
Feroz Khan
Division of Biochemical Sciences, National Chemical Laboratory, Pune 411 008, India
M. Islam Khan
Division of Biochemical Sciences, National Chemical Laboratory, Pune 411 008, India
International Journal of Biological Chemistry
Year: 2011 | Volume: 5 | Issue: 1 | Page No.: 1-20







ABSTRACT
The aim of this study is to provide concise information about recent developments in understanding of fungal lectins. Lectins, a well-known class of multivalent carbohydrate binding proteins of non-immune origin that recognize diverse sugar structures with a high degree of stereospecificity in a non-catalytic manner are wide spread in distribution. Plant and animal lectins are subjected to extensive studies and only limited information was available on fungal lectins. In last few years mushroom and other fungal lectins have attracted wide attention due to their antitumor, antiproliferative and immunomodulatory activities. Earlier fungal lectin reports deal only with their purification, carbohydrate specificity, basic characterization and possible roles. In last ten years, several fungal lectins have been cloned, sequenced and crystallized. More subtle information about their structure and binding properties is available, obtained by employing more advanced techniques such as X-ray crystallography, surface plasmon resonance and enzyme linked lectinsorbent assay etc. Several fungal lectins have been discovered in the recent years and their structural and biochemical properties have been explored. However, some of them show resemblance with plant and bacterial lectins, but still there are enough evidence to place them in a diverse lectin group. This article will provide concise information about recent advancement in understanding of fungal lectins regarding their biochemical and molecular properties.
PDF Abstract XML References Citation
Received: May 06, 2010;
Accepted: May 21, 2010;
Published: August 30, 2010
How to cite this article
Feroz Khan and M. Islam Khan, 2011. Fungal Lectins: Current Molecular and Biochemical Perspectives. International Journal of Biological Chemistry, 5: 1-20.
URL: https://scialert.net/abstract/?doi=ijbc.2011.1.20
URL: https://scialert.net/abstract/?doi=ijbc.2011.1.20
INTRODUCTION
Lectins, a well-known class of multivalent carbohydrate binding proteins of non-immune origin which recognize diverse sugar structures with a high degree of stereospecificity in a non-catalytic manner are wide spread in distribution (Sharon and Lis, 1989).
Lectins have been implicated in cellular signaling, malignancy, host pathogen interactions, scavenging of glycoproteins from the circulatory system, cell-cell interactions in the immune system, differentiation and protein targeting to cellular compartments (Ashwell and Harford, 1982; Sharon and Lis, 1989; Springer and Lasky, 1991). Plant and animal lectins are subjected to extensive studies (Rini and Lobsanov, 1999; Rudiger and Gabiu, 2001; Occena et al., 2007; Tanaka et al., 2009; Fujii et al., 2009; Kaur et al., 2006) and very little information is available on lectins from fungi (Guillot and Konska, 1997; Konska, 2006; Wang et al., 1998). However, first fungal lectin phallin was reported by Kobert in 1891 from Amanita phalloides, which was a hemolytic agent (Horejsi and Kocourek, 1978) and later a fungal hemagglutinin was discovered in the fly agaric (Ford, 1910). In last few years mushroom and other fungal lectins have attracted wide attention due to their antitumor, antiproliferative and immunomodulatory activities (She et al., 1998; Wang et al., 2000; Waiser and Weis, 1999). More recently, there are several reports on lectins from lower and pathogenic fungi but their physiological role still remains uncertain (Candy et al., 2001, 2003; Khan et al., 2007c; Tronchin et al., 2002; Wu et al., 2001).
There are few reviews about fungal and mushroom lectins (Guillot and Konska, 1997; Konska, 2006; Singh et al., 2010; Wang et al., 1998) till date, mainly dealing about their taxonomic and biological perspectives. This article will provide concise but exclusive information regarding advancement in understanding of fungal lectins about their biochemical and molecular aspects as proteins.
Relevant abstracts and references were collected from PubMed (http://www.ncbi.nlm.nih.gov/pubmed) and Science Alert (http://www.scialert.net/index.php). Sequence alignment studies were carried out by obtaining amino acid sequences of the lectins from NCBI (http://www.ncbi.nlm.nih.gov/protein) and aligned by ClustalW2 (http://www.ebi.ac.uk/Tools/clustalw2/ index.html).
FUNGAL LECTINS
Occurrence and localization: Fungal lectins have been isolated from mycelium (Candy et al., 2003; Khan et al., 2007c), conidia (Tronchin et al., 2002), sporomes (Guillot and Konska, 1997) basidiomes (Guillot and Konska, 1997) and fruiting bodies (Thakur et al., 2007b; Wang and Ng, 2003) whereas, lectin from pathogenic fungus Macrophomina phaseolina is extracellular in nature (Bhowal et al., 2005). In case of few strains of Rhizoctonia solani, the amount of lectin in the sclerotia was higher than in the mycelium (Hamshou et al., 2007).
Detection and assay: Lectins were earlier discovered as hemagglutinating agents and hemagglutination is still a universally adopted and easiest detection technique of the lectin activity in crude or purified protein extracts. The hemagglutination assay is carried out using animal as well as human erythrocytes (Khan et al., 2007c; Thakur et al., 2007b). Sometimes erythrocytes are treated with enzymes like pronase, trypsin, papain and neuraminidase or any other proteolytic enzyme (Khan et al., 2007c; Thakur et al., 2007b). Other types of cells like lymphocytes, sperms and yeast have also been used for lectin detection (Nicolson, 1974). Lectins also form cross-links between polysaccharide (Chien et al., 1975; Van Wauwe et al., 1973) or glycoproteins (Yachnin, 1972, 1975) in solution and induce their precipitation.
Purification: Since, majority of fungal lectins are intracellular, most of the purification procedures, involves lysis of cells by some physical method, followed by conventional protein purification techniques such as chromatography using various resins.
Ion-exchangers like DEAE- CM-cellulose and QAE-Toyopearl, most readily available resins, have been used for the purification of some fungal lectins (Kossowska et al., 1999; Li et al., 2008; Zhao et al., 2009). Gel filtration has been used as one of the purification steps for lectins from Rhizopus stolonifer (Oda et al., 2003) and Aspergillus fumigatus (Tronchin et al., 2002). In addition, hydrophobic matrix like Phenyl-Sepharose has been employed for the purification of lectins from A. fumigatus (Tronchin et al., 2002), Beauveria Bassiana (Kossowska et al., 1999) and Fusarium solani (Khan et al., 2007c).
Affinity chromatography, as more specific technique for lectin purification, has been extensively used for purification of fungal lectins. Several affinity resins have been generated and used according to the lectin specificity such as fetuin-Sepharose (Bhowal et al., 2005), BSM-Toyopearl (Kawagishi et al., 1997), PSM-Sepharose (Chumkhunthod et al., 2006), lactosyl-Sepharose (Pohleven et al., 2009) and gal-Sepharose (Candy et al., 2003). In some cases, Sephadex and Sepharose or acid treated Sepharose have been used to purify glucopyranosyl/mannopyranosyl and galactopyranoside binding lectins respectively, where they serve as affinity matrix and not as molecular sieves. Sepharose was used for the isolation of a lectin from Ischnoderma resinosum, specific for methyl β-galactoside (Kawagishi and Mizuno, 1988). The intracellular lectin, calnexin, from Aspergillus oryzae was purified from conjugating synthetic N-linked glycan to Sepharose beads (Watanabe et al., 2007). In some cases, ability of lectins to agglutinate erythrocytes has been utilized for their purification, by embedding erythrocytes or erythrocyte stromas in polyacrylamide (Guillot et al., 1983).
Modern purification techniques like HPLC and FPLC have also been employed for the purification of lectins from Hygrophorus hypothejus (Veau et al., 1999), Ganoderma capense (Ngai and Ng, 2004), Peziza sylvestris (Wang and Ng, 2005) and Pholiota adiposa (Zhang et al., 2009). Recombinant lectin of Agrocybe aegerita is expressed in E. coli based expression system with addition of the Histidine tag and purified using Ni-column chromatography (Yang et al., 2005a).
Molecular mass and subunit structure: Molecular masses of fungal lectins range from 15-90 kDa, but majority of them are between 23-36 kDa (Table 1). In general, most of them are dimeric proteins and subunits are held together by non-covalent interactions, with few exceptions such as Phallus impudicus (Entlicher et al., 1985) and Lactarius lignyotus (Sychrova et al., 1985), where subunits are linked together by disulphide linkages.
| Table 1: | Physical properties of some fungal lectins |
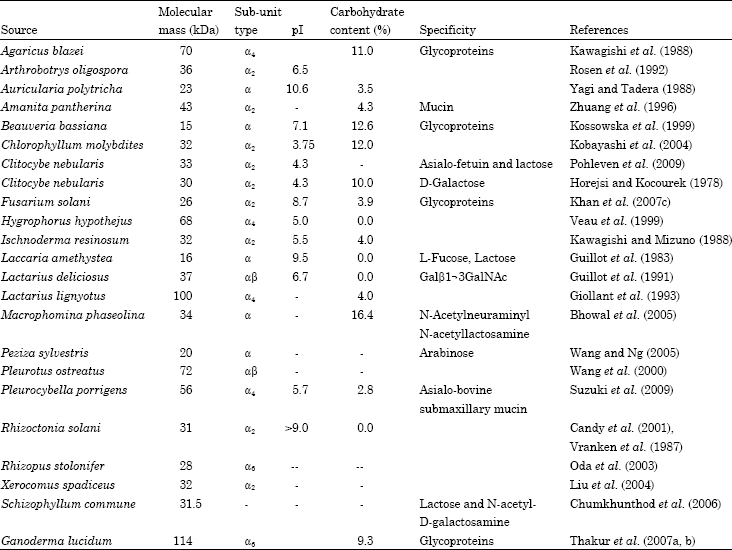 | |
Lectin L1 from Lentinus edodes is monomeric having molecular mass 45 kDa (Tsivileva et al., 2008). A lectin, CML from Cordyceps militaris is also monomeric with a molecular mass 31 kDa (Jung et al., 2007). The lectins of Pleurocybella porrigens (Suzuki et al., 2009) and Agaricus blazei (Kawagishi et al., 1988) are homotetrameric, whereas, Lactarius rufus lectin (Panchak and Antoniuk, 2007) and Rhizopus stolonifer (Oda et al., 2003) are hexameric. The lectin of Hericium erinaceum is a heterotetramer with molecular mass of 54 kDa and has two different subunits with molecular mass of 15 and 16 kDa (Kawagishi et al., 1994) and interestingly, a lectin from Kluyveromyces bulgaricus forms octamer at high concentration (Al-Mahmood et al., 1991).
Isoelectric point: Isoelectric points of majority of fungal lectins are in the range of 5-8 (Table 1). Lectin from Chlorophyllum molybdites (Kobayashi et al., 2004) is highly acidic proteins with a pI of 3.75. On the other hand, lectins from Laccaria amethystea (Guillot et al., 1983), Auricularia polytricha (Yagi and Tadera, 1988) and Rhizoctonia solani (Candy et al., 2001) are highly basic proteins with the pI values of 9.5, 10.6 and >9, respectively.
Carbohydrate content: The carbohydrate content varies from lectin to lectin and in some cases it could be high as 30% as in Rigidoporus lignosus lectin (Guillot and Konska, 1997) while in some cases it is totally absent as in Laccaria amethystea (Guillot et al., 1983) and Laetiporus sulphureus lectins (Konska et al., 1994) (Table 1).
Metal ion requirement: Fungal lectins in general do not require metal ions for their activity. The lectin of Xerocomus chrysenteron require Mn2+ for its activity (Sychrova et al., 1985). In case of Xerocomus spadiceus lectin Zn2+ and Al3+ stimulated the activity (Liu et al., 2004) while Fe3+ stimulated the activity of Polyporus adusta lectin (Wang et al., 2003). Contrarily, Fe3+, Al3+ and Zn2+ inhibited activity of Armillaria luteo-virens lectin (Feng et al., 2006). Activity of Pholiota adiposa lectin is enhanced by Cu2+, Fe2+ and Al3+ (Zhang et al., 2009). The hemagglutinating activity of Inocybe umbrinella is inhibited by Ca2+, Mn2+ and Mg2+ but not affected by Fe3+, Zn2+, Al3+ (Zhao et al., 2009). Interestingly, hemagglutinating activity of Pleurotus citrinopileatus lectin was inhibited by HgCl2 but enhanced by AlCl3 (Li et al., 2008).
Stability and denaturation: Some lectins showed considerable stability such as Pholiota adiposa lectin, which is stable upto 50°C and in the low molarity NaOH and HCl solutions (Zhang et al., 2009). Lectin from Pleurocybella porrigens is stable below 60°C and pH range 4.5-8.0 (Suzuki et al., 2009), whereas, lectin from ascomycete Cordyceps militaris is stable between pH range 2-13 (Wong et al., 2009). The Agaricus edulis lectins showed very high thermostability and could withstand high concentration of urea and extreme pH (Eifler and Ziska, 1980). Ganoderma capense lectin is stable at 100°C for 60 min (Ngai and Ng, 2004), whereas, Armillaria luteo-virens lectin is stable up to 70°C but sensitive to high and low pH (Feng et al., 2006).
Fusarium solani lectin was studied for conformation stability at extreme pH, temperature and in the presence urea and guanidine thiocyanate. The lectin showed flexible tertiary structure at low pH but maintain secondary and quaternary structure. In the presence of urea and guanidine thiocyanate, it showed monophasic unfolding curves and its heat capacity remained constant during thermal denaturation (Khan et al., 2007a).
Fluorometric property: In Agrocybe cylindracea lectin fluorescence quenching and modification of tryptophan residues indicated that there were about two tryptophan residues in the protein and one of them might be located on the surface, while the other was buried in the hydrophobic shallow groove near the surface (Liu et al., 2008). In case of Fusarium solani lectin fluorescence quenching revealed that the single tryptophan in the subunit is 100% accessible to acrylamide but only 50% to succinimide and effect of pH on iodide and cesium induced quenching indicated that the tryptophan residue might be present in vicinity of positively charged amino acids (Khan et al., 2010).
Chemical modification: Chemical modification studies of Ischnoderma resinosum lectin showed involvement of lysine, carboxylate, arginine, tryptophan, histidine, cysteine and tyrosine in the binding activity of the lectin. Inhibitory sugar of the lectin viz. lactulose could not protect inactivation against NBS mediated modification, whereas, it protected inactivation induced by glycine ethyl ester, cyclohexane-1,2-dione, ethoxyformic anhydride and N-acetylimidazole (NAI) (Kawagishi and Mori, 1991). Tryptophan, carboxylate and tyrosine have been implicated in the binding activity of the lectin form Hericium erinaceum (Gong et al., 2004).
Chemical modification of serine/threonine and histidine showed the partial necessity of these residues for the hemagglutinating activity of Agrocybe cylindracea lectin. However, modifications of arginine, tyrosine and cysteine residues had no effect on its hemagglutinating activity (Liu et al., 2008). In case of Fusarium solani lectin only tyrosine residues were found to involve in hemagglutinating activity of the lectin and the inactivation caused by modification of tyrosine by N-acetylimidazole (NAI) was reversed by hydroxylamine (Khan et al., 2010). Tryptophan, lysine and histidine have been implicated in the hemagglutinating activity of Ganoderma lucidum lectin (Thakur et al., 2007b).
Agglutination: Lectins are recognized by their important property to agglutinate various types of cells such as erythrocytes. Some of the fungal lectins showed discrimination between erythrocytes of different blood groups. Treating erythrocytes with proteolytic enzymes can enhance their activity or modify specificity (Guillot and Konska, 1997). In some cases higher specific activity was observed with trypsinized (Konska, 1988) or pronase treated erythrocytes (Khan et al., 2007c). Some lectin showed distinction among human blood group types, e.g., Marasmius oreades lectin specifically agglutinates blood group B erythrocytes (Grahn et al., 2007).
In some cases only animal erythrocytes were agglutinated and some lectins can also discriminate among different animal erythrocytes. Lectins from few Aspergillus species were found to agglutinate human and pig erythrocytes but not sheep or goat erythrocytes (Singh et al., 2008). A lectin from Cordyceps militaris can agglutinate mouse and rat erythrocytes but not human ABO erythrocytes (Jung et al., 2007). Pleurocybella porrigens lectin showed good activity with human ABO pronase and neuraminidase treated erythrocytes, but no activity with untreated erythrocytes, it could also distinguish between mouse and rat erythrocytes (Suzuki et al., 2009). Several other erythrocytes from mammals, birds, reptiles and amphibians have also been used for hemagglutination (Kawagishi et al., 1994; Panchak and Antoniuk, 2007; Stepanova et al., 2007).
Carbohydrate specificity: Fungal lectins exhibit broad carbohydrate specificity, varying from simple sugars to glycoproteins. Peziza sylvestris (Wang and Ng, 2005) lectin is specific for arabinose while Hericium erinaceus lectin (Kawagishi et al., 1994) is specific for sialic acid (Table 1). Several techniques are used to determine carbohydrate specificity such as simple and conventional hemagglutination inhibition (Khan et al., 2007c) to more advanced glycan microarray (Pohleven et al., 2009), enzyme linked lectinsorbent assay (Wu et al., 2001) and frontal affinity chromatography (Van Damme et al., 2007). The hemagglutinating activity of Grifola frondosa lectin was inhibited only by the homogeneous preparation of a polysaccharide, linear D-rhamnan (Stepanova et al., 2007). The hemagglutinating activity of Pleurotus citrinopileatus lectin was inhibited by maltose, O-nitrophenyl-β-D-galactopyranoside, O/P-nitrophenyl-β-D-glucuronide and insulin (Li et al., 2008). Glycan microarray analysis revealed that the Clitocybe nebularis lectin recognizes human blood group A determinant GalNAcα1-3(Fucα1-2)Galβ-containing carbohydrates and GalNAcβ1-4GlcNAc (N,N'-diacetyllactosediamine) (Pohleven et al., 2009). Frontal affinity chromatography revealed that the binding site of Sclerotinia sclerotiorum agglutinin primarily accommodates a non-reducing terminal GalNAc (Van Damme et al., 2007). Among glycoproteins tested for binding, Sclerotium rolfsii lectin reacted strongly with GalNAcα1→Ser/Thr (Tn) and/or Galβ1→3GalNacα1→(Tα) containing glycoproteins such as human Tα and Tn glycophorin, asialo-BSM, asialo-PSM and asialofetuin, but its reactivity towards sialated glycoprotein was reduced significantly (Wu et al., 2001).
Some other lectins showed specificity for unusual carbohydrates, e.g. mushroom lectin Xylaria hypoxylon for inulin and xylose (Liu et al., 2006), Agrocybe Cylindracea lectin for sialic acid (Wang et al., 2002; Yagi et al., 1997), Polyporus adusta lectin for turanose (Wang et al., 2003), Peziza sylvestris lectin for arabinose (Wang and Ng, 2005), whereas hemagglutinating activity of Lyophyllum shimeiji lectin could not be inhibited by simple sugars and glycoproteins (Ng and Lam, 2002).
Interestingly, the lectin Pleurotus ostreatus was also found to have α-galactosidase activity, the enzyme activity and carbohydrate binding property was not due to same site since the lectin accepts both α- and β-glycosides, whereas the enzyme activity was restricted to the α-anomer only. Moreover, the α-galactosidase activity was inhibited by α-galactose but not by β-galactose (Brechtel et al., 2001).
Affinity constants (Ka) and thermodynamics of interactions with carbohydrates: The kinetic data for binding of lectins to carbohydrate ligands are available for few fungal lectins. Several modern techniques such as surface plasmon resonance (SPR) have been used, which involves immobilization of either lectin (Khan et al., 2007c) or ligand (Suzuki et al., 2009). Affinity constants have been determined for the Fusarium solani lectin using spectrofluorometry (Khan et al., 2007b) and surface plasmon resonance (SPR) (Khan et al., 2007c), the lectin showed very low affinity constants for mono- and oligosaccharides but showed very high affinity constants for glycoproteins such as asialo-mucin (Ka = 1.61 e10 M-1). The binding was exothermic and enthalpically driven. The association rate constants were several orders magnitude slower than diffusion controlled reactions (Khan et al., 2007c). Affinity of Ganoderma lucidum lectin for glycoproteins was also explored using SPR, it was observed that the affinity for N-glycans was two order higher than O-glycans (Thakur et al., 2007a).
In SPR studies, the dimeric Aleuria aurantia lectin (AAL) found to have 5 binding sites per subunit and one binding site in recombinant AAL had unusually high affinities towards fucose and fucose-containing oligosaccharides with KD values in the nanomolar range (Olausson et al., 2008). Comparative binding study of Aleuria aurantia lectin (AAL) and Aspergillus oryzae lectin (AOL) employing frontal affinity chromatography revealed that AOL showed 2.9-6.2 times higher affinity constants (Ka) for α-1,6-fucosylated oligosaccharides than AAL and only AAL, additionally recognized oligosaccharides which were α-1,3-fucosylated at the reducing terminal GlcNAc (Matsumura et al., 2009). Isothermal titration calorimetry (ITC) measurements carried out for CGL3, lectin from Coprinopsis cinerea, indicated strongly enthalpically driven binding for chitotriose (Walti et al., 2008).
Amino acid composition: Pleurocybella porrigens lectin showed high amounts of ASX (12.4%), Thr (11.2%), Leu (8.8%) but complete absence of Cys, Met and Lys (Suzuki et al., 2009). Similarly, the amino acid analysis of the Grifola frondosa lectin showed a greater percentage of amino acids with positively charged R groups, arginine, lysine and histidine, but complete absence of sulfur-containing amino acids, cysteine and methionine (Stepanova et al., 2007). Lectin L2 isolated from Lentinus edodes contains high amount of Asn, i.e., 42% (w/v) (Tsivileva et al., 2008). The lectin from Ischnoderma resinosum contained substantial amounts of acidic and hydroxy amino acids, glycine, valine and leucine, but minute quantities of methionine, histidine and arginine (Kawagishi et al., 1988). The Fusarium solani lectin showed high amount of Gly and Lys, but very low amount of Trp and Cys (Khan et al., 2007c).
Amino acid sequence homology: The amino acid sequence of Xerocomus chrysenteron lectin showed 69 and 64% homology with Agaricus bisporus and Arthrobotrys oligospora (Birck et al., 2004). The N-terminal sequence of Pholiota adiposa lectin showed little similarity to sequence of Agaricales lectins (Zhang et al., 2009). The amino acid sequence of Pleurocybella porrigens lectin showed similarity with ricin-B-chain (33%), lectin from Polyporus squamosus (36%) and hemagglutinin from Clostridium botulinum, HA-1 (40%) (Suzuki et al., 2009). The sequence homology and structure prediction revealed that Clitocybe nebularis lectin belongs to ricin B-like superfamily (Pohleven et al., 2009). The N-terminal sequence of Inocybe umbrinella lectin (Zhao et al., 2009), Fusarium solani lectin (Khan et al., 2007c) and Pleurotus citrinopileatus lectin (Li et al., 2008) did not show similarity to any know lectin or hemagglutinin, whereas N-terminal sequence of Pleurotus tuber-regium lectin showed some similarity to that of Agaricus bisporus lectin (Wang and Ng, 2003). The deduced amino acid sequence of Grifola frondosa lectin showed 26.1 and 22.8% homology with jacalin related plant lectins from Helianthus tuberosus and Parkia platycephala (Nagata et al., 2005). On the other hand, Sclerotinia sclerotiorum lectin showed significant similarity only to the lectin from the fungus Ciborinia camelliae but not with any other lectins (Candy et al., 2003) (Fig. 1A, B).
Structure: Aleuria aurantia was the first fungal lectin to solve the crystal structure. The crystal structure of the lectin complexed with fucose revealed that each monomer consists of a six-bladed β-propeller fold and a small antiparallel two-stranded β-sheet that plays a role in dimerization (Wimmerova et al., 2003). The six-bladed β-propeller structure was also found to have three bound fucose residues (Fujihashi et al., 2003), suggesting that the binding sites, although all very similar in geometry, do not have the same affinity for ligands. Flammulina velutipes lectin showed unique folds never before observed in lectins (Paaventhan et al., 2003) and showed structural similarity to human fibronectin and lectins from Xerocomus chrysenteron (Birck et al., 2004) and Agaricus bisporus (Carrizo et al., 2005), resembling actinoporins, a family of pore-forming toxins from sea anemones. CGL2 from Coprinus cinerea showed fold similar to galectins (Vasta et al., 2004), a large family of lectins from all classes of vertebrates (Walser et al., 2004). The lectin from Laetiporus sulphureus (Mancheno et al., 2005) showed resemblance with the ricin-B domain, a trefoil-based fold observed in many lectins and carbohydrate-binding domains and referred to as the (QxW)3 domain (Hazes, 1996).
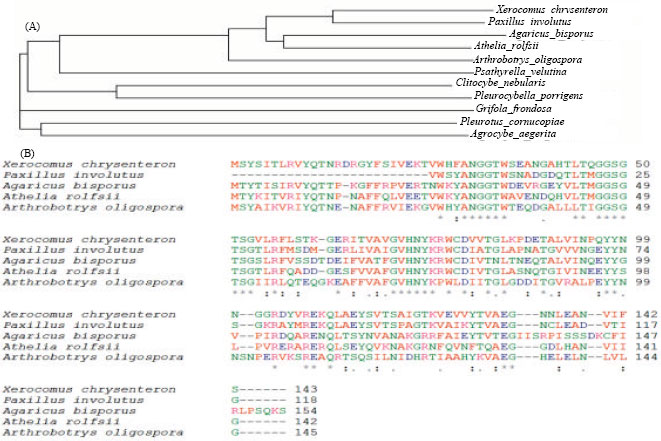 | |
| Fig. 1: | (A) Phylogenetic tree (B) and sequence alignment of some related fungal lectins. Conserved residues have been marked by stars at bottom. Amino acid sequences of the lectins were obtained from NCBI (http://www.ncbi.nlm.nih.gov/protein) and aligned by ClustalW2 (http://www.ebi.ac.uk/Tools/clustalw2/index.html) |
This lectin assembles as a trimer, each monomer consisting of a ricin-B domain and an elongated domain with structural similarity to aerolysin, a bacterial β-pore-forming toxin. This domain could be directly involved in pore formation because the lectin displays hemolytic activity (Tateno and Goldstein, 2003). The ricin-like domain displayed structural similarity to similar domains from plant toxins (ricin, abrin and mistletoe lectin), but was even more similar to the hemagglutinin component of bacterium Clostridium botulinum (Inoue et al., 2003). Structural comparison of the (QxW)3 domains from mushroom toxin, clostridial hemagglutinin and ricin illustrated their high similarity. During evolution, this domain has been conserved and often duplicated and/or combined with other domains. It has been identified in bacteria, fungi and plants and also in sponge, insects and mammals, generally conserving its role of targeting a sugar-coated substrate (Hazes, 1996).
Agrocybe aegerita lectin showed structural similarity with galectins, the carbohydrate recognition domain (CRD) of which contains consensus sequence motif, which consists of His-44, Arg-48, Val-59, Asn-61, Trp-68, Glu-71 and Arg-73 (Yang et al., 2005b), later it was found that in dimeric Agrocybe aegerita lectin each protomer adopts a prototype galectin fold (Yang et al., 2009).
Structure of Marasmius oreades agglutinin (MOA), was solved in complex with blood group B trisaccharide at 1.8 resolution. It was found that the carbohydrate ligand binds to all three binding sites of N-terminal β-trefoil domain. The structure was solved in the presence of Ca2+ which binds to MOA dimer and alters the conformation of the C-terminal domain by opening up the cleft containing a putative catalytic site (Grahn et al., 2007, 2009).
Sclerotium rolfsii lectin (SRL) crystals were grown by the hanging-drop vapor diffusion method, which belongs to the tetragonal space group P42212 group (Leonidas et al., 2003). The crystal structure the lectin in its free form and in complex with N-acetyl-D-galactosamine (GalNAc) and N-acetyl-D-glucosamine (GlcNAc) has been determined at 1.1 A, 2.0 A and 1.7 A resolution, respectively. The protein structure is composed of two beta-sheets, which consist of four and six beta-strands, connected by two alpha-helices. Sequence and structural comparisons revealed that SRL is the third member of a newly identified family of fungal lectins, which includes lectins from Agaricus bisporus and Xerocomus chrysenteron that share a high degree of structural similarity and carbohydrate specificity (Leonidas et al., 2007). Ambiguity in X-ray structure regarding Glx and Asx was solved by mass spectrometry in Sclerotium rolfsii lectin (Sathisha et al., 2008).
Crystal structure of the lectin from the mushroom Psathyrella velutina showed that it adopts a very regular seven-bladed beta-propeller fold with the N-terminal region tucked into the central cavity around the pseudo 7-fold axis (Cioci et al., 2006). Sclerotinia sclerotiorum agglutinin, based on molecular modeling, is expected to constitute a novel lectin family (Van Damme et al., 2007).
Cloning and site directed mutagenesis: The cDNA for Agrocybe aegerita lectin was prepared from total RNA using RT-PCR, cloned and expressed in BL-21 (DE3) strain of Escherichia coli (Yang et al., 2005a). The cDNA of the Pleurocybella porrigens lectin was cloned and sequenced (Suzuki et al., 2009).
Site-directed mutagenesis carried out in Aleuria aurantia lectin showed that in β-2 mutant, out of five binding sites, substitution of Tyr26 (Site 1), Tyr79 (Site 2) and Tyr181 (Site 4) with Val resulted in loss of hemagglutinating activity, whereas mutation at Tyr133 (Site 3) and Tyr228 (Site 5) did not cause any loss of activity (Amano et al., 2004). In the case of AAL (Agrocybe aegerita lectin) 11 mutants were prepared, which concluded that dimerization of the lectin is essential for its tumor cell apoptosis-inducing activity. A hydrophobic pocket consisting Leu33, Leu35, Phe93 and Ile44 was found to be involved in conferring the tumor cell apoptosis-in ducing activity. Single mutant such as Phe93Gly or Ile114Gly didn’t disrupt carbohydrate binding and homodimerization capabilities, but abolished the bioactivity of the lectin (Yang et al., 2009). Coprinopsis cinerea lectin, CGL3, showed homology with galectins with all but one conserved residues: Arg and mutation of this residue to Trp (R81W) changed its specificity from chitooligosaccharides to lactose (Walti et al., 2008).
Biological role: Lectins, due to their ability to recognize different structures, constitute an important element of biological system. Their physiological role is related with the identification of glycosylated structures at the level of cells, tissues and the whole organisms. Some of their biological roles have documented earlier, such as participate in the formation of primordia, creation of mycelium structures to facilitate, penetration of parasitic fungi into the host organism as well as mycorrhization, introducing morphological changes in host and in dormancy (Guillot and Konska, 1997; Konska, 2006).
Growth and morphogenesis: A lectin may be involved in ensuring cohesion between hyphae during the development of the basidiome as seen in Pleurotus cornucopiae (Kaneko et al., 1993). Paracoccin is an N-acetyl-glucosamine-binding lectin from Paracoccidioides brasiliensis, Immunoelectron microscopy with mouse anti-paracoccin IgG localized the antigen to the cell wall of P. brasiliensis yeast forms. Paracoccin interacted with chitin and colocalized with beta-1,4-homopolymer of GlcNAc to the budding sites of P. brasiliensis yeast cell. When the yeast cells were cultivated in the presence of anti-paracoccin antibodies, a significant reduction of both colony forming units and individual yeast cells as well as morphological alterations such as smaller colonies were observed. It was concluded that binding of specific antibodies to paracoccin may disrupt the paracoccin/chitin interactions, resulting in the inhibition of P. brasiliensis growth (Ganiko et al., 2007). Lactarius deterrimus lectin was produced by cultured mycelia and could be localized by immuno-fluorescence in the cell wall. Receptors for the lectin were localized on the root hairs of axenically grown spruce seedlings. This inferred a role of the fungal lectin in recognition and specificity during the early stages of mycorrhiza formation (Giollant et al., 1993). Activity of Lentinus edodes intracellular lectins varied at various developmental stages of the fungus, the specific activity increased in brown mycelium film stage exceeded the corresponding value for nonpigmented mycelium (Vetchinkina et al., 2008).
The role of CGL1 and CGL2, lectins from Coprinopsis cinerea, in tissue development was studied by Walser et al. (2004, 2005). The developmentally regulated ligands for galectins were co-localized with the galectin expression, in the veil surrounding the developing primordium and the outer cells of the young stipe. In addition, galectin ligands were observed in the hymenium. The subcellular localization of the galectin ligands suggested these to be present in the cellular compartments distinct from galectin transport. The sensitivity of in situ interactions with exogenous galectins towards detergents and organic solvents inferred that these ligands were lipid-borne. Accordingly, lipid fractions from primordia were shown to contain galectin-binding compounds. Based on these observations it was hypothesized that β-galactoside-containing lipids (basidiolipids) found in mushrooms are the physiological ligands for the galectins in C. cinerea (Walser et al., 2005).
