received: February 25, 2011;
Accepted: May 05, 2011;
Published: June 03, 2011
How to cite this article
E.C. Titgemeyer, K.S. Spivey, L.K. Mamedova and B.J. Bradford, 2011. Effects of Pharmacological Amounts of Nicotinic Acid on Lipolysis and Feed Intake in Cattle. International Journal of Dairy Science, 6: 134-141.
DOI: 10.3923/ijds.2011.134.141
URL: https://scialert.net/abstract/?doi=ijds.2011.134.141
DOI: 10.3923/ijds.2011.134.141
URL: https://scialert.net/abstract/?doi=ijds.2011.134.141
INTRODUCTION
At pharmacological doses, Nicotinic Acid (NA) has been shown to inhibit lipolysis in cattle (Pires and Grummer, 2007; Pires et al., 2007), presumably by acting on the niacin receptor GPR109A, a Gi-coupled receptor (Gille et al., 2008). Activation of GPR109A by NA leads to inhibition of adenylyl cyclase activity and, subsequently, a decrease in cAMP concentrations within the cell. Decreases in cAMP in adipocytes lead to sequelae (inactivation of protein kinase A and decreased phosphorylation of hormone sensitive lipase) that reduce lipolysis (Gille et al., 2008). The NA receptor GPR109A is predominantly localized in adipose tissue and immune cells in humans (Wise et al., 2003) and mice (Tunaru et al., 2003) but present study has indicated a much wider tissue distribution pattern for GPR109A in cattle with the concentrations of GRP109A protein being as great in liver as in adipose tissue (Bradford et al., 2009). In addition, we observed GPR109A mRNA in multiple regions of the bovine brain (our unpublished observations) which was not overly surprising because it has also been found in human brain (Miller and Dulay, 2008).
The United State Food and Drug Administration has approved several β-agonists, including ractopamine and zilpaterol, as growth promotants for cattle. These β-agonists act through β1- and β2-receptors, which are Gs-coupled receptors that increase intracellular concentrations of cAMP (Moody et al., 2000). The β1- and β2-receptors are found in various tissues including muscle and adipose (Sillence and Matthews, 1994). Responses to β-agonists are increases in muscle deposition with reductions in adipose accumulation (Moody et al., 2000).
A large proportion of variation in back fat and intramuscular fat in cattle is dependent on management and environment (Mirzaei et al., 2009), so an improved ability to control fat deposition might help to improve carcass quality. Within adipocytes which contain GPR109A and β-adrenergic receptors, NA and β-agonists have opposing actions. The objectives of the current research were to determine 1) if NA could reduce lipolysis induced by β-agonists, 2) the amount of NA supplementation required to induce this effect and 3) the impact of NA supplementation in cattle fed zilpaterol, a β-adrenergic agonist.
MATERIALS AND METHODS
Animals and management: Experimental procedures were approved by the Kansas State University Institutional Animal Care and Use Committee in 2008. The same 6 ruminally cannulated Holstein steers (initial BW 225±22 kg) were used in these experiments. Steers were fed a grain-based diet (Table 1) near ad libitum intake at 12-h intervals and were housed in metabolism crates throughout the experiments. Infusion lines were placed through the ruminal cannulae and the omasum and into the abomasum, where they were anchored with a 10 cm diameter rubber flange. The infusion lines were designed to allow for continuous post-ruminal infusion of NA. The NA was infused abomasally to prevent ruminal microbial degradation of NA which can be substantial (Santschi et al., 2005).
Dose-response trial: Six steers were used to determine what amount of NA supplementation might reduce lipolysis stimulated by β-agonists. Nicotinic acid (≥99.5%, Fluka Biochemika, Buchs, Switzerland) was continuously infused into the abomasum at 0, 8, or 16 g day-1 in a replicated 3x3 Latin square with 1-day periods.
| Table 1: | Ingredient and nutrient composition of the diet (g kg-1 dry matter) |
 | |
| *Provided (per kg diet DM): Mn: 55 mg, Zn: 55 mg, Cu: 9 mg, I: 0.5 mg, Se: 0.25 mg, Retinyl acetate: 0.76 mg, Cholecalciferol: 6.75 μg, All-rac-α-tocopherol: 27 mg | |
Pires and Grummer (2007) showed almost complete inhibition of lipolysis induced by feed-restriction in non-lactating cows given 6 mg NA/kg BW hourly but lower doses were not tested. Present greatest dose provided NA at about one-half of their rate.
