
R. Ikawaty
Department of Medical Microbiology, University Medical Centre Utrecht, Room G04.614, P.O. Box 85500, 3508 GA Utrecht, The Netherlands
E.C. Brouwer
Department of Medical Microbiology, University Medical Centre Utrecht, Room G04.614, P.O. Box 85500, 3508 GA Utrecht, The Netherlands
E. Van Duijkeren
Faculty of Veterinary Medicine Utrecht University, Yalelaan 1, Utrecht, The Netherlands
D. Mevius
Faculty of Veterinary Medicine Utrecht University, Yalelaan 1, Utrecht, The Netherlands
J. Verhoef
Department of Medical Microbiology, University Medical Centre Utrecht, Room G04.614, P.O. Box 85500, 3508 GA Utrecht, The Netherlands
A.C. Fluit
Department of Medical Microbiology, University Medical Centre Utrecht, Room G04.614, P.O. Box 85500, 3508 GA Utrecht, The Netherlands
International Journal of Dairy Science
Year: 2010 | Volume: 5 | Issue: 2 | Page No.: 60-70







ABSTRACT
The pathogenesis of bovine mastitis caused by Staphylococcus aureus and the presence of S. aureus virulence genes in bovine mastitis are poorly understood. In this study, the virulence factor profiles from different genotyped clinical bovine mastitis S. aureus isolates from The Netherlands were investigated. A total of 76 S. aureus isolates from 2 institutions that represented regional and national strains were examined by PCR for 19 toxin genes, 12 adhesin genes, 3 immune evasion genes and 6 other genes. The presence of hlb, ebps, efb, sspA and sspB was observed in all isolates, coa in all isolates but one, while the presence of sec3, seg, seh, sei, sel, sem, sen, tst, lukE, fnbA, fnbB, icaB, icaC, icaD, clfA, sdrE, cna, cap8, cap5 and map was variable. Sak and the genes for the human specific immune evasion proteins, SCIN and CHIPS were detected in only one isolate. In addition, none of isolates were lukPV-positive. Five major virulence factor profiles were observed which, with the exception of SaPIbov, were in general agreement with the clustering obtained from PFGE, MLST, spa-typing and MLVA. In conclusion, hlb, ebps, efb, sspA and sspB were present in all isolates and other virulence genes were variably present. The virulence factor profiles generally matched genotyping results for Dutch bovine mastitis isolates.
PDF Abstract XML References Citation
How to cite this article
R. Ikawaty, E.C. Brouwer, E. Van Duijkeren, D. Mevius, J. Verhoef and A.C. Fluit, 2010. Virulence Factors of Genotyped Bovine Mastitis Staphylococcus aureus Isolates in The Netherlands. International Journal of Dairy Science, 5: 60-70.
DOI: 10.3923/ijds.2010.60.70
URL: https://scialert.net/abstract/?doi=ijds.2010.60.70
DOI: 10.3923/ijds.2010.60.70
URL: https://scialert.net/abstract/?doi=ijds.2010.60.70
INTRODUCTION
Virulence of a microorganism can be defined as the capacity of the microorganism to damage the host which may lead to signs and symptoms of disease. A large array of virulence factors are produced by Staphylococcus aureus. S. aureus may produce a number of exotoxins including the classical Staphylococcal Enterotoxins (SEs) A to E, the recently characterized SE types (SEG-SEU) (Dinges et al., 2000; Smyth et al., 2005), toxic shock syndrome toxin 1 (TSST-1), β-hemolysin (hlb), the bi-component Panton-Valentine Leukocidin (Luk-PVL), proteases with the ability to induce T-cell proliferation and exfoliative toxin A and B (ETA and ETB). A number of Microbial Surface Components Recognizing Adhesive Matrix Molecules (MSCRAMMs) which promote adhesion to host extra-cellular matrix components (Foster, 2005). Clumping factor A and B (ClfA and ClfB) are two dominant fibrinogen-binding adhesins (Foster, 2005), while fibronectin-binding protein A and B (FnBPA and FnBPB) mediate binding to fibronectin and collagen adhesin (CNA) binding to collagen. Serine-aspartate repeat proteins (Sdr) are surface proteins related to ClfA and ClfB (Josefsson et al., 1998) which have been hypothesized to mediate interactions of S. aureus with the extra-cellular matrix (George et al., 2006). Coagulase induces polymerization of fibrinogen to fibrin. Distinct from