It was speculated that the Aspergillus fumigatus lectin may contribute to the attachment of conidia to the extracellular matrix components through the recognition of the numerous terminal sialic acid residues of their carbohydrate chains (Tronchin et al., 2002).
Involved in pathogenesis: Paracoccin, a GlcNAc-binding lectin with a molecular mass 160 kDa, purified from human pathogen Paracoccidioides brasiliensis, bind to laminin and induced TNF-alpha production by macrophages (Coltri et al., 2006).
Molecular recognition: Presence of lectins, with strict specificities, in fungal cells of ectomycorrhizal symbiosis, suggests that lectins might be involved in recognition between the tree and its symbiont (Giollant et al., 1993).
Some of the molecules present in the soil can combine with fungal lectins and block their specific binding sites and in the rhizosphere, e.g. certain phenolic acids that can modify carbohydrate receptors on the roots of the host trees, will prevent recognition and consequently mycorrhization (Guillot et al., 1994).
TBF-1, the main soluble protein in the Tuber borchii fruiting body that is able selectively to bind the exopolysaccharides produced by ascoma-associated Rhizobium sp. was found to a phase-specific lectin, involved in molecular recognition (Cerigini et al., 2008).
Self defense against predator: Feeding of a mushroom galectin, Coprinopsis cinerea CGL2, to the namatode Caenorhabditis elegans inhibited development and reproduction and ultimately resulted in killing of the host. The lack of toxicity of a carbohydrate-binding defective CGL2 variant and the resistance of a C. elegans mutant defective in GDP-fucose biosynthesis suggested that CGL2-mediated nematotoxicity depends on the interaction between the galectin and a fucose-containing glycoconjugate. These results indicated a possible role of fungal galectins in defense of fungi against predators by binding to the specific glycoconjugates of the hosts (Butschi et al., 2010).
Applications: Ability of lectins to interact with simple, aminated, acetylated, sialated and complex carbohydrates has been exploited for typing blood cells, carrier for chemotherapeutic agents, mitogens, fractionation of animal cells, or for studying cellular surfaces. They have been used for the isolation and purification of the serum glycoconjugates, identification and differentiation of various microorganisms and in cell sorting. They have also been successfully utilized as epidemiologic as well as taxonomic markers of specific microorganism (Guillot and Konska, 1997; Slifkin and Doyle, 1990).
As biomarker: Fungal lectin have been used to study glycoconjugates, involved in the cellular interactions during the uro-genital morphogenesis in bird embryos, to understand the mechanisms responsible for the migration of germ cells. In birds, the primordial germ cells (PGCs), localized at the primitive line stage in the Swift crescent, subsequently migrate via the bloodstream towards future genital ridges. There are many arguments in favor of involvement of membrane glycoproteins in the mechanisms of recognition between PGCs and tissues during the migration. FITC-labeled lectins and NC-1/HNK-1 mono-clonal antibodies used in indirect fluorescence reactions bind to PGCs and afford ready visualization. The lectin of Laetiporus sulphureus, which recognizes N-acetyllactosamine residues is one of the most specific to this cell type (Didier et al., 1984, 1990; Fargeix et al., 1980).
Surface plasmon binding studies carried for Aspergillus oryzae lectin towards 1-fucose containing chains, indicated that it has strongest preference for the alpha1,6-fucosylated chain among alpha1,2-, alpha1,3-, alpha1,4- and alpha1,6-fucosylated pyridylaminated (PA)-sugar chains. Moreover positive staining of Aspergillus oryzae lectin, but not Aleuria aurantia lectin, was completely abolished in the cultured embryo fibroblast (MEF) cells obtained from alpha1,6-fucosyltransferase (Fut8) knock-out mice, as assessed by cytological staining, suggested that Aspergillus oryzae lectin is more suitable for detecting core fucose than Aleuria aurantia lectin or Lens culimaris agglutinin-A (Matsumura et al., 2007).
In cancer research: Clitocybe nebularis lectin elicited antiproliferative effect on human leukemic T cells (Pohleven et al., 2009). The AAL (Agrocybe aegerita lectin) possesses a potent tumor-suppressing function against several human tumor cell lines, including HeLa, SW480, SGC-7901, MGC80-3, BGC-823 and HL60, as well as the mouse sarcoma cell line S-180. It can also inhibit the viability of S-180 tumor cell in vivo. It also displayed apoptosis-inducing activities for cancer cells, indicating that the lectin exerts its antitumor abilities stemming from apoptosis-inducing activities (Zhao et al., 2003). The lectin was further found to translocate in the nucleus and induce cell apoptosis. Among the several mutants, dimer interface mutant I25G, carbohydrate recognition domain (CRD) mutant R63H and loop region mutant L33A could not enter the nucleus and lost the ability to induce apoptosis (Liang et al., 2009). Boletus satanas lectin inhibited DNA and protein synthesis in Maidn Darby canine kidney cells with an IC50 of 0.62 and 0.14 μM, respectively and the inhibitory effects could be reversed by addition of the galactose (Kretz et al., 1991).
The Pleurotus citrinopileatus lectin exerted potent antitumor activity in mice bearing sarcoma 180 and caused approximately 80% inhibition of tumor growth when administered intraperitonealy at 5 mg kg-1 daily for 20 days. It also elicited a mitogenic response from murine splenocytes in vitro with the maximal response at a lectin concentration of 2 μM (Li et al., 2008). Similarly, lectin of Pleurotus ostreatus also exerted potent antitumor activity in mice, bearing sarcoma S-180 and hepatoma H-22 cells. Survival in these mice was prolonged and body weight increase reduced after lectin treatment (Wang et al., 2000). There are several lectins which show antiproliferative activities towards several cells such as Pholiota adiposa lectin (Zhang et al., 2009), Inocybe umbrinella lectin (Zhao et al., 2009), Schizophyllum commune lectin (Han et al., 2005), Tricholoma mongolicum lectin (Wang et al., 1995) and Xerocomus spadiceus lectin (Liu et al., 2004).
The Volvariella volvacea lectin also demonstrated immunomodulatory activity as a potent stimulatory activity towards the murine splenic lymphocytes. It was also found to markedly enhance the transcriptional expression of interleukin-2 and interferon-gamma by reverse transcriptase-polymerase chain reaction. As revealed by its N-terminal amino acid sequence, it was distinct from other immunomodulatory proteins previously reported (Hsu et al., 1997) and VAG (Lin and Chou, 1984) from the same fungus.
In AIDS research: Some fungal lectins were also tested for their potential inhibitory effect against HIV. The fungal lectins from Pleurotus citrinopileatus inhibited HIV-1 reverse transcriptase with an IC50 of 0.93 μM (Li et al., 2008).
As bio-insecticide: In some cases fungal lectins were found to exhibit insecticidal activity, which renders their use as bio-insecticide. Myzus persicae a polyphagous aphid, showed no significant differences of mortality when fed with the XCL (Xerocomus chrysenteron lectin) associated with Glucose and Mannose or fed on XCL diet only. At the opposite, the mortality rates related to artificial diet supplemented with Galactose or GalNAc and XCL were significantly reduced. There was then a significant mortality difference between M. persicae fed on an artificial diet incorporated specific carbohydrate binding Lectin with those fed with lectin only, it concluded that XCL can show potential insecticidal activity in the absence of inhibitory sugars (Jaber et al., 2006, 2007, 2008).
CONCLUSIONS
The fungal lectins, reported so far, still constitute a minority among carbohydrate interacting proteins and hemagglutinins. In the light of diverse information obtained about their functions and biological roles, it is difficult to say whether they are present ubiquitously among fungi, or restricted to only some selected members among 1.5 million fungal species. Availability of more sequence information of the fungal lectin and fungal genomes, in the future will be able to make more assumptions about their origin and evolution.
On the other side, fungal lectins have also been seen as promising candidates for targeted drug delivery; however, such a use requires engineered lectin molecules with precisely defined specificity. That can be only be achieved by clubbing information for all the carbohydrate binding proteins.
ACKNOWLEDGMENTS
F.K. was supported by research fellowship by Council of Scientific and Industrial Research, India. The project was conducted as a part of doctoral thesis of FK during 2001-2007 NCL, Pune, India, while the manuscript was prepared during 2009-2010.
REFERENCES
- Al-Mahmood, S., S. Colin and R. Bonaly, 1991. Kluyveromyces bulgaricus yeast lectins. Isolation of two galactose-specific lectin forms from the yeast cell. J. Biol. Chem., 266: 20882-20887.