After 1 day of adaptation to the NA treatment, steers were challenged at 3 h after feeding with a pulse dose of isoproterenol-HCl (0.5 μg kg-1 BW; 0.1125 mg in 5 mL saline) into a jugular vein and contralateral jugular blood samples were collected just before isoproterenol dosing as well as 8 min after the isoproterenol challenge. Isoproterenol is a β-agonist that quickly stimulates lipolysis from bovine adipocytes, leading to a peak in plasma Free Fatty Acid (FFA) concentrations 5 to 10 min after dosing (Chilliard and Ottou, 1995). Blood samples (10 mL) were collected in Vacutainer tubes (Becton Dickinson, Franklin Lakes, NJ, USA) containing EDTA as an anticoagulant and immediately placed in ice. Plasma was isolated by centrifugation at 1,000xg for 15 min and immediately analyzed for FFA (Shimizu et al., 1980; NEFA HR kit, Wako Chemicals USA, Richmond, VA, USA).
Zilpaterol feeding and responsiveness to NA: This trial was initiated 3 days after completion of the dose-response trial. Cattle were maintained without any β-agonist in the diet and without NA supplementation for the initial 4 days of this trial. Next, zilpaterol feeding (60 mg day-1) was initiated for all steers and continued without NA treatment for 4 days. Then, NA treatments were initiated; 3 steers received continuous abomasal infusions of 16 g NA in 2 L of water daily and 3 steers received continuous abomasal infusions of water only. All steers continued to receive zilpaterol during the NA treatment period. The initial plan was to maintain steers on their NA treatment for 7 days. However, cattle receiving 16 g day-1 of NA demonstrated progressive reductions in voluntary feed intake, so NA infusions were terminated after 3.2 days of treatment. Steers were maintained on their diets through the initially planned trial (4 days after NA withdrawal) and then euthanized with 187 mg of sodium pentobarbital kg-1 BW followed by exsanguination.
Blood samples were collected 4 h after feeding on day 1 (no treatments applied), day 5 (after 1 day of zilpaterol feeding to all steers), day 8 (4 h after initiation of NA treatments), day 9 (28 h after initiation of NA treatments), day 10 (52 h after initiation of NA treatments), day 12 (1 day after termination of NA treatments) and day 14 (3 days after termination of NA treatments). Jugular blood (10 mL) was collected in Vacutainer tubes containing EDTA, immediately placed in ice and centrifuged at 1,000xg for 15 min to separate plasma. Plasma was frozen and later analyzed for FFA (Shimizu et al., 1980; NEFA HR kit, Wako Chemicals USA), glucose (Raabo and Terkildsen, 1960; Autokit Glucose, Wako Chemicals USA), insulin (Park et al., 2010) and glucagon (RIA kit GL-32K, Linco Research Inc., St. Charles, MO, USA).
Statistical analyses: Data from the dose-titration trial were analyzed as a Latin square with steer included as a random effect and data from the NA responsiveness trial were analyzed as a completely randomized design using the mixed procedure of SAS (SAS Inst. Inc., Cary, NC, USA). Data with repeated observations over time (e.g., feed intake, blood metabolites) were analyzed as repeated measures using the mixed procedure of SAS, with steer as the subject for repeated measures. Means over time were compared using pairwise t-tests.
RESULTS AND DISCUSSION
Isoproterenol challenge: Feed refusals during this trial were small. After the isoproterenol challenge, plasma FFA concentrations were 696 μmol L-1 for control steers, 651 μmol L-1 for steers receiving 8 g day-1 NA and 363 μmol/l for steers receiving 16 g day-1 NA (p<0.01; SEM 166).
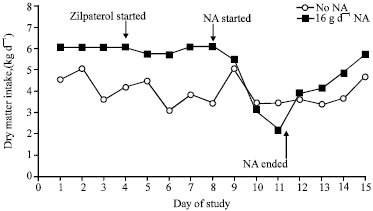 | |
| Fig. 1: | Dry matter intake (SEM = 1.1) of cattle receiving 0 or 16 g day-1 of Nicotinic Acid (NA) abomasally. Cattle received no treatment for the initial 4 days and then received dietary zilpaterol (60 mg day-1) continuously from that point forward. The 16 g day-1 NA treatment was provided continuously from the beginning of day 9 for a total of 3.2 days and then discontinued. Intake never differed from day-1 amounts for steers receiving no NA but for steers receiving 16 g day-1 NA, dry matter intakes were less (p = 0.02) than day-1 amounts on day 11 |
Thus, 16 g day-1 NA was effective in limiting isoproterenol stimulation of FFA release but 8 g day-1 NA was not effective in inhibiting lipolysis. Pre-challenge FFA concentrations were not different among treatments (p = 0.76, Mean = 87 μmol L-1), so they did not affect conclusions.