coagulase, fibrinogen-binding protein (Efb) is an intracellular protein which specifically promotes binding to fibrinogen (Bodén and Flock, 1989). Elastin-binding protein of S. aureus (EbpS) mediates binding to soluble tropoelastin (Downer et al., 2002). Adherence of S. aureus is also enhanced by a MHC class II analog protein (MAP), a protein with broad binding activity to fibrinogen, collagen, fibronectin, vitronectin and elasqtin which may enable colonization of different tissues. Plasmin-sensitive protein (Pls) has an adhesive function by mediating interaction between bacterial cells, bacterial binding to immobilized fibronectin and immunoglobulin G and invasion of epithelial cells, as well as anti-adhesive functions at a certain stages during infection which allow the bacteria to spread. S. aureus can produce biofilm which is mediated by gene products of the icaADBC locus (Rohde et al., 2007). S. aureus exoenzymes including the extra-cellular serine protease (SspA) and cysteine protease (SspB) are involved in elastin degradation known as elastinolytic activity.
Capsular Polysaccharide (CP) produced by S. aureus renders the bacteria resistant to phagocytosis. Eleven CP serotypes have been described and CP type 5 and 8 were predominantly found in clinical isolates (O’Riordan and Lee, 2004; Poutrel et al., 1988; Sordelli et al., 2000). Furthermore, S. aureus phages encoding immune evasion molecules such as staphylokinase (SAK), chemotaxis inhibitory protein of Staphylococcus aureus (CHIPS) and staphylococcal complement inhibitor (SCIN) may provide S. aureus the ability to escape the human immune system. In addition, an Arginine Catabolic Mobile Element (ACME) which inhibits polymorphonuclear cell production, is considered to enhance the virulence and colonization by S. aureus (Diep et al., 2006a, b).
S. aureus is generally recognized as the most common pathogen of contagious bovine mastitis and it causes significant economic losses in the diary industry. Chronic and deep infections in the mammary glands of bovines are often associated with poor success of treatment. Different suites of virulence factors may result in differences in severity and course of mastitis. However, little is known about the presence of S. aureus virulence genes in bovine mastitis and so far, dominant virulence factors among bovine mastitis S. aureus have not been identified. In this study, we investigated the virulence factor profiles from different genotyped clinical bovine mastitis S. aureus isolates from The Netherlands.
MATERIALS AND METHODS
Bacterial Isolates
The study was conducted between September 2007 and September 2009 at the Department of Medical Microbiology of the University Medical Center Utrecht, Utrecht, The Netherlands. Seventy six Staphylococcus aureus isolates from clinical cases of bovine mastitis were included. Thirty isolates were obtained by the Faculty of Veterinary Medicine Utrecht University, Utrecht, The Netherlands, from 26 farms near Utrecht. Isolates from farms sampled twice were taken at least one year apart. Another 46 isolates were collected by the Animal Health Service in Deventer, The Netherlands for the Central Veterinary Institute (CVI), Lelystad and The Netherlands from farms throughout The Netherlands.
 | |
| Fig. 1: | Virulence factor profiles observed within various typing methods. Place where the isolates were taken. FVM: Veterinary Faculty Utrecht University; CVI: Central Veterinary Institute, Lelystad, The Netherlands. The presence and absence of genes is indicated by red and green spots. Virulence factor profiles were defined based on the presence and absence of the genes, and resulted 5 main profiles, A, B, C, D and E |
The sources were individual teat milk samples from dairy cattle from all over the country from clinical or subclinical cases of mastitis. Each isolate from CVI represents one farm, the location of the farms is unknown but they are distributed all over The Netherlands. All isolates were methicillin-susceptible S. aureus (MSSA) and collected between 1988 and 2005.
These isolates were genotyped by Multilocus Sequence Typing (MLST), staphylococcus protein A (spa)-typing, Multiple Locus Variable Number Tandem of Repeat Analysis (MLVA) and Pulsed-Field Gel Electrophoresis (PFGE) (Fig. 1) (Ikawaty et al., 2008). Most of the 76 bovine S. aureus isolates used in this study belonged to 4 dominant MLST types. We observed 3 dominant spa-types and 2 dominant MLVA types. DNA of 76 isolates was digested with SmaI and showed 21 PFGE profiles, 4 dominant profiles, 10 profiles related to the dominant profiles and 7 unique PFGE profiles (Fig. 1). No particular difference was observed between regionally and nationally obtained S. aureus bovine mastitis strains in terms of PFGE profile, MLST and spa-types, except for MT118 that was only present among regionally obtained isolates. Despite the fact that isolates could belong to the same cluster, they were not considered to belong to local outbreaks as at least 90% of the isolates were obtained on different farms whereas the remaining isolates were obtained at least one year apart when sampled on the same farm. MLST generally showed a distinct cluster of bovine S. aureus compared to isolates from the human population, although a little sharing of strains between the bovine and human population was observed. In addition, the ST or Clonal Complexes (CC) of bovine mastitis isolates from The Netherlands differed from those described elsewhere with the exception of 4 isolates (S0337, S0426, S0431 and S0436) that belonged to CC97 which has previously been reported to be obtained from humans.
Genomic DNA Preparation
Preparation of bacterial genomic DNA was performed using the NucleoSpin kit (Macherey-Nagel) following the protocols from the manufacturer with the exception that the bacterial pellet is resuspended in 20 mM Tris/HCl, 2 mM EDTA and 1% Triton X-100 supplemented with 0.2 mg mL-1 of lysostaphin, achromopeptidase and RNase.
PCR Amplifications
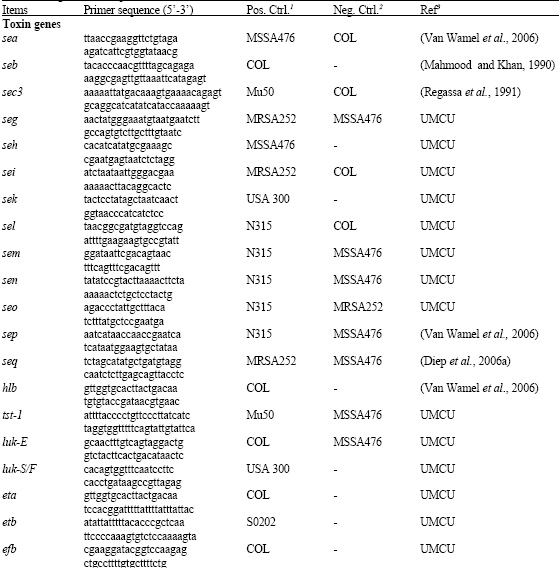
PCRs were performed to detect the genes of 40 potential virulence factors including 19 staphylococcal toxin genes, 12 adhesion genes; 3 genes encoding extra-cellular proteases, 3 immune evasion genes, 2 genes for capsular polysaccharide and SCCACME (Table 1). Table 1 lists the oligonucleotide primers, positive and negative control strains used in this study.
