
Jingxiu Xuan
Department of Medicine, SUNY at Buffalo School of Medicine and Biomedical Sciences, Buffalo, NY
Long Shen
Department of Medicine, SUNY at Buffalo School of Medicine and Biomedical Sciences, Buffalo, NY
Chongjie Zhang
Department of Immunology, Sichuan University, Sichuan, China
Richard J. Ford
Department of Hemato-pathology, MD Anderson Cancer Center, Houston, Texas
Julian L. Ambrus
Department of Medicine, SUNY at Buffalo School of Medicine and Biomedical Sciences, Buffalo, NY
International Journal of Cancer Research
Year: 2012 | Volume: 8 | Issue: 3 | Page No.: 83-94







ABSTRACT
Follicular lymphoma is a common B cell-derived malignancy that often follows a prolonged indolent phase followed by a terminal aggressive phase. The growth requirements for follicular lymphomas are poorly understood. B cells from patients with follicular lymphoma die rapidly when grown in culture. We have been able to keep B cells from patients with follicular lymphomas alive in vitro for 20 days using native Interleukin 14 (IL-14) which was originally identified as a B cell growth factor. Native IL-14 contains two molecules produced from the IL14 gene, IL-14α and IL-14β. Interleukin 14α utilizes exons 3-10 while IL-14β is produced from the opposite strand of the IL14 gene to IL-14α and utilizes only exon 10. We have previously demonstrated that transgenic mice expressing IL-14α develop autoimmunity and large B cell lymphomas. In this manuscript we demonstrate that IL-14β transgenic mice develop lymphomas that closely resemble follicular lymphoma. When IL-14β Tg mice are crossed with c-Myc transgenic mice, the Double Transgenic mice (DTgβ) are born with lymphomas resembling the aggressive phase of follicular lymphoma. We also demonstrate that the mRNA for IL-14β is expressed constitutively in normal B cells, T cells and follicular dendritic cells as well as in the Burkitt lymphoma line Namalva. The expression of IL-14β mRNA is decreased with cell activation and in the spleen cells of autoimmune (NZBxNZW) F1 mice. Thus, IL-14β is a distinct protein with differential regulation and physiological effects from IL-14α. IL-14β Tg mice and DTgβ mice are new animal models to study the dormant and aggressive phases of follicular lymphoma.
PDF Abstract XML References Citation
Received: November 12, 2011;
Accepted: March 08, 2012;
Published: May 26, 2012
How to cite this article
Jingxiu Xuan, Long Shen, Chongjie Zhang, Richard J. Ford and Julian L. Ambrus, 2012. IL-14β Transgenic Mice Develop Tumors Consistent with Follicular Lymphoma. International Journal of Cancer Research, 8: 83-94.
DOI: 10.3923/ijcr.2012.83.94
URL: https://scialert.net/abstract/?doi=ijcr.2012.83.94
DOI: 10.3923/ijcr.2012.83.94
URL: https://scialert.net/abstract/?doi=ijcr.2012.83.94
INTRODUCTION
Follicular lymphoma is the second most common form of non-Hodgkin lymphoma and represents 20-30% of all cases (Harris et al., 1994; Tan and Horning, 2008; Van Besien and Schouten, 2007; Vitolo et al., 2008; Abdel-Fattah and Yassine, 2006). Most patients are diagnosed with advanced disease because it only becomes symptomatic after it has spread diffusely, usually after growing indolently for up to 10 years (Tan and Horning, 2008). The tumor grows as what appears to be an expansion of normal germinal center B cell growth that includes follicular dendritic cells and follicular CD4+ T cells and are dependent upon the signals provided by the helper T cells (Kuppers, 2005). One of the characteristic genetic changes of follicular lymphoma is a t (14; 18) translocation which places the bcl-2 gene under the control of the heavy chain promoter (Van Besien and Schouten, 2007). However, normal cells have been found with this translocation and over-expression of bcl-2 in transgenic mice rarely leads to the development of lymphoma (Limpens et al., 1991; McDonnell et al., 1989; McDonnell and Korsmeyer, 1991). Other genes must participate in the pathophysiology of this tumor and in fact most patients have a complex karyotype with multiple chromosomal changes besides t (14; 18) (Hoglund et al., 2004). Interestingly, there are also patients with classical follicular lymphoma who lack the (14; 18) translocation (Biagi and Seymour, 2002). Various strategies have been attempted to identify other contributing genes, including micro-array and spectral karyotyping with array-based high-resolution chromosomal analysis (De Jong, 2005; Husson et al., 2002). This is an important area of ongoing research (Aurora and Winter, 2006; Roulland et al., 2008). The study on follicular lymphoma is hampered by the lack of any good animal models and the inability to grow the tumors in vitro.
Interleukin 14 (IL-14) was initially identified as a B cell growth factor activity in the supernatant of a Burkitt lymphoma cell line (Ambrus and Fauci, 1985). It was cloned using a monoclonal antibody and polyclonal antiserum that turned out to recognize a protein encoded by the positive strand of the IL14 gene, utilizing exons 3-10 which we have called IL-14α(2). IL-14α transgenic mice were produced and demonstrated to have many of the activities predicted for IL-14: expanded B cell numbers, especially B1 cells, enhanced antibody responses to T-dependent and T-independent antigens, autoimmunity most consistent with primary Sjogren’s disease and development of large B cell lymphoma in old age (Shen et al., 2006, 2009, 2010). When IL-14α transgenic mice were crossed with C-Myc transgenic mice, double transgenic mice rapidly developed a tumor closely resembling blastoid-variant mantle cell lymphoma (Ford et al., 2007).
At the same time, IL-14 protein produced from Burkitt lymphoma cells was purified and subjected to Edman degradation amino acid sequencing. Some of the peptides obtained by this methodology appeared to encoded by the negative rather than the positive strand of the IL14 gene utilizing only exon 10 (Ambrus et al., 1996). We have called the protein encoded by this transcript IL-14β. The purpose of the studies in this manuscript was to evaluate the expression of the IL-14β transcript and the physiological effects of IL-14β utilizing transgenic mice. We show that IL-14β Tg mice develop lymphoma that closely resembles follicular lymphoma: (1) it has the histological appearance of follicular lymphoma, (2) grows in the presence of T lymphocytes and macrophages, (3) expresses CD19, CD22 and CD79a but not CD5, CD21 or CD23 and (4) express bcl-2 at high levels. When IL-14β transgenic mice were bred with C-Myc transgenic mice, the IL-14β c-Myc double transgenic mice (DTgβ) rapidly developed tumors that closely resemble the aggressive phase of follicular lymphoma.
MATERIALS AND METHODS
Growth of follicular lymphoma cells in vitro: Freshly isolated follicular lymphoma cells were cultures in RPMI 1640 and 20% Fetal Calf Serum (FCS) (Hyclone, Ogden, UT) with IL-14 containing Namalva supernatants Ambrus and Fauci (1985) at 37°C in an incubator containing 5% CO2.
Production of il-14β transgenic mice: IL-14β transgenic mice were produced using the pE μSR vector expressing human IL-14β in the same manner as the production of IL-14α transgenic mice (Shen et al., 2006).
Histological evaluation of tissues: Kidneys, lymph nodes and spleens were harvested from IL-14β transgenic and littermate control mice and placed in 10% formalin (Baxter Diagnostics). Tissues in 10% formalin were sectioned and stained with Hematoxylin and eosin and evaluated by standard light microscopy, as described (Kandeel et al., 2001).
Flow cytometry: Cells collected from tumor tissue of the mice were stained with panels composite with anti-mouse antibody CD19-APC, CD21-FITC, CD5-PE-Cy5, CD23-PE, IgD-FITC (BD Pharmingen) and IgM-PE (ABcam) and analyzed on a BD Biosciences FACSCalibur machine and Winlist software.
Determination of monoclonality of tumors from IL-14β Tg mice: The determination of the clonality of the mouse tumors was performed by high fidelity PCR using 0.1 mg of DNA extracted from normal mouse liver and spleen and transgenic mouse tumors. The primers for immunoglobulin gene rearrangement in the lymphomas from the IL-14β transgenic mice were:
| DHL | : | 5'GGAATTCGMTTTTTGTSAAGGGATCTACTACTGTG |
| J3 | : | 5' GTCTAGATTCTCACAAGAGTCCGATAGACCCTGG |
| DHR | : | 5'TTTTGYTGMTGGATATAKCACTGAG |
| J3 | : | 5' GTCTAGATTCTCACAAGAGTCCGATAGACCCTGG |
PCR was performed as described (Kppers et al., 1995) with the following protocol: Incubation 95°C 5 min then 95°C 1 min, 55°C 1 min and 72°C 2 min for 35 cycles, followed by extension at 72°C for 10 min. The PCR products were analyzed on 1.2% agarose gel electrophoresis. C57Bl/6J mouse was used as the normal control.