Feed intake responses to NA: Before NA administration, control cattle consumed less feed than cattle that were to be infused with NA (Fig. 1). The 16 g day-1 dose of NA decreased intake (Fig. 1) and this effect led to feed intakes on the third day of NA infusion (2.2 kg dry matter) being less (p=0.02) than those on day 1 (6.0 kg dry matter). Because feed intakes were decreasing at a rapid rate, NA infusions were terminated after 3.2 days of treatment. Feed intake progressively increased over the next 4 days after termination of NA treatment (Fig. 1).
Depression of feed intake by supplementation with pharmacological amounts of NA has not been previously demonstrated in cattle, likely due to the methodologies that have been employed. In some previous work, pharmacological doses of NA were administered to cows that were feed-restricted for 48 h as a means of inducing lipolysis (Pires and Grummer, 2007; Pires et al., 2007) and thus feed intake responses could not be measured. Piresa et al. (2009) did not observe changes in feed intake when they infused pharmacological doses of NA for 3 days to cows that were fed only 33% of ad libitum intake; however, their research model was unlikely to demonstrate depressions in feed intake due to the experimentally restricted intake as well as the moderately short length of NA infusion. Modest doses of NA (6 g day-1 for 632 kg cows) have been provided post-ruminally to cows for a longer period of time (3 wk) without depressing feed intake (Ottou et al., 1995) but the ineffectiveness of NA in reducing isoproterenol-stimulated lipolysis in that study (Chilliard and Ottou, 1995) suggested that the NA supply was not a pharmacological dose. Other studies have fed relatively large doses of NA to cattle without large effects on feed intake. However, because only a small portion of dietary niacin reaches the small intestine (Santschi et al., 2005), feeding unprotected NA is an ineffective in providing pharmacological amounts of NA to cattle.
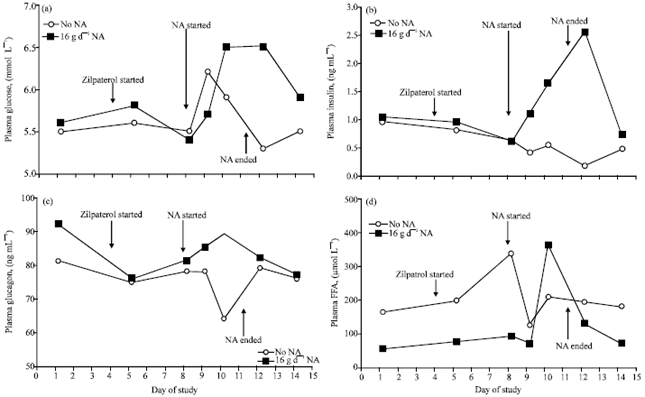 | |
| Fig. 2: | Effects of abomasal supplementation of growing steers with 0 or 16 g day-1 of Nicotinic Acid (NA) on plasma concentrations of (a) glucose; SEM = 0.34; for 16 g day-1 NA, glucose increased (p<0.10) above day-1 concentrations on day 10 and 12 (b) insulin; SEM = 0.49; for 16 g day-1 NA, insulin increased (p=0.04) above day-1 concentrations on day 12 (c) glucagon; SEM = 15 and (d) FFA; SEM = 114; for 16 g day-1 NA, FFA increased (p = 0.07) above day-1 concentrations on day 10. Cattle received no treatment for the initial 4 days and then received dietary zilpaterol (60 mg day-1) continuously from that point forward. The 16 g day-1 NA treatment was provided continuously from the beginning of day 9 for a total of 3.2 days and then discontinued |
We are unaware of any reports from the many human and rodent studies that pharmacological amounts of NA depress intake.
A significant proportion of dairy cattle develop ketosis during the initial 2 months of lactation (Haghighat-Jahromi and Nahid, 2011) and these cows exhibit elevated serum concentrations of β-hydroxybutyrate (Nazifi et al., 2008). Because β-hydroxybutyrate is the endogenous ligand for GPR109A (Gille et al., 2008), it is possible that reductions in feed intake associated with ketosis could be related to stimulation of GPR109A in cattle. This is a particularly intriguing hypothesis because GPR109A is much more widely distributed in cattle than in other species (Bradford et al., 2009).
Blood metabolite responses to NA: Plasma glucose concentration (Fig. 2a) was somewhat elevated (p<0.10) in response to the NA infusion on days 10 and 12 (6.5 mmol L-1 on day 10 and 12 vs. 5.6 mmol L-1 on day 1). This is notable because it occurred in the face of reductions in feed intake. The elevation in blood glucose persisted for at least 1 day after termination of NA treatment but was not different from day-1 values 3 days after termination of NA treatment (5.9 mmol L-1). Ghorbani et al. (2008) demonstrated that 6 or 12 g or niacin fed orally as a solute in water to lactating dairy cows led to increases in plasma glucose.
The NA treatment also elevated plasma insulin concentration (Fig. 2b). There seemed to be a progressive increase in insulin during the initial 2 days that steers received 16 g day-1 NA. Plasma insulin on day 12 (1 day after termination of NA treatment; 2.55 ng mL-1) was greater (p<0.05) than on day 1 (1.0 ng mL-1). The increase in insulin in steers receiving 16 g day-1 NA appeared in the face of reduced feed intake. It is difficult to know if increases in plasma glucose led to the elevation in insulin or if insulin resistance led to the elevations in glucose.
Previous studies with cattle have shown various responses in plasma insulin in response to NA. Feed-restricted cows that received pharmacologic doses of NA infused for 8 h had increased plasma insulin concentrations during the rebound phase, around 4 to 8 h after NA provision was terminated (Pires and Grummer, 2007). Similar short-term treatment of feed-restricted cows with NA transiently (1 to 4 h) reduced plasma insulin and improved glucose clearance at 8 h after NA initiation (Pires et al., 2007). This response was attributed to reductions in plasma FFA concentrations. However, NA treatment of feed-restricted lactating cows for 48 or 72 h but not for 24 h, increased plasma insulin and reduced glucose clearance at 72 h, demonstrating insulin resistance in response to longer-term treatment of cattle with pharmacological doses of NA (Piresa et al., 2009). In goats, Thornton and Schultz (1980) observed increases in blood glucose 2 days after providing an oral pulse-dose of NA (0.8 g kg-0.75 BW); goats also demonstrated impaired glucose tolerance during this time period. In a single cow dosed orally with 160 g of NA, insulin increased more in response to a glucose challenge at 1 or 2 days after the NA dose than before the NA dose (Thornton and Schultz, 1980). Because Thornton and Schultz (1980) used large oral doses of NA, it is difficult to know when and how much of the NA the animals were absorbing. Thus, it is unknown what time periods would lead to direct effects or rebound effects after dissipation of NA supply.
Plasma glucagon was largely unaffected by treatment (Fig. 2c), suggesting that it played a minor role, if any, in the NA effects on blood glucose. However, it is possible that the relatively unchanged plasma glucagon concentrations reflect parallel changes in glucagon secretion and uptake.
Baseline concentrations of FFA (Fig. 2d) were low, at least for steers that later received 16 g day-1 NA; the greater FFA concentrations for the control steers can be attributed to a single steer that demonstrate delevated plasma FFA (over this study, FFA averaged 468, 54 and 75 μmol L-1 for each of the 3 control steers). The elevations in plasma FFA in response to NA on day 10 (360 μmol L-1 vs. 53 μmol L-1 on day 1) could be related to reductions in feed intake; however, the elevation in FFA, which presumably reflects increases in lipolysis, suggests that 16 g day-1 NA did not completely block the steers’ ability to mobilize fatty acids.
Presumably during an NA rebound response, Thornton and Schultz (1980) observed in goats that elevations in plasma insulin occurred before increases in glucose were present which suggests the NA rebound was leading to insulin resistance. Pires and Grummer (2007) suggested that elevations in insulin and glucose during an NA rebound could be induced by the extremely high concentrations of plasma FFA. The temporal patterns that we observed for FFA and insulin do not suggest that elevated FFA concentrations were causatively related to insulin resistance during continuous NA provision.
CONCLUSION
The reductions in feed intake of cattle provided with continuous NA supplementation indicates there may be risks associated with over-consumption of NA. The unusual distribution of GPR109A in comparison to other species is likely related to the impact of NA on feed intake and insulin. It is also possible that NA acts directly on the brain because GPR109A mRNA was also observed in bovine brain, including the hypothalamus (our unpublished observations).
ACKNOWLEDGMENTS
Financial support for this project was provided by Lonza. This study is contribution No. 11-208-J from the Kansas Agricultural Experiment Station, Manhattan.
REFERENCES
- Ghorbani, B., N. Vahdani and S. Zerehdaran, 2008. Effects of niacin on milk production and blood parameters in early lactation of dairy cows. Pak. J. Biol. Sci., 11: 1582-1587.
CrossRefPubMedDirect Link - Gille, A., E.T. Bodor, K. Ahmed and S. Offermanns, 2008. Nicotinic acid: Pharmacological effects and mechanisms of action. Ann. Rev. Pharmacol. Toxicol., 48: 79-106.
PubMed - Haghighat-Jahromi, M. and S. Nahid, 2011. Incidence of subclinical ketosis in dairy cows in fars province of Iran and reproductive performance in early lactetion period. Asian J. Anim. Sci., 5: 158-161.
CrossRefDirect Link - Miller, C.L. and J.R. Dulay, 2008. The high-affinity niacin receptor HM74A is decreased in the anterior cingulate cortex of individuals with schizophrenia. Brain Res. Bull., 77: 33-41.
PubMed - Mirzaei, H.R., A.P. Verbyla, M.P.B. Deland and W.S. Pitchford, 2009. Describing variation in carcass quality traits of crossbred cattle. Pak. J. Biol. Sci., 12: 222-230.
CrossRefPubMedDirect Link - Nazifi, S., M.M. Fani, E. Rowghani and M.R. Behbood, 2008. Studies on the relationship between sub-clinical ketosis and liver injuries within the first two months of lactation period in high producing. Int. J. Dairy. Sci., 3: 29-35.
CrossRefDirect Link - Ottou, J.F., M. Doreau and Y. Chilliard, 1995. Duodenal infusion of rapeseed oil in midlactation cows. 6. Interaction with niacin on dairy performance and nutritional balance. J. Dairy Sci., 78: 1345-1352.
CrossRefDirect Link - Park, A.F., J.E. Shirley, E.C. Titgemeyer, R.C. Cochran, J.M. DeFrain, E.E. Wickersham and D.E. Johnson, 2010. Characterization of plasma metabolites in holstein dairy cows during the periparturient period. Int. J. Dairy Sci., 5: 253-263.
CrossRefDirect Link - Pires, J.A.A. and R.R. Grummer, 2007. The use of nicotinic acid to induce sustained low plasma nonesterified fatty acids in feed-restricted holstein cows. J. Dairy Sci., 90: 3725-3732.
CrossRefPubMedDirect Link - Pires, J.A.A., J.B. Pescara and R.R. Grummer, 2007. Reduction of plasma NEFA concentration by nicotinic acid enhances the response to insulin in feed-restricted holstein cows. J. Dairy Sci., 90: 4635-4642.
CrossRef - Piresa, J.A.A., L.F. Stumpf, I.D. Soutullo, J.B. Pescara, S.E. Stebulis and R.R. Grummer, 2009. Effects of Abomasal Infusion of Nicotinic Acid on Responses to Glucose and &beta,-Agonist Challenges in Partially Feed-Restricted Lactating Cows. In: Ruminant Physiology. Digestion, Metabolism and Effects of Nutrition on Reproduction and Welfare, Chilliard, Y., F. Glasser, Y. Faulconnier, F. Bocquier, I. Veissier and M. Doreau (Eds.)., Wageningen Academic Publishers, Wageningen, The Netherlands, pp: 638-639.
- Raabo, E. and T.C. Terkildsen, 1960. On the enzymatic determination of blood glucose. Scand. J. Clin. Lab. Invest., 12: 402-407.
CrossRefDirect Link - Santschi, D.E., R. Barthiaume, J.J. Matte, A.F. Mustafa and C.L. Girard, 2005. Fate of supplementary B-vitamins in the gastrointestinal tract of dairy cows. J. Dairy Sci., 88: 2043-2054.
CrossRefPubMedDirect Link - Shimizu, S., Y. Tani, H. Yamada, M. Tabata and T. Murachi, 1980. Enzymatic determination of serum-free fatty acids: A colorimetric method. Anal. Biochem., 107: 193-198.
CrossRef - Sillence, M.N. and M.L. Matthews, 1994. Classical and atypical binding sites for beta-adrenoceptor ligands and activation of adenylyl cyclase in bovine skeletal muscle and adipose tissue membranes. Br. J. Pharmacol., 111: 866-872.
PubMed - Thornton, J.H. and L.H. Schultz, 1980. Effects of administration of nicotinic acid on glucose, insulin and glucose tolerance in ruminants. J. Dairy Sci., 63: 262-268.
CrossRefDirect Link - Tunaru, S., J. Kero, A. Schaub, C. Wufka, A. Blaukat, K. Pfeffer and S. Offermanns, 2003. PUMA-G and HM74 are receptors for nicotinic acid and mediate its anti-lipolytic effect. Nat. Med., 9: 352-355.
PubMed - Wise, A., S.M. Foord, N.J. Fraser, A.A. Barnes and N. Elshourbagy et al., 2003. Molecular identification of high and low affinity receptors for nicotinic acid. J. Biol. Chem., 278: 9869-9874.
PubMed