| Table 1: | Oligonucleotide primers used |
 | |
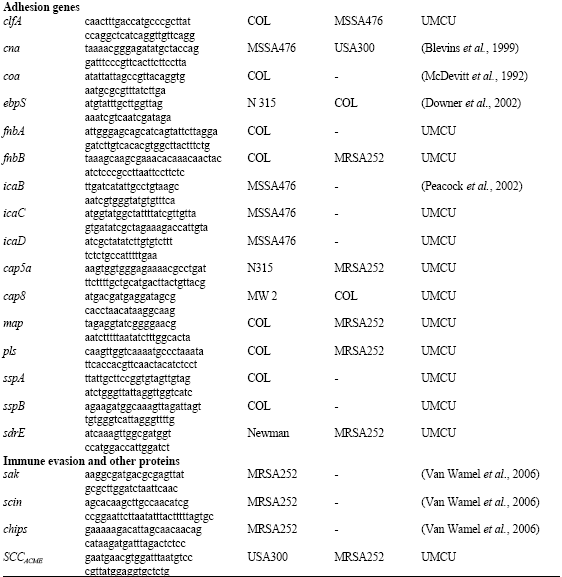 | |
1Positive control strains; 2Negative control strains; 3References. UMCU: Primers were designed at Dept. of Medical Microbiology, University Medical Centre Utrecht, Utrecht, The Netherlands | |
RESULTS AND DISCUSSION
Among the 18 toxin genes tested, hlb, sspA and sspB were detected in all isolates. The genes for enterotoxins A (sea), B (seb), K (sek), O (seo), P (sep), Q (seq), leukocidin encoded by LukS/F, exfoliative toxin A (eta) and B (etb) could not be amplified while the genes encoding the other 8 toxins were present in 1-80% of the isolates. The recently described enterotoxins G and I were the most prevalent (78 and 80% of the isolates, respectively). We observed that 56 of 76 (76%) isolates encoded both SEI and SEG. Forty (52.6%) isolates encoded SEM and SEN, although these two genes were not present simultaneously in 27 (35.5%) isolates. The seh gene was present in only 1 isolate (1%). The SaPIbov pathogenicity island signature combination of sec3, sel and tst was found in 15 (19.7%) isolates. Two (2.6%) isolates encoded sec3 and tst but lacked sel, while 6 (7.9%) other isolates encoded either sec3, sel or tst only. In addition, lukE was detected in 20% of isolates.
Ebps and efb could be amplified for all isolates, while pls was absent. PCR specific for fnbA and fnbB showed that 73 (96%) isolates possessed fnbA and 33 (43%) isolates had fnbB. Three gene targets involved in biofilm formation, icaB, icaC and icaD, were found in 64 (84%), 63 (83%) and 65 (86%) isolates, respectively. ClfA was detected in 16 (21%) isolates, 33% (n = 16) isolates were positive for sdrE, 49% (n = 37) were cna positive, 18% (n = 14) contained map and most of isolates (99%, n=75) obtained coa.
The three immune evasion genes, sak, scin and chips, could not be amplified in almost all isolates, except for one isolate (S0336). SCCACME was not amplified in any of the isolates. Cap8 was detected in 73 isolates, while cap5 was present in 3 isolates.
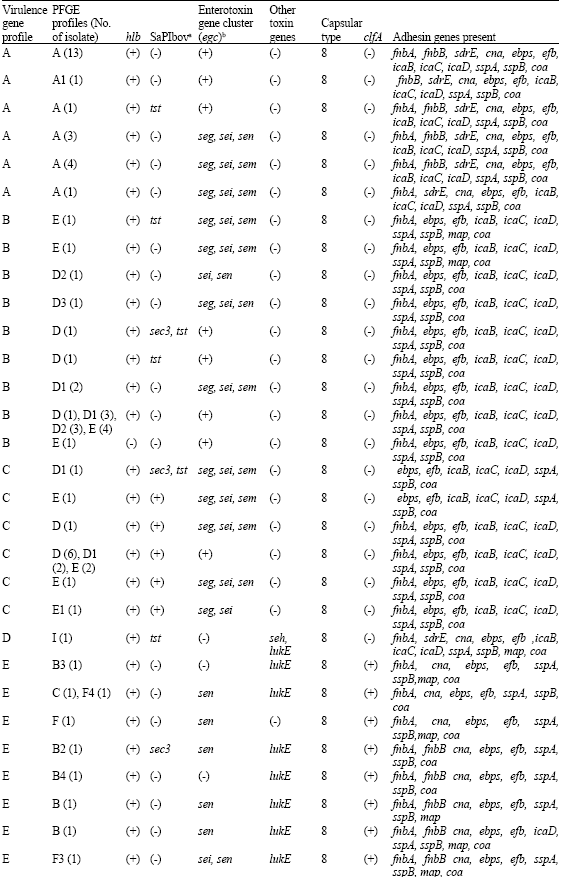
Based on the presence and absence of virulence factors five main clusters (A, B, C, E and F) were identified among the 76 bovine mastitis S. aureus isolates (Fig. 1). In addition, a unique gene pattern (called D) was observed for 1 isolate (S0366). There is general agreement between the 4 typing methods used (PFGE, MLST, spa-typing and MLVA) and the 6 clusters of isolates, but exceptions were obviously present. Cluster B and C could not be identified based on PFGE profiles, since some of the isolates with an identical PFGE profile were distributed across both virulence profile clusters. These isolates were also considered closely related based on the other typing methods (MLST, spa-typing and MLVA).
Cluster A isolates mostly possessed sdrE, finbB and cna genes which were totally absent in cluster B and C isolates and occasionally present in cluster D and E isolates (Table 2). Identical and closely related isolates identified by MLST, spa-typing and MLVA in cluster B and C had similar virulence gene patterns, except that SaPIbov (tst, sec3 and sel) were predominantly detected in cluster C isolates. Virulence gene patterns detected in cluster E and F isolates mostly lack SaPIbov similar to isolates of cluster A. The gene for enterotoxin sem was absent, while sen, seg and sei were occasionally detected. Almost all isolates from both clusters contained lukE, clfA and map, while other clusters did not. icaB-D and cap8 were detected in all clusters (except cluster E isolates). The only isolate in cluster D showed a unique gene pattern with the presence of seh and a unique combination of other virulence genes.
Thus in general, there is an agreement between the virulence profile and the typing methods and the clusters with exception of clusters B and C. Cluster C contains SaPIbov-positive isolates, whereas this is absent in cluster B isolates, but the overall virulence gene profiles are similar for both clusters.
By analyzing the presence of all virulence factors tested in all isolates fnbA, hlb, ebps, sspA, sspB and coa were always present in Dutch isolates, while cap8, sem, seg, sei, icaB, icaC, icaD and sen were variable present.
The presence of virulence factor encoding genes and variation in their presence in extensively genotyped S. aureus isolates collected from bovine mastitis was investigated. Hlb, ebps, efb, sspA and sspB were found in all isolates tested and coa in all isolates except one, while sea, seb, sek, seo, sep, seq, PV- LukS/F, eta, etb, pls and SCCACME were completely absent. Variation in presence was shown for 9 toxin genes: sec3, seg, seh, sei, sel, sem, sen, tst and lukE and 9 adhesion related genes: fnbA, fnbB, icaB, icaC, icaD, clfA, sdrE, cna and map and other genes such as scin, chips, cap8 and cap5. The clustering of tst, sec3 and sel genes is specific for a bovine pathogenicity island known as SaPIbov (Smyth et al., 2005) was present in only part of the isolates. From these data a minimum set of genes which may be required for S. aureus bovine mastitis was distilled which includes hlb, efb, ebps, sspA, sspB and fnbA. However, efb has been reported to be present in only part of the isolates by others (Zecconi et al., 2006). Probably other virulence factors do contribute to the development and severity of disease. Details of the pathogenesis of bovine mastitis may differ for different gene profiles.
| Table 2: | Presence and absence of toxin and adhesion gene profiles in different bovine PFGE-types |
 | |
 | |
aBovine staphylococcal pathogenicity island characterized by the presence of sec, sel, and tst. The (+) means sec, sel and tst genes were amplified, while the (-) means none of the genes were detected. Written gene/s depicted their presence. bEnterotoxin gene cluster (egc) consists of 5 SEs, seg, sei, sem, sen, and seo. Since all of our isolates lack of seo, the (+) indicated the presence of seg, sei, sem and sen | |
The virulence factor gene profiles in general agreed with the clustering obtained from the 4 genotyping methods. The exception of cluster B and C was caused by the presence of SaPIbov in a part of the isolates belonging to PFGE types D, D1, D2 and E. This result indicates that SaPIbov was acquired independently by several isolates belonging to different genetic lineages. SaPIbov was present in 19.7% of the isolates and10.5% of all isolates carried either 1 or 2 genes representative for SaPIbov. The tst gene has also been reported on other pathogenicity islands (Lindsay et al., 1998). Whether the presence of sec3 and sel in two isolates and sec3 or sel in other isolates implicates that SaPIbov is incomplete or that the sec3 and sel can be present on other locations is unknown. Although SaPIbov may contribute to mastitis it not required, because other lineages appear to be equally widespread as cluster C (Fig. 1).
The presence of hlb, coa in nearly all isolates is in agreement with other reports (Dinges et al., 2000; Zecconi et al., 2006). The genes that were not identified among Dutch bovine mastitis isolates were also generally absent from isolates from other countries with the exception of sea, sek, seo and eta and etb (Fournier et al., 2008; Haveri et al., 2007, 2008; Monecke et al., 2007; O’Riordan and Lee, 2004; Vautor et al., 2009; Zecconi et al., 2006). Sea and seo for example have been reported to be present in 77 and 52% of mastitis isolates in Switzerland (Fournier et al., 2008). However, Turkish isolates also lacked sea, but eta, etb, sek and seo were not tested (Karahan et al., 2009). The variability of the presence of other toxin genes reported here is in general agreement with other reports (Fournier et al., 2008; Haveri et al., 2007; Kenny et al., 1993; Monecke et al., 2007; Zecconi et al., 2006). The coexistence of the toxin genes seg and sei has been described before and further explained by the presence of both genes in a tandem orientation and belong to an operon of enterotoxin gene cluster (egc) (Jarraud et al., 2001; Karahan et al., 2009). In addition, we detected sem and sen within seg- and sei- harboring S. aureus isolates without the presence of seo. This finding is in contrast to those studies which demonstrated that sem, sen, seg, sei and seo predominate in S. aureus from animal hosts (Haveri et al., 2007; Smyth et al., 2005). The reason for these discrepancies is not known, although the number of different genetic clusters observed among the Dutch isolates was low. In addition, many of the virulence factors are encoded on mobile genetic elements which can easily be lost or exchanged.
None of our isolates were lukPV-positive in agreement with the study done by Fueyo et al. (2005), although, Zecconi et al. (2006) identified pvl in more than 50% of isolates from bovine mastitis (Zecconi et al., 2006). However, also among human methicillin-susceptible S. aureus only a few percent of the isolates carry the pvl genes.
One of the common features of invasive bacterial pathogens is a capsule. In this study, the cap8 was predominant compared to the cap5. Poutrel et al. (1988) described the difference in distribution of CP5 and CP8 between clinical S. aureus isolates collected from humans and bovines. He observed 51.4% of S. aureus from bovine origin were CP5 positive and 18% were CP8 positive, while the percentages of human S. aureus harboring CP5 and CP8 were 20 and 60%, respectively. Several studies described that CP5 and CP8 were commonly found in the clinical human S. aureus isolates (Sordelli et al., 2000). Since CP 5 and 8 could be recovered from either human or animal derived S. aureus isolates, we conclude that CP5 and 8 are non-host specific.
Only one isolate carried the genes for CHIPS and SCIN, which interfere with activation of the human complement system but not the bovine complement system. Both genes are located on the same bacteriophage that is integrated in the hlb gene (Van Wamel et al., 2006). The hlb product is poorly effective against human erythrocytes, but highly effective against bovine erythrocytes and bacteriophage insertion disrupts the hlb gene. Therefore, we consider this isolate as human derived. ST97 isolates have been obtained from both humans and bovines (Smith et al., 2005). The gene profiles of these isolates resemble those of bovine mastitis isolates and lack the human complement specific interference factors (Table 2 and Fig. 1). Based on these data we believe that ST97 S. aureus isolates are from bovine origin.
CONCLUSION
In summary the virulence factor profiles with the exception of SaPIbov were generally in agreement with the typing results. This suggests independent mobilization of this pathogenicity island. Five major virulence factor profiles were observed. Of the tested virulence genes hlb, ebps, sspA, sspB, fnbA and coa were present in all isolates from bovine mastitis.
REFERENCES
- Blevins J.S., A.F. Gillaspy, T.M. Rechtin, B.K. Hurlburt and M.S. Smeltzer, 1999. The Staphylococcal accessory regulator (sar) represses transcription of the Staphylococcus aureus collagen adhesin gene (can) in an agr-independent manner. Mol. Microbiol., 33: 317-326.
PubMed - Boden, M.K. and J.I. Flock, 1989. Fibrinogen-binding protein/clumping factor from Staphylococcus aureus. Infect. Immun., 57: 2358-2363.
PubMed - Diep, B.A., H.A. Carleton, R.F. Chang, G.F. Sensabaugh and F. Perdreau-Remington, 2006. Roles of 34 virulence genes in the evolution of hospital- and community-associated strains of methicillin-resistant Staphylococcus aureus. J. Infect. Dis., 193: 1495-1503.
PubMed - Diep, B.A., S.R. Gill, R.F. Chang, T.H. Phan and J.H. Chan et al., 2006. Complete genome sequence of USA300, an epidemic clone of community-acquired methicillin-resistant Staphylococcus aureus. Lancet, 367: 731-739.
PubMed - Dinges, M.M., P.M. Orwin and P.M. Schlievert, 2000. Exotoxins of Staphylococcus aureus. Clin. Microbiol. Rev., 13: 16-34.
CrossRefPubMedDirect Link - Downer, R., F. Roche, P.W. Park, R.P. Mecham and T.J. Foster, 2002. The elastin-binding protein of Staphylococcus aureus (EbpS) is expressed at the cell surface as an integral membrane protein and not as a cell wall-associated protein. J. Biol. Chem., 277: 243-250.
PubMed - Fournier, C., P. Kuhnert, J. Frey, R. Miserez and M. Kirchhofer et al., 2008. Bovine Staphylococcus aureus: Association of virulence genes, genotypes and clinical outcome. Res. Vet. Sci., 85: 439-448.
PubMed - Fueyo, J.M., M.C. Mendoza, M.R. Rodicio, J. Muniz, M.A. Alvarez and M.C. MartIn, 2005. Cytotoxin and pyrogenic toxin superantigen gene profiles of Staphylococcus aureus associated with subclinical mastitis in dairy cows and relationships with macrorestriction genomic profiles. J. Clin. Microbiol., 43: 1278-1284.
CrossRefDirect Link - George, N.P.E., Q. Wei, P.K. Shin, K, Konstantopoulos and J.M. Ross, 2006. Staphylococcus aureus adhesion via Spa, ClfA and SdrCDE to immobilized platelets demonstrates shear-dependent behavior. Arterioscler. Thromb. Vasc. Biol., 26: 2394-2400.
PubMed - Haveri, M., A. Roslof, L. Rantala and S. Pyorala, 2007. Virulence genes of bovine Staphylococcus aureus from persistent and nonpersistent intramammary infections with different clinical characteristics. J. Applied Microbiol., 103: 993-1000.
PubMed - Haveri, M., M. Hovinen, A. Roslof and S. Pyorala, 2008. Molecular types and genetic profiles of Staphylococcus aureus strains isolated from bovine intramammary infections and extramammary sites. J. Clin. Microbiol., 46: 3728-3735.
PubMed - Ikawaty, R., E.C. Brouwer, M.D. Jansen, E. van Duijkeren, D. Mevius, J. Verhoef and A.C. Fluit, 2008. Characterization of Dutch Staphylococcus aureus from bovine mastitis using a multiple locus variable number tandem repeat analysis. Vet. Microbiol., 136: 277-284.
PubMed - Jarraud, S., M.A. Peyrat, A. Lim, A. Tristan and M. Bes et al., 2001. egc, a highly prevalent operon of enterotoxin gene, forms a putative nursery of superantigens in Staphylococcus aureus. J. Immunol., 166: 669-677.
PubMed - Josefsson, E., K.W. McCrea, D. Ni Eidhin, D. O'Connell, J. Cox, M. Hook and T.J. Foster, 1998. Three new members of the serine-aspartate repeat protein multigene family of Staphylococcus aureus. Microbiology, 144: 3387-3395.
PubMed - Karahan, M., M.N. Acik and B. Cetinkaya, 2009. Investigation of toxin genes by polymerase chain reaction in Staphylococcus aureus strains isolated from bovine mastitis in Turkey. Foodborne Pathog. Dis., 6: 1029-1035.
PubMed - Kenny, K., R.F. Reiser, F.D. Bastida-Corcuera and N.L. Norcross, 1993. Production of enterotoxins and toxic shock syndrome toxin by bovine mammary isolates of Staphylococcus aureus. J. Clin. Microbiol., 31: 706-707.
PubMed - Lindsay, J.A., A. Ruzin, H.F. Ross, N. Kurepina and R.P. Novick, 1998. The gene for toxic shock toxin is carried by a family of mobile pathogenicity islands in Staphylococcus aureus. Mol. Microbiol., 29: 527-543.
PubMed - Mahmood, R. and S.A. Khan, 1990. Role of upstream sequences in the expression of the staphylococcal enterotoxin B gene. J. Biol. Chem., 265: 4652-4656.
PubMed - McDevitt, D., P. Vaudaux and T.J. Foster, 1992. Genetic evidence that bound coagulase of Staphylococcus aureus is not clumping factor. Infect. Immun., 60: 1514-1523.
PubMed - Monecke, S., P. Kuhnert, H. Hotzel, P. Slickers and R. Ehricht, 2007. Microarray based study on virulence-associated genes and resistance determinants of Staphylococcus aureus isolates from cattle. Vet. Microbiol., 125: 128-140.
PubMed - O'Riordan, K. and J.C. Lee, 2004. Staphylococcus aureus capsular polysaccharides. Clin. Microbiol. Rev., 17: 218-234.
PubMed - Peacock, S.J., C.E. Moore, A. Justice, M. Kantzanou and L. Story et al., 2002. Virulent combinations of adhesin and toxin genes in natural populations of Staphylococcus aureus. Infect. Immunity, 70: 4987-4996.
CrossRefPubMedDirect Link - Poutrel, B., A. Boutonnier, L. Sutra and J.M. Fournier, 1988. Prevalence of capsular polysaccharide types 5 and 8 among Staphylococcus aureus isolates from cow, goat and ewe milk. J. Clin. Microbiol., 26: 38-40.
PubMed - Regassa, L.B., J.L. Couch and M.J. Betley, 1991. Steady-state staphylococcal enterotoxin type c mRNA is affected by a product of the accessory gene regulator (agr) and by glucose. Infect. Immun., 59: 955-962.
PubMed - Rohde, H., E.C. Burandt, N. Siemssen, L. Frommelt and C. Burdelski et al., 2007. Polysaccharide intercellular adhesin or protein factors in biofilm accumulation of Staphylococcus epidermidis and Staphylococcus aureus isolated from prosthetic hip and knee joint infections. Biomaterials, 28: 1711-1720.
PubMed - Smith, E.M., L.E. Green, G.F. Medley, H.E. Bird and L.K. Fox et al., 2005. Multilocus sequence typing of intercontinental bovine Staphylococcus aureus isolates. J. Clin. Microbiol., 43: 4737-4743.
PubMed - Smyth, D.S., P.J. Hartigan, W.J. Meaney, J.R. Fitzgerald, C.F. Deobald, G.A, Bohach and C.J. Smyth, 2005. Superantigen genes encoded by the egc cluster and SaPIbov are predominant among Staphylococcus aureus isolates from cows, goats, sheep, rabbits and poultry. J. Med. Microbiol., 54: 401-411.
PubMed - Sordelli, D.O., F.R. Buzzola, M.I. Gomez, L. Steele-Moore and D. Berg et al., 2000. Capsule expression by bovine isolates of Staphylococcus aureus from Argentina: Genetic and epidemiologic analyses. J. Clin. Microbiol., 38: 846-850.
PubMed - Van Wamel, W.J.B., S.H.M. Rooijakkers, M. Ruyken, K.P. van Kessel and J.A. van Strijp, 2006. The innate immune modulators staphylococcal complement inhibitor and chemotaxis inhibitory protein of Staphylococcus aureus are located on β-hemolysin-converting bacteriophages. J. Bacteriol., 188: 1310-1315.
PubMed - Vautor, E., V. Magnone, G. Rios, K. Le Brigand and D. Bergonier et al., 2009. Genetic differences among Staphylococcus aureus isolates from dairy ruminant species: A single-dye DNA microarray approach. Vet. Microbiol., 133: 105-114.
PubMed - Zecconi, A., L. Cesaris, E. Liandris, V. Dapra and R. Piccinini, 2006. Role of several Staphylococcus aureus virulence factors on the inflammatory response in bovine mammary gland. Microb. Pathog., 40: 177-183.
CrossRefPubMedDirect Link