PubMedDirect Link - Amano, K., M. Fujihashi, A. Ando, K. Miki and Y. Nagata, 2004. Involvement of tyrosine at fucose-binding sites of Aleuria aurnatia lectin: Non-equal response to site-directed mutagenesis among five sites. Biosci. Biotechnol. Biochem., 68: 841-847.
CrossRefPubMedDirect Link - Ashwell, G. and J. Harford, 1982. Carbohydrate-specific receptors of the liver. Annu. Rev. Biochem., 51: 531-554.
CrossRefPubMedDirect Link - Birck, C., L. Damian, C. Marty-Detraves, A. Lougarre and C. Schulze-Briese et al., 2004. A new lectin family with structure similarity to actinoporins revealed by the crystal structure of Xerocomus chrysenteron lectin XCL. J. Mol. Biol., 344: 1409-1420.
CrossRefPubMedDirect Link - Brechtel, R., H. Watzig and H. Rudiger, 2001. The lectin from the mushroom Pleurotus ostreatus: A phosphatase-activating protein that is closely associated with an alpha-galactosidase activity. A part of this paper has been presented as a preliminary report at the 17th Interlec. Meeting 1997 in Wurzburg, Germany. Plant Sci., 160: 1025-1033.
CrossRefPubMed - Butschi, A., A. Titz, M.A. Walti, V. Olieric and K. Paschinger et al., 2010. Caenorhabditis elegans N-glycan core beta-galactoside confers sensitivity towards nematotoxic fungal galectin CGL2. PLoS. Pathogens, 6: e1000717-e1000717.
CrossRefPubMedDirect Link - Candy, L., E.J.M. van Damme, W.J. Peumans, L. Menu-Bouaouiche, M. Erard and P. Rouge, 2003. Structural and functional characterization of the GalNAc/Gal-specific lectin from the phytopathogenic ascomycete Sclerotinia sclerotiorum (Lib.) de Bary. Biochem. Biophys. Res. Commun., 308: 396-402.
CrossRefPubMedDirect Link - Carrizo, M.E., S. Capaldi, M. Perduca, F.J. Irazoqui, G.A. Nores and H.L. Monaco, 2005. The antineoplastic lectin of the common edible mushroom (Agaricus bisporus) has two binding sites, each specific for a different configuration at a single epimeric hydroxyl. J. Biol. Chem., 280: 10614-10623.
CrossRefPubMedDirect Link - Cerigini, E., F. Palma, E. Barbieri, M. Buffalini and V. Stocchi, 2008. The Tuber borchii fruiting body-specific protein TBF-1, a novel lectin which interacts with associated Rhizobium species. FEMS Microbiol. Lett., 284: 197-203.
CrossRefPubMedDirect Link - Chien, S.M., S. Singla and R.D. Poretz, 1975. Hapten inhibition of adsorption: Specificity of the Sophora japonica lectin. J. Immunol. Methods, 8: 169-174.
PubMed - Chumkhunthod, P., S. Rodtong, S.J. Lambert, A.P. Fordham-Skelton and P.J. Rizkallah et al., 2006. Purification and characterization of an N-acetyl-D-galactosamine-specific lectin from the edible mushroom Schizophyllum commune. Biochim. Biophys. Acta, 1760: 326-332.
PubMed - Cioci, G., E.P. Mitchell, V. Chazalet, H. Debray and S. Oscarson et al., 2006. β-propeller crystal structure of Psathyrella velutina lectin: An integrin-like fungal protein interacting with monosaccharides and calcium. J. Mol. Biol., 357: 1575-1591.
CrossRefPubMedDirect Link - Didier, E., P. Didier, J. Guillot, Y. Croisille and J.P. Thiery, 1990. Expression and distribution of carbohy-drate sequences in chick germ cells: A comparative study with lectins and the NC-1/HNK-1 monoclonal antibody. Int. J. Dev. Biol., 34: 421-431.
PubMedDirect Link - Eifler, R. and P. Ziska, 1980. The lectins from Agaricus edulis. Isolation and characterization. Experientia, 36: 1285-1286.
PubMed - Fujihashi, M., D.H. Peapus, N. Kamiya, Y. Nagata and K. Miki, 2003. Crystal structure of fucose-specific lectin from Aleuria aurantia binding ligands at three of its five sugar recognition sites. Biochemistry, 42: 11093-11099.
CrossRefPubMedDirect Link - Fujii, Y., S.M.A. Kawsar, R. Matsumoto, H. Yasumitsu, N. Kojima and Y. Ozeki, 2009. Purification and characterization of a d-galactoside-binding lectin purified from bladder moon shell (Glossaulax didyma Roding). J. Biol. Sci., 9: 319-325.
CrossRefDirect Link - Giollant, M., J. Guillot, M. Damez, M. Dusser, P. Didier, and E. Didier, 1993. Characterization of a Lectin from Lactarius deterrimus. Plant Physiol., 101: 513-522.
PubMedDirect Link - Grahn, E.M., H.C. Winter, H. Tateno, I.J. Goldstein and U. Krengel, 2009. Structural characterization of a lectin from the mushroom Marasmius oreades in complex with the blood group B trisaccharide and calcium. J. Mol. Biol., 390: 457-466.
CrossRefPubMedDirect Link - Guillot, J., L. Genaud, J. Gueugnot and M. Damez, 1983. Purification and properties of two hemagglutinins of the mushroom Laccaria amethystina. Biochemistry, 22: 5365-5369.
CrossRefDirect Link - Guillot, J., M. Giollant, M. Damez and M. Dusser, 1991. Isolation and characterization of a lectin from the mushroom, Lactarius deliciosus. J. Biochem., 109: 840-845.
PubMedDirect Link - Guillot, J., M. Giollant, M. Damez and M. Dusser, 1994. Intervention des lectines fongiques dans les evenements precoces de reconnaissance arbre/champignon au cours de la formation des ectomycorhizes. Acta Botanica Gallica, 141: 443-447.
Direct Link - Hamshou, M., G. Smagghe and E.J. Van Damme, 2007. Analysis of lectin concentrations in different Rhizoctonia solani strains. Commun. Agric. Appl. Biol. Sci., 72: 639-644.
PubMed - Hazes, B., 1996. The (QxW)3 domain: A flexible lectin scaffold. Protein Sci., 5: 1490-1501.
CrossRefPubMedDirect Link - Horejsi, V. and J. Kocourek, 1978. Studies on lectins. XXXVI. Properties of some lectins prepared by affinity chromatography on O-glycosyl polyacrylamide gels. Biochim. Biophys. Acta, 538: 299-315.
PubMed - Hsu, H.C., C.I. Hsu, R.H. Lin, C.L. Kao and J.Y. Lin, 1997. Fip-vvo, a new fungal immunomodulatory protein isolated from Volvariella volvacea. Biochem. J., 323: 557-565.
PubMedDirect Link - Inoue, K., M. Sobhany, T.R. Transue, K. Oguma, L.C. Pedersen and M. Negishi, 2003. Structural analysis by X-ray crystallography and calorimetry of a haemagglutinin component (HA1) of the progenitor toxin from Clostridium botulinum. Microbiology, 149: 3361-3370.
CrossRefPubMedDirect Link - Jaber, K., D.G. Cuartero, E. Haubruge and F. Francis, 2008. Investigation of carbohydrate binding property of a fungal lectin from Xerocomus chrysenteron and potential use on Myzus persicae aphid. Commun. Agric. Applied Biol. Sci., 73: 629-638.
PubMed - Jaber, K., F. Francis, L. Paquereau, D. Fournier and E. Haubruge, 2007. Effect of a fungal lectin from Xerocomus chrysenteron (XCL) on the biological parameters of aphids. Commun. Agric. Applied Biol. Sci., 72: 629-638.
PubMed - Jaber, K., L. Paquereau, D. Fournier, E. Haubruge and F. Francis, 2006. Use of artificial diet system to assess the potential bio-insecticide effect of a fungal lectin from Xerocomus chrysenteron (XCL) on Myzus persicae. Commun. Agric. Applied Biol. Sci., 71: 497-505.
PubMed - Kaneko, T., S. Oguri, S. Kato and Y. Nagata, 1993. Developmental appearance of lectin during fruit body formation in Pleurotus cornucopiae. J. Gen. Applied Microbiol., 39: 83-90.
CrossRefDirect Link - Kawagishi, H. and T. Mizuno, 1988. Purification and properties of a beta-galactosyl-specific lectin from the fruiting bodies of Ischnoderma resinosus. FEBS Lett., 227: 99-102.
CrossRef - Khan, F., A. Ahmad and M.I. Khan, 2007. Chemical, thermal and pH-induced equilibrium unfolding studies of Fusarium solani lectin. IUBMB. Life, 59: 34-43.
CrossRefPubMedDirect Link - Khan, F., A. Ahmad and M.I. Khan, 2007. Interaction of Fusarium solani lectin with monosaccharides and oligosaccharides: A fluorometric study. Photochem. Photobiol., 83: 966-970.
CrossRefPubMedDirect Link - Khan, F., A. Ahmad and M.I. Khan, 2007. Purification and characterization of a lectin from endophytic fungus Fusarium solani having complex sugar specificity. Arch. Biochem. Biophys., 457: 243-251.
CrossRefPubMedDirect Link - Khan, F., A. Ahmad and M.I. Khan, 2010. Steady state and time resolved fluorescence quenching and chemical modification studies of a lectin from endophytic fungus Fusarium solani. J. Fluorescence, 20: 305-313.
CrossRefPubMedDirect Link - Kaur, A., S.S. Kamboj and J. Singh, 2006. Isolation of a new lectin from the bulbs of Crinum latifolium (L.). J. Biological Sci., 6: 9-14.
CrossRefDirect Link - Konska, G., 2006. Lectins of higher fungi (Macromycetes): Their occurrence, physiological role and biological activity. Int. J. Med. Mushrooms, 8: 19-30.
CrossRefDirect Link - Konska, G., J. Guillot, M. Dusser, M. Damez and B. Botton, 1994. Isolation and characterization of an N-acetyllactosamine-binding lectin from the mushroom Laetiporus sulphureus. J. Biol. Chem., 116: 519-523.
PubMedDirect Link - Kossowska, B., E. Lamer-Zarawska, M. Olczak and I. Katnik-Prastowska, 1999. Lectin from Beauveria bassiana mycelium recognizes Thomsen-Friedenreich antigen and related structures. Comp. Biochem. Physiol. B: Biochem. Mol. Biol., 123: 23-31.
PubMed - Leonidas, D.D., B.M. Swamy, A.G. Bhat, S.R. Inamdar and M.N. Kosmopoulou et al., 2003. Crystallization and preliminary X-ray crystallographic analysis of Sclerotium rolfsii lectin. Acta Crystallogr. D. Biol. Crystallogr., 59: 363-365.
CrossRefPubMedDirect Link - Lin, J.Y. and T.B. Chou, 1984. Isolation and characterization of a lectin from edible mushroom Volvariella Volvacea. J. Biochem., 96: 35-40.
PubMedDirect Link - Mancheno, J.M., H. Tateno, I.J. Goldstein, M. Martinez-Ripoll and J.A. Hermoso, 2005. Structural analysis of the Laetiporus sulphureus hemolytic pore-forming lectin in complex with sugars. J. Biol. Chem., 280: 17251-17259.
CrossRefPubMedDirect Link - Matsumura, K., K. Higashida, Y. Hata, J. Kominami, S. Nakamura-Tsuruta and J. Hirabayashi, 2009. Comparative analysis of oligosaccharide specificities of fucose-specific lectins from Aspergillus oryzae and Aleuria aurantia using frontal affinity chromatography. Anal. Biochem., 386: 217-221.
CrossRefPubMed - Matsumura, K., K. Higashida, H. Ishida, Y. Hata and K. Yamamoto et al., 2007. Carbohydrate binding specificity of a fucose-specific lectin from Aspergillus oryzae: A novel probe for core fucose. J. Biol. Chem., 282: 15700-15708.
CrossRefPubMedDirect Link - Nagata, Y., M. Yamashita, H. Honda, J. Akabane and K. Uehara et al., 2005. Characterization, occurrence and molecular cloning of a lectin from Grifola frondosa: Jacalin-related lectin of fungal origin. Biosci. Biotechnol. Biochem., 69: 2374-2380.
CrossRefPubMedDirect Link - Nicolson, G.L., 1974. The interactions of lectins with animal cell surfaces. Int. Rev. Cytol., 39: 89-190.
PubMed - Oda, Y., T. Senaha, Y. Matsuno, K. Nakajima and R. Naka et al., 2003. A new fungal lectin recognizing α(1-6)-linked fucose in the N-glycan. J. Biol. Chem., 278: 32439-32447.
CrossRefPubMedDirect Link - Olausson, J., L. Tibell, B.H. Jonsson and P. Pahlsson, 2008. Detection of a high affinity binding site in recombinant Aleuria aurantia lectin. Glycoconjugate J., 25: 753-762.
CrossRefPubMedDirect Link - Occena, I.V., E.R.E. Mojica and F.E. Merca, 2007. Isolation and partial characterization of a lectin from the seeds of Artocarpus camansi Blanco. Asian J. Plant Sci., 6: 757-764.
CrossRefDirect Link - Paaventhan, P., J.S. Joseph, S.V. Seow, S. Vaday and H. Robinson et al., 2003. A 1.7A structure of Fve, a member of the new fungal immunomodulatory protein family. J. Mol. Biol., 332: 461-470.
CrossRefPubMedDirect Link - Panchak, L.V. and V.O. Antoniuk, 2007. Purification of lectin from fruiting bodies of Lactarius rufus (Scop.: Fr.)Fr. and its carbohydrate specificity. Ukr. Biokhim. Zh., 79: 123-128.
PubMed - Pohleven, J., N. Obermajer, J. Sabotic, S. Anzlovar and K. Sepcic et al., 2009. Purification, characterization and cloning of a ricin B-like lectin from mushroom Clitocybe nebularis with antiproliferative activity against human leukemic T cells. Biochim. Biophys. Acta (BBA)-Gen. Subj., 1790: 173-181.
CrossRefPubMedDirect Link - Rini, J.M. and Y.D. Lobsanov, 1999. New animal lectin structures. Curr. Opin. Struct. Biol., 9: 578-584.
CrossRefPubMedDirect Link - Rosen, S., B. Ek, L. Raska and A. Tunlid, 1992. Purification and characterization of a surface lectin from the nematode- trapping fungus Arthrobotrys oligospora. J. Gen. Microbiol., 138: 2663-2672.
PubMedDirect Link - Rudiger, H. and H.J. Gabius, 2001. Plant lectins: Occurrence, biochemistry, functions and applications. Glycoconjugate J., 18: 589-613.
CrossRefPubMedDirect Link - Sathisha, G.J., Y.K. Prakash, V.B. Chachadi, N.N. Nagaraja and S.R. Inamdar et al., 2008. X-ray sequence ambiguities of Sclerotium rolfsii lectin resolved by mass spectrometry. Amino Acids, 35: 309-320.
CrossRefPubMedDirect Link - Sharon, N. and H. Lis, 1989. Lectins as recognition molecules. Science, 246: 227-234.
CrossRefPubMedDirect Link - Singh, R.S., R. Bhari and H.P. Kaur, 2010. Mushroom lectins: Current status and future perspectives. Crit Rev. Biotechnol.
CrossRefDirect Link - Singh, R.S., A.K. Tiwary and R. Bhari, 2008. Screening of Aspergillus species for occurrence of lectins and their characterization. J. Basic Microbiol., 48: 112-117.
CrossRefPubMedDirect Link - Slifkin, M. and R.J. Doyle, 1990. Lectins and their application to clinical microbiology. Clin. Microbiol. Rev., 3: 197-218.
PubMedDirect Link - Springer, T.A. and L.A. Lasky, 1991. Cell adhesion. Sticky sugars for selectins. Nature, 349: 196-197.
CrossRefPubMedDirect Link - Stepanova, L.V., V.E. Nikitina and A.S. Boiko, 2007. Isolation and characterization of lectin from the surface of Grifola frondosa (FR.) S.F. Gray mycelium. Mikrobiologiia, 76: 488-493.
PubMed - Suzuki, T., Y. Amano, M. Fujita, Y. Kobayashi and H. Dohra et al., 2009. Purification, characterization and cDNA cloning of a lectin from the mushroom Pleurocybella porrigens. Biosci. Biotechnol. Biochem., 73: 702-709.
CrossRefPubMedDirect Link - Sychrova, H., M. Ticha and J. Kocourek, 1985. Studies on lectins. LIX. Isolation and properties of lectins from fruiting bodies of Xerocomus chrysenteron and Lactarius lignyotus. Can. J. Biochem. Cell Biol., 63: 700-704.
PubMed - Tanaka, H., J. Toyama and R. Akashi, 2009. Molecular characterization of a galactose-binding lectin from Momordica charantia seeds and its expression in tobacco cells. Asian J. Plant Sci., 8: 544-550.
CrossRefDirect Link - Tateno, H. and I.J. Goldstein, 2003. Molecular cloning, expression and characterization of novel hemolytic lectins from the mushroom Laetiporus sulphureus, which show homology to bacterial toxins. J. Biol. Chem., 278: 40455-40463.
CrossRefPubMedDirect Link - Thakur, A., L. Pal, A. Ahmad and M.I. Khan, 2007. Complex carbohydrate specificity of lectin from fruiting body of Ganoderma lucidum. A surface plasmon resonance study. IUBMB. Life, 59: 758-764.
CrossRefPubMedDirect Link - Tronchin, G., K. Esnault, M. Sanchez, G. Larcher, A. Marot-Leblond and J.P. Bouchara, 2002. Purification and partial characterization of a 32-kilodalton sialic acid-specific lectin from Aspergillus fumigatus. Infect. Immunol., 70: 6891-6895.
CrossRefPubMedDirect Link - Tsivileva, O.M., V.E. Nikitina and E.A. Loshchinina, 2008. Isolation and characterization of Lentinus edodes (Berk.) singer extracellular lectins. Biochemistry, 73: 1154-1161.
PubMed - Vasta, G.R., H. Ahmed and E.W. Odom, 2004. Structural and functional diversity of lectin repertoires in invertebrates, protochordates and ectothermic vertebrates. Curr. Opin. Struct. Biol., 14: 617-630.
CrossRefPubMedDirect Link - Vetchinkina, E.P., O.I. Sokolov and V.E. Nikitina, 2008. Intracellular lectins of Lentinus edodes at various developmental stages of the fungus. Mikrobiologiia, 77: 496-501.
PubMed - Vranken, A.M., E.J.M. Van Damme, A.K. Allen and W.J. Peumans, 1987. Purification and properties of an N-acetylgalactosamine specific lectin from the plant pathogenic fungus Rhizoctonia solani. FEBS Lett., 216: 67-72.
CrossRef - Walti, M.A., P.J. Walser, S. Thore, A. Grunler, M. Bednar, M. Kunzler and M. Aebi, 2008. Structural basis for chitotetraose coordination by CGL3, a novel galectin-related protein from Coprinopsis cinerea. J. Mol. Biol., 379: 146-159.
CrossRefPubMedDirect Link - Wang, H. and T.B. Ng, 2005. First report of an arabinose-specific lectin. Biochem. Biophys. Res. Commun., 337: 621-625.
CrossRefPubMedDirect Link - Wang, H., T.B. Ng and V.E. Ooi, 1998. Lectins from mushroom. Mycol. Res., 102: 897-906.
CrossRefDirect Link - Wang, H., J. Gao and T.B. Ng, 2000. A new lectin with highly potent antihepatoma and antisarcoma activities from the oyster mushroom Pleurotus ostreatus. Biochem. Biophys. Res. Commun., 275: 810-816.
CrossRefPubMedDirect Link - Wang, H. and T.B. Ng, 2003. Isolation of a novel N-acetylglucosamine-specific lectin from fresh sclerotia of the edible mushroom Pleurotus tuber-regium. Protein Expression Purification, 29: 156-160.
CrossRefPubMedDirect Link - Wang, H., T.B. Ng and Q. Liu, 2003. A novel lectin from the wild mushroom Polyporus adusta. Biochem. Biophys. Res. Commun., 307: 535-539.
CrossRefPubMedDirect Link - Wang, H.X., T.B. Ng, W.K. Liu, V.E. Ooi and S.T. Chang, 1995. Isolation and characterization of two distinct lectins with antiproliferative activity form the cultured mycelium of the edible mushroom Trocholoma mongolicum. Int. J. Peptide Protein Res., 46: 508-513.
CrossRefPubMedDirect Link - Wasser, S.P. and A.L. Weis, 1999. Therapeutic effects of substances occurring in higher basidiomycetes mushrooms: A modern perspective. Crit. Rev. Immunol., 19: 65-96.
CrossRefPubMedDirect Link - Watanabe, T., I. Matsuo, J. Maruyama, K. Kitamoto and Y. Ito, 2007. Identification and characterization of an intracellular lectin, calnexin, from Aspergillus oryzae using N-glycan-conjugated beads. Biosci. Biotechnol. Biochem., 71: 2688-2696.
CrossRefPubMedDirect Link - Wimmerova, M., E. Mitchell, J.F. Sanchez, C. Gautier and A. Imberty, 2003. Crystal structure of fungal lectin: Six-bladed beta-propeller fold and novel fucose recognition mode for Aleuria aurantia lectin. J. Biol. Chem., 278: 27059-27067.
CrossRefPubMedDirect Link - Wong, J.H., H. Wang and T.B. Ng, 2009. A haemagglutinin from the medicinal fungus Cordyceps militaris. Biosci. Rep., 29: 321-327.
PubMedDirect Link - Wu, A.M., J.H. Wu, M.S. Tsai, G.V. Hegde, S.R. Inamdar, B.M. Swamy and A. Herp, 2001. Carbohydrate specificity of a lectin isolated from the fungus Sclerotium rolfsii. Life Sci., 69: 2039-2050.
CrossRefPubMedDirect Link - Yachnin, S., 1972. Inhibition of phytohemagglutinin-induced lymphocyte transformation by globulins; lack of correlation with phytohemagglutinin precipitation by serum proteins. J. Immunol., 108: 845-847.
PubMedDirect Link - Yachnin, S., 1975. Fetuin, an inhibitor of lymphocyte transformation. The interaction of fetuin with phytomitogens and a possible role for fetuin in fetal development. J. Exp. Med., 141: 242-256.
PubMedDirect Link - Yagi, F., M. Miyamoto, T. Abe, Y. Minami, K. Tadera and I.J. Goldstein, 1997. Purification and carbohydrate-binding specificity of Agrocybe cylindracea lectin. Glycoconjugate J., 14: 281-288.
CrossRefPubMedDirect Link - Yagi, F. and K. Tadera, 1988. Purification and characterization of lectin from Auricularia polytricha. Agric. Biol. Chem., 52: 2077-2079.
Direct Link - Yang, N., D.F. Li, L. Feng, Y. Xiang, W. Liu, H. Sun and D.C. Wang, 2009. Structural basis for the tumor cell apoptosis-inducing activity of an antitumor lectin from the edible mushroom Agrocybe aegerita. J. Mol. Biol., 387: 694-705.
CrossRefPubMedDirect Link - Yang, N., X. Tong, Y. Xiang, Y. Zhang, Y. Liang, H. Sun and D.C. Wang, 2005. Molecular character of the recombinant antitumor lectin from the edible mushroom Agrocybe aegerita. J. Biochem., 138: 145-150.
CrossRefPubMedDirect Link - Yang, N., X. Tong, Y. Xiang, Y. Zhang, H. Sun and D.C. Wang, 2005. Crystallization and preliminary crystallographic studies of the recombinant antitumour lectin from the edible mushroom Agrocybe aegerita. Biochim. Biophys. Acta (BBA)-Proteins Proteomics, 1751: 209-212.
CrossRefPubMedDirect Link - Zhang, G.Q., J. Sun, H.X. Wang and T.B. Ng, 2009. A novel lectin with antiproliferative activity from the medicinal mushroom Pholiota adiposa. Acta Biochim. Polonica, 56: 415-421.
PubMedDirect Link - Zhao, C., H. Sun, X. Tong and Y. Qi, 2003. An antitumour lectin from the edible mushroom Agrocybe aegerita. Biochem. J., 374: 321-327.
CrossRefPubMedDirect Link - Guillot, J. and G. Konska, 1997. Lectins in higher fungi. Biochem. Syst. Eco., 25: 203-230.
CrossRefDirect Link - Kobayashi, Y., K. Kobayashi, K. Umehara, H. Dohra, T. Murata, U. Usui and H. Kawagishi, 2004. Purification, characterization and sugar binding specificity of an N-glycolylneuraminic acid-specific lectin from the mushroom Chlorophyllum molybdites. J. Biol. Chem., 279: 53048-53055.
CrossRefPubMedDirect Link