Northern blots: The RNA probes for the Northern blots were prepared as follows: RT-PCR using the primers CTGAGAAGCATCATTGGCCT (downstream end) and ATACACCAGGTCTTGACTG (upstream end) for IL-14β and TGATGCTTCTGTGCTCGG (downstream end) and GTATTCACCACATTCAAGCAGG (upstream end) for IL-14 were used to generate cDNA. The T7 promoter adapter was then added to these cDNA and upstream primers and adapter primer were used to produce a transcription template for antisense probes and downstream primers and adapter primer used to produce a transcription template for sense probes, according to the manufacturer’s instructions (No-cloning promoter addition kit, Ambion). Using these templates the RNA probes were generated by in vitro transcription using T7 phage RNA polymerase, according to the manufacturer’s instructions (MAXIscript in vitro transcription kit, Ambion). The probes were purified using 6% TBE-urea gels, according to the manufacturer’s instructions (Invitrogen). The probes were labeled with biotin using the BrightStar Psoralen-Biotin Nonisotopic Labeling kit, according to the manufacturer’s instructions (Ambion).
Total RNA was purified from Namalva Cells before and after stimulation with 10 μg mL-1 PHA for 8 h at 37°8 in RPMI/5% FBS, as previously described.
RNA was separated in denaturing gels containing formaldehyde for 2 h and then transferred to positively charged nylon membranes (Ambion), as described.
Hybridization of filters with sense or antisense probes was performed according to the NorthernMax system for Northern blots (Ambion). The signals of the probes were detected with the BioDetect Kit, according to the manufacturer’s instructions (Ambion).
Semi-quantitative RT-PCR: The analysis of IL-14β mRNA in the peripheral blood of normal human donors was performed as follows. Total RNA was purified by TRIzol, according to the manufacturer’s instructions (Invitrogen). Complementary DNA was produced using reverse transcription with the Superscript first-strand synthesis system for RT-PCR, according to the manufacturer’s instructions (Invitrogen). The analysis of IL-14β mRNA involved PCR with the primers upstream-AGTCAGTGGCTTGTCTGTGAGC and downstream-TCCTGACCAGCACAT CATCC. The primers for the actin control were 5’ -GTGGGGCGCCCCAGGCACCA and 3’-CTCCT TAATGTCACGCACGATTTC. The following conditions for PCR were used for both sets of primers: 3 min of denaturation at 94°C, followed by 35 cycles of annealing at 58°C for 1 min, extension at 72°C for 0.5 min and denaturation at 94°C for 1 min. At the end of the 35 cycles, the reactions were maintained at 72°C for 5 min and then stored at 4°C.
RT-PCR products were separated on 1% agarose gels and visualized by incorporation of ethidium bromide with UV light.
RESULTS AND DISCUSSION
Growth of follicular lymphoma cells in vitro: Previous studies had demonstrated that IL-14 is an autocrine growth factor for various high-grade B cell lymphomas (Ford et al., 1992, 1995). Interestingly, mRNA for IL-14 was not regularly identified in purified B cells from patients with follicular lymphoma but was found in the T cells surrounding them (Ford et al., 1992). We therefore, sought to determine whether IL-14 might act as a growth factor for purified B cells from patients with follicular lymphoma. Follicular lymphoma B cells generally die rapidly when placed in culture with normal media. Figure 1 shows a representative growth curve from 6 patients studied. As can be seen, when 1000 purified B cells are placed in culture with standard media (RPMI/10% FBS), they begin to die with 24 h and are completely gone in 10 days. On the other hand, when similar cells were placed in culture with standard media plus supernatant from Namalva cells containing both IL-14α and IL-14β, there was an initial expansion of the follicular lymphoma B cells followed by a slow decline in their numbers. There were still viable B cell lymphoma cells at 20 days of culture. These studies did not distinguish between IL-14α and IL-14β.
Later studies demonstrated that IL-14α transgenic mice do not get follicular lymphoma but do get large B cell lymphomas in the setting of autoimmunity (Ford et al., 2007; Shen et al., 2006). We were therefore, interested in evaluating the physiological effects of IL-14β in transgenic mice.
Il-14β-transgenic mice: We produced IL-14β transgenic mice (IL-14β Tg) utilizing pE μSR and the human cDNA for IL-14β, by exactly the same strategy as was utilized to produce IL-14α transgenic mice (IL-14α Tg) (Shen et al., 2006).
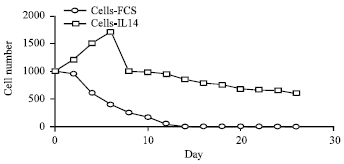 | |
| Fig. 1: | Growth of B cells from a patient with follicular lymphoma in vitro using IL-14 |
As with the IL-14α Tg mice, IL-14β was expressed exclusively in B cells and IL-14β protein could be found in the sera of the mice (data not shown). In addition, IL-14β Tg mice were bred with C-Myc Tg mice to observe the consequences of expressively both of these transcripts at high levels at the same time, as has been done with IL-14α Tg mice and C-Myc transgenic mice (Ford et al., 2007).
The IL-14β Tg mice lived a normal life span, without evidence of disease until they were autopsied, at 20-22 months of age at the latest. When evaluated at 6 and 12 months of age, the blood counts of IL-14β were normal. Serum immunoglobulins were normal. Autoantibodies, including ANA, anti-histone and anti-cardiolipin antibodies were identified in less than 20% of the mice (data not shown) (Shen et al., 2006).
Because IL-14α Tg mice demonstrated enhanced responses to T-dependent and T-independent antigens (Shen et al., 2006), we vaccinated IL-14β Tg mice with NP-OVA (T-dependent) and NP-Ficoll (T-independent). The responses to T-dependent and T-independent antigens of IL-14β Tg mice were no different than littermate controls (data not shown).
However, IL-14β Tg mice developed tumors in old age. They were B cell lymphomas that occurred in the spleen and/or gastrointestinal tract (Fig. 2a). The tumors had a nodular appearance on histology that was consistent with follicular lymphomas (Fig. 2b). Flow cytometry of the tumors revealed that they were CD5–, CD19+, CD21–, CD22+, CD23–, CD79a+, sIgM+ (data not shown) (Ford et al., 2007; Shen et al., 2006). The tumors from the IL-14β Tg mice expressed bcl-2 constitutively while the tumors of the IL-14α Tg mice did not (Ford et al., 2007) (Fig. 2c). The tumors of the IL-14β Tg mice were monoclonal (Fig. 2d). The tumors of the IL-14β Tg mice therefore, most closely resemble human follicular lymphoma.
Because the breeding of IL-14α Tg mice with C-Myc transgenic mice had resulted in Double Transgenic mice (DTgα) that rapidly developed aggressive B cell lymphomas (Ford et al., 2007), we also crossed C-Myc transgenic with IL-14β Tg mice to get DTgβ mice. The DTgβ mice develop fatal aggressive B cell lymphomas by 2 months of age and in fact die earlier than DTgα mice. An autopsy of a typical DTgβ transgenic mouse at 2 months of age is shown in Fig. 3a. Large blastic B cells are present in the blood stream, bone marrow, lymph nodes, spleen, liver, gastrointestinal tract and kidney of these mice (Fig. 3b). Flow cytometry demonstrated that the DTgβ tumors are CD19+, surface IgM+ but CD5, CD21 and CD23 negative (Fig. 3c). DTgβ tumors express bcl-2, similar to tumors of IL-14β Tg mice (Fig. 2c). These tumors are most consistent with the blastic phase of follicular lymphoma, often rapidly fatal in patients.
Expression of IL-14β: Because the physiological effects of transgenic expression of IL-14β were so different that IL-14α, we determined the expression of IL-14β in various normal and abnormal cells. Because Interleukin 14 was initially identified as a product of activated Namalva cells, a Burkitt lymphoma cell line (Ambrus and Fauci, 1985) we first evaluated the differential expression of IL-14α and IL-14β in these cells (Fig. 4). Northern blot analysis of mRNA from Namalva cells done using strand specific probes based on exon 10 revealed that resting Namalva cells produced an mRNA from the negative strand that was slightly larger than the 18S marker (Fig. 4). This is IL-14β. After activation of Namalva cells with PHA, the amount of this mRNA was increased. On the other hand, the transcript produced from the positive strand of the IL-14 gene was identified only after PHA stimulation of Namalva cells and was greater than 28 S. The transcript produced from the positive strand is IL-14α which has been identified in follicular dendritic cells, activated T cells and a subpopulation of activated B cells (Ambrus et al., 1993; Shen et al., 2006).
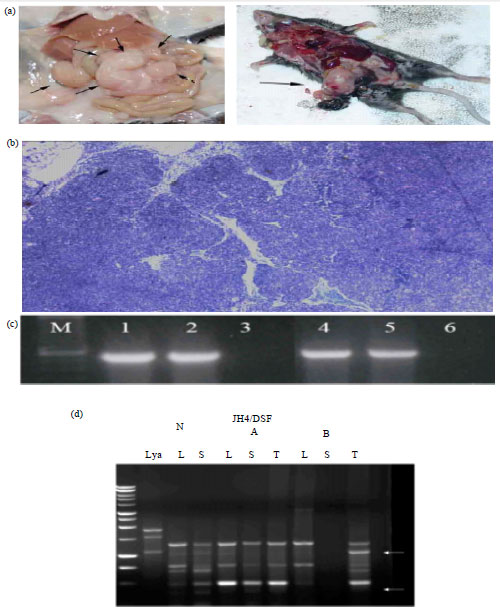 | |
| Fig. 2(a-d): | (a) Lymphoma in IL14 transgenic mice: Autopsies of two IL14 transgenic mice are shown at 20 months of age. The mouse on the left shows tumor in the gastrointestinal tract while the mouse on the right shows tumor in the spleen, (b) Histology of a tumor from an IL14 transgenic mouse: A characteristic tumor infiltrated lymph node from an IL-14 transgenic mouse is shown after staining with H and E, as outlined in materials and methods, (c) Expression of bcl-2 by a tumor from an IL14 -transgenic mouse and a tumor from an IL14 x c-Myc double transgenic mouse (DTG ): Data shown are the ethidium bromide gels of the DNA fragments from RT-PCR obtained for actin (Lanes 1, 2) and bcl-2 (Lanes 4,5) for an IL-14 TG tumor (Lane 1 and 4) and a DTG tumor (Lane 2 and 5). Lane 3 and 6 are negative controls that included all the reagents except for the template, (d) Monoclonality of a tumor from an IL14 transgenic mouse: Data shown represent DNA prepared from the liver (L), spleen (S) and tumor (T) of IL14 transgenic mice. Two mice with tumors (A and B) and a normal mouse (N) were evaluated. The first row on the left of the gel is molecular weight markers and the second lane (Lya) is a lymphoma used as a positive control. Arrows indicate unique bands indicating monoclonality of the tumor |
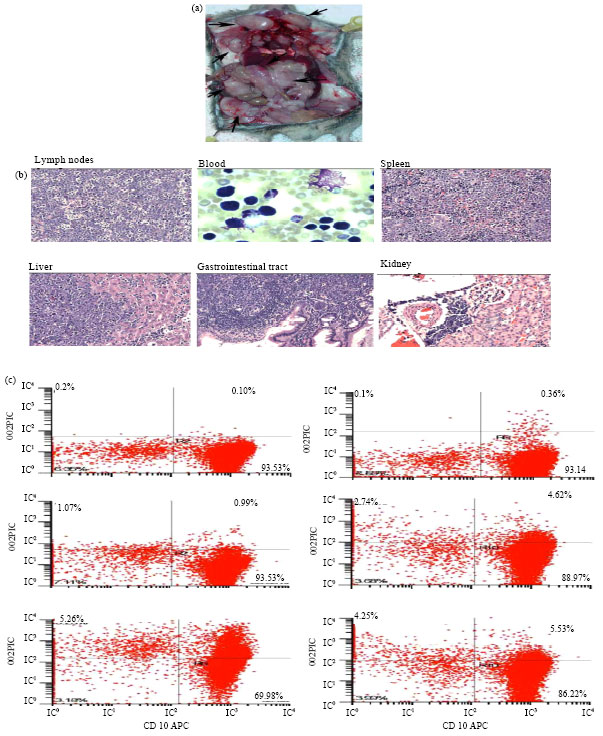 | |
| Fig. 3(a-c): | (a) Lymphoma in a DTG mouse: The autopsy of a typical DTG mouse at 2 months of age is shown, (b) Histology of a tumor from a DTG mouse: The histological evaluation of a typical DTG transgenic mouse at 2 months of age is shown. Demonstrated are H and E stains of lymph nodes, spleen, liver, gastrointestinal tract and kidney, as indicated. A Wright stain of peripheral blood is shown, (c) In all panels the x-axis shows staining with anti-CD19: The y-axis for the various panels shows staining with the following reagents: top right anti-CD21, top middle anti-CD23, top left anti-CD5, bottom left anti-IgD, bottom middle anti-IgM and bottom right anti-CD5 |
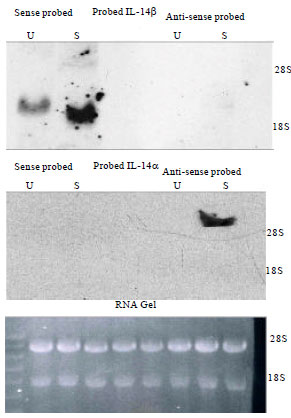 | |
| Fig. 4: | Expression of IL-14α and IL-14β in Namalva cells: Data shown are northern blots. U: Unstimulated Namalva mRNA and S: mRNA collected after Namalva cells after stimulation with PHA for 8 h. The top panel utilizes strand-specific probes that recognize a region of exon 10 present only in the IL-14β mRNA. The sense probe is shown on the left and the anti-sense probe on the right. The middle panel utilizes strand-specific probes recognizing a region of exons 4-7 present only in IL-14α mRNA. The sense probe is shown on the left and the anti-sense probe on the right. The bottom panel shows the ethidium bromide staining of the RNA utilized in the 8 lanes of the Northern blots shown in the top and middle panels in order from left to right |
The transcript produced from the minus strand is IL-14β and is constitutively expressed in most germinal center cells (Fig. 5a). Interestingly while IL-14α is highly expressed in the spleens of MRL lpr and (NZBxNZW) F1 mice (Shen et al., 2006), murine models of SLE, the expression of IL-14β was decreased in the spleens of these animals (Fig. 5b).
The IL-14 gene expresses transcripts from both the positive and negative strands. The expression of these two transcripts is differentially regulated (Fig. 4, 5). The expression of IL-14β is constitutive in B cells, T cells and follicular dendritic cells while the expression of IL-14α occurs only after cell activation. The IL-14β Tg mice demonstrate no significant immunological abnormalities until they are very old, 20-22 months of age. At this age the majority of the mice develop a tumor that closely resembles human follicular lymphoma (Fig. 2). When IL-14β Tg mice are bred with C-Myc Tg mice, the DTgβ develop vary rapid tumors most closely resembling the blastic phase of follicular lymphoma (Fig. 3). The ability of IL-14 to support the growth of B cells from patients with follicular lymphoma in vitro (Fig. 1) was likely related to the presence of IL-14β.
It is currently understood that both strands of eukaryotic DNA may be transcribed for various purposes (Yelin et al., 2003). In the case of the immunoglobulin genes, transcripts from the anti-sense strand participate in the regulation of somatic hyper-mutation and class switch recombination (Perlot et al., 2008).
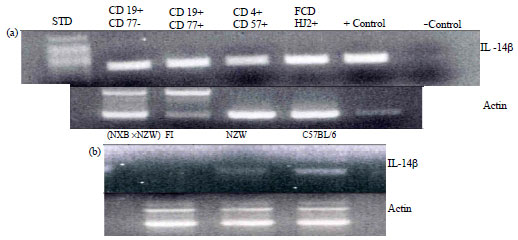 | |
| Fig. 5(a-b): | (a) Expression of IL-14β in germinal center B cells, T cells and follicular dendritic cells: Ethidium bromide gel of the DNA fragments produced from RT-PCR reactions for extra-follicular B cells (CD19+, CD77–), germinal center B cells (CD19+, CD77+), follicular T cells (CD4+, CD57+) and follicular dendritic cells (HJ2+) isolated from human tonsils. The positive control is mRNA from activated Namalva cells and the negative control contains all the reagents without the primers for the RT-PCR reaction, (b) Lack of constitutive expression of IL-14β in Autoimmune (NZBxNZW) F1 Mice: The figure demonstrates the ethidium bromide gel of the DNA fragments produced from RT-PCR reactions evaluating IL-14β in female 8 week-old C57BL/6, NZW and (NZBxNZW) F1 mice |
Many of the antisense transcripts may have regulatory roles regarding the transcription and translation of one or several sense transcripts (Storz, 2002). Abnormal antisense transcripts have been identified as the cause of genetic disease (Tufarelli et al., 2003). The IL14 gene represents an interesting phenomenon as both sense and antisense DNA are transcribed and over expression of either of these transcripts leads to a clinical phenotype. Increased expression of the IL-14α transcript results in autoimmune disease and large B cell lymphoma (Shen et al., 2006, 2009, 2010). Increased expression of the IL-14β transcript does not result in autoimmune disease but does result in lymphoma very similar to follicular lymphoma (Fig. 2, 3). The expression of IL-14α is increased in both human and murine autoimmune diseases, correlating with its ability to induce autoimmune disease in transgenic mice (Shen et al., 2009, 2006). The expression of IL-14β is decreased in (NZBxNZW) F1 mice (Fig. 5b), an animal model for SLE. The expression of IL-14β is constitutive in germinal center B cells, T cells and follicular dendritic cells (Fig. 5a) but does not lead to increased antibody or memory B cell responses. On the other hand, expression of IL-14α increases in these cells when antigen-specific responses occur (Shen et al., 2006). It is possible that IL-14β plays a role in regulating the expression of IL-14α under these circumstances but this is currently unknown. It is an area of active investigation.
The development of Follicular Lymphoma (FL) in IL-14β Tg mice may be related to the increased expression of bcl-2 in response to IL-14 (Fig. 2) (McDonnell et al., 1989). Follicular lymphoma is characterized by the t (14; 18) translocation involving the bcl-2 gene (Vitolo et al., 2008). Follicular lymphoma is the most common indolent B cell non-Hodgkin lymphoma (Van Besien and Schouten, 2007; Vitolo et al., 2008). Histologically it consists of follicles containing populations of neoplastic centrocyte- and centroblast-like lymphoma cells, involving lymphoid Germinal Centers (GC) (Tan and Horning, 2008). The ontogeny and pathogenesis of FL cells remains unclear, although it appears FL appears to involve long-lived Germinal Center (GC) neoplastic B cells, most of which have undergone Somatic Hypermutation (SHM) and Class-switch Recombination (CSR) (Vitolo et al., 2008). The clinical behavior of FL is presumed to be determined by the immuno-regulatory network within the FL lesion (Biagi and Seymour, 2002). The only animal model for follicular lymphoma involves the expression of bcl-2 with a Vav promoter (Egle et al., 2003). The availability of the IL-14β Tg mouse will allow additional studies to be done examining the pathophysiology of follicular lymphoma.
The DTgβ mouse provides another interesting tumor model (Fig. 3). The tumor is histologically and genetically consistent with the blastic phase of follicular lymphoma, a tumor for which there is no other animal model (De Jong and de Boer, 2009). The constitutive expression of Myc in this model results in animals that are born with tumor cells in their blood stream and succumb to overwhelming tumors by two months of age. Myc is known to participate in the development of a wide range of tumors by several mechanisms including increasing cell cycle entry (Adhikary and Eilers, 2005). The development of a tumor requires the dysregulation of a gene increasing cell cycle entry (Myc), a gene inducing proliferation (IL-14) and a gene inhibiting apoptosis (bcl-2; up regulated by IL-14) ( Rui and Goodnow, 2006; Sanchez-Beato et al., 2003; Staudt and Dave, 2009; Khan and Jamil, 2008). Interestingly, both IL-14α and IL-14β can participate independently in this process.
In conclusion, the IL-14β transcript and protein are unusual in that they are produced from the opposite strand of DNA opposite from another cytokine, IL-14α. Aberrant expression of IL-14β results in lymphoma closely resembling follicular lymphoma. Further study will be necessary to examine the regulation of IL-14β and to understand how it contributes to lymphomagenesis.
REFERENCES
- Abdel-Fattah, M.M. and O.G. Yassine, 2006. Non-Hodgkin's lymphomas in alexandria, Egypt: Incidence rates and trend study (1995-2004). Int. J. Cancer Res., 2: 345-357.
CrossRefDirect Link - Adhikary, S. and M. Eilers, 2005. Transcriptional regulation and transformation by Myc proteins. Nat. Rev. Mol. Cell Biol., 6: 635-645.
CrossRef - Ambrus, Jr. J.L., J. Pippin, A. Joseph, C. Xu and D. Blumenthal et al., 1996. Identification of a cDNA for a human high molecular-weight B-cell growth factor. Proc. Natl. Acad. Sci. USA., 93: 8154-8154.
Direct Link - Ambrus, Jr. J.L. and A.S. Fauci, 1985. Human B lymphoma cell line producing B cell growth factor. J. Clin. Invest., 75: 732-739.
CrossRef - Aurora, V. and J.N. Winter, 2006. Current controversies in follicular lymphoma. Blood Rev., 20: 179-200.
PubMed - Biagi, J.J. and J.F. Seymour, 2002. Insights into the molecular pathogenesis of follicular lymphoma arising from analysis of geographic variation. Blood, 99: 4265-4275.
CrossRefPubMedDirect Link - De Jong, D. and J.P. de Boer, 2009. Predicting transformation in follicular lymphoma. Leuk Lymphoma, 50: 1406-1411.
PubMed - De Jong, D., 2005. Molecular pathogenesis of follicular lymphoma: A cross talk of genetic and immunologic factors. JCO, 23: 6358-6363.
CrossRef - Egle, A., A.W. Harris, M.L. Bath, L. O'Reilly and S. Cory, 2003. VavP-Bcl2 transgenic mice develop follicular lymphoma preceded by germinal center hyperplasia. Blood, 103: 2276-2283.
CrossRefPubMedDirect Link - Ford, R., A. Tarnayo, B. Martin, K. Niu, K. Claypool, F. Cabanillas and J. Jr. Arnbrus, 1995. Identification of B-cell growth factors (interleukin-14; high molecular weight-B-cell growth factors) in effusion fluids from patients with aggressive B-cell lymphomas. Blood, 86: 283-295.
Direct Link - Ford, R.J., A. Tamayo and J.L. Jr. Ambrus, 1992. The role of growth factors in human lymphomas. Curr.Top Microbiol. Immunol., 182: 341-347.
PubMed - Ford, R.J., L. Shen, Y.C. Lin-Lee, L.V. Pham and A. Multani et al., 2007. Development of a murine model for blastoid variant mantle cell lymphoma. Blood, 109: 4899-4906.
CrossRefDirect Link - Harris, N.L., E.S. Jaffe, H. Stein, P.M. Banks and J.K. Chan et al., 1994. A revised European-American classification of lymphoid neoplasms: A proposal from the International Lymphoma study group. Blood, 84: 1361-1392.
PubMedDirect Link - Hoglund, M., L. Sehn, J.M. Connors, R.D. Gascoyne and R. Siebert et al., 2004. Identification of cytogenetic subgroups and karyotypic pathways of clonal evolution in follicular lymphomas. Genes, Chromosomes Cancer, 39: 195-204.
CrossRef - Husson, H., E.G. Carideo, D. Neuberg, J. Schultze and O. Munoz et al., 2002. Gene expression profiling of follicular lymphoma and normal germinal center B cells using cDNA arrays. Blood, 99: 282-289.
CrossRef - Kandeel, A.A., M. Zeid, T. Helm, M.A. Lillie, E. Donaheu and J.L. Ambrus, 2001. Evaluation of chronic urticaria in patients with Hashimotos thyroiditis. J. Clin. Immunol., 21: 335-347.
CrossRef - Khan, M. and J. Kaiser, 2008. Genomic distribution, expression and pathways of cancer metasignature genes through knowledge based data mining. Int. J. Cancer Res., 4: 137-145.
CrossRefDirect Link - Kppers, R., M.L. Hansmann, V. Diehl and K. Rajewsky, 1995. Molecular single-cell analysis of hodgkin and reed-sternberg cells. Mol. Med. Today, 1017: 26-30.
Direct Link - Kuppers, R., 2005. Mechanisms of B-cell lymphoma pathogenesis. Nat. Rev. Cancer, 5: 251-262.
CrossRefDirect Link - Limpens, J., D. de Jong, J.H. van Krieken, C.G. Price, B.D. Young, G.J. van Ommen and P.M. Kluin, 1991. Bcl-2/JH rearrangements in benign lymphoid tissues with follicular hyperplasia. Oncogene, 6: 2271-2276.
PubMed - McDonnell, T.J. and S.J. Korsmeyer, 1991. Progression from lymphoid hyperplasia to high-grade malignant lymphoma in mice transgenic for the t(14;18). Nature, 349: 254-256.
CrossRefPubMedDirect Link - McDonnell, T.J., N. Deane, F.M. Platt, G. Nunez, U. Jaeger, J.P. McKearn and S.J. Korsmeyer, 1989. bcl-2-immunoglobulin transgenic mice demonstrate extended B cell survival and follicular lymphoproliferation. Cell, 57: 79-88.
PubMed - Perlot, T., G. Li and F.W. Alt, 2008. Antisense transcripts from immunoglobulin heavy-chain locus V(D)J and switch regions. PNAS, 105: 3843-3848.
CrossRef - Roulland, S., F. Suarez, O. Hermine and B. Nadel, 2008. Pathophysiological aspects of memory B-cell development. Trends Immunol., 29: 25-33.
PubMed - Rui, L. and C.C. Goodnow, 2006. Lymphoma and the control of B cell growth and differentiation. Curr. Mol. Med., 6: 291-308.
PubMed - Sanchez-Beato, M., A. Sanchez-Aguilera and M.A. Piris, 2003. Cell cycle deregulation in B-cell lymphomas. Blood, 101: 1220-1235.
PubMed - Shen, L., C. Zhang, T. Wang, S. Brooks and R.J. Ford et al., 2006. Development of autoimmunity in interleukin 14 alpha transgenic mice. J. Immunol., 177: 5676-5686.
PubMedDirect Link - Shen, L., L. Suresh, H. Li, C. Zhang, V. Kumar, O. Pankewycz and J.L. Ambrus Jr., 2009. IL-14 α, the nexus for primary Sjogren's disease in mice and humans. Clin. Immunol., 130: 304-312.
PubMed - Shen, L., L. Suresh, H. Li, C. Zhang, V. Kumar, O. Pankewycz and J.L. Ambrus Jr., 2010. A role for lymphotoxin in primary sjogren's disease. J. Immunol., 185: 6355-6363.
PubMedDirect Link - Tan, D. and S.J. Horning, 2008. Follicular lymphoma: Clinical features and treatment. Hematol. Oncol. Clin. North Am., 22: 863-882.
PubMed - Van Besien, K. and H. Schouten, 2007. Follicular lymphoma: A historical overview. Leuk Lymphoma, 48: 232-243.
PubMed - Vitolo, U., A.J.M. Ferreri and S. Montoto, 2008. Follicular lymphomas. Crit. Rev. Oncol. Hematol., 66: 248-261.
CrossRefDirect Link - Yelin, R., D. Dahary, R. Sorek, E.Y. Levanon and O. Goldstein et al., 2003. Widespread occurrence of antisense transcription in the human genome. Nat. Biotechnol., 21: 379-386.
CrossRef