
T.O. Adeyemi
Molecular Systematics Laboratory, Department of Botany, University of Lagos, Akoka, Lagos, Nigeria
O.T. Ogundipe
Molecular Systematics Laboratory, Department of Botany, University of Lagos, Akoka, Lagos, Nigeria
J.D. Olowokudejo
Molecular Systematics Laboratory, Department of Botany, University of Lagos, Akoka, Lagos, Nigeria
International Journal of Botany
Year: 2012 | Volume: 8 | Issue: 1 | Page No.: 45-49







ABSTRACT
Members of the family Sapindaceae provides various benefits in combating increasing malnutrition and poverty in West Africa. Due to their multiple uses, their high nutritional and medicinal value, members of Sapindaceae have been identified as one of the most important forest species to be conserved and valued in Africa. Therefore, it is important to study the potential future distribution of this species and determine strategies for conservation. In order to achieve these, suitability of sites in Africa was evaluated for potential conservation using spatial environmental data in MAXENT modelling framework. A total of 153 geo-referenced records of Sapindaceae were assembled from herbarium records and fieldwork and climatic data were acquired from the Worldclim Database. The main variables that contributed towards predicting the species distribution were annual precipitation and temperature seasonality. Results suggest that the distribution model was excellent with training AUC value of 0.974 and test AUC value of 0.960 confirming the wide distribution of Sapindaceae in West Africa. Also, the environmental variables that affected the model the most are minimum temperature of the coldest month (35.4%), precipitation of the coldest quarter (16.9%) and precipitation of wettest month (14.9%). Recommendations for different conservation strategies include in situ conservation in Protected Areas; ex situ conservation in seed banks and conservation through ‘sustainable utilization’.
PDF Abstract XML References Citation
Received: January 03, 2012;
Accepted: January 30, 2012;
Published: April 25, 2012
How to cite this article
T.O. Adeyemi, O.T. Ogundipe and J.D. Olowokudejo, 2012. Species Distribution Modelling of Family Sapindaceae in West Africa. International Journal of Botany, 8: 45-49.
DOI: 10.3923/ijb.2012.45.49
URL: https://scialert.net/abstract/?doi=ijb.2012.45.49
DOI: 10.3923/ijb.2012.45.49
URL: https://scialert.net/abstract/?doi=ijb.2012.45.49
INTRODUCTION
Climate change is an increasing concern worldwide; issues such as crop failure and food security are progressively becoming more worrisome. The fruit pulp, leaves and seeds of the several genera in the family Sapindaceae are a good source of vitamins and minerals (Metcalfe and Chalk, 1957; Chapman, 1984) and are common ingredients in traditional herbal remedies in rural and urban Africa (Odugbemi and Akinsulire, 2006). Apart from food, they also serve as shade to local people (Watson and Dallwitz, 2008). Owing to the combination of health claims (e.g. anti-inflammatory (Sofidiya et al., 2007), anti-fungal (Adesegun et al., 2008), anti-diarrhoeal, (Antwi et al., 2009), aphrodisiac (Muanya and Odukoya, 2008), pre-biotic and antioxidant functions (Ripa et al., 2010), for eye infections (Pendota et al., 2008), high calcium content) and food technological functions (high pectin and fiber content), fruit pulp of several genera has been identified as a suitable candidate for a new generation of functional foods and drinks (Chapman, 1984). Some yield valuable most widely used is Tulipwood (Harpullia pendula). Many other members of the Sapindaceae are important nuts, or sources of beverages, or oils. Saponins are present in the fruits, seeds, barks, twigs, leaves, pericarp and other tissues of several species; the bark of Jagera pseudorhus Blume contains large amounts and is used as a foaming agent and as a fish poison. Some types such as Sapindus saponaria are used as soap substitutes in the tropics. The fruit of Dodonaea viscosa (hop bush) has been used as a substitute for hops in the brewing of beer.
Also, due to formation of residential, industrial and other layouts, the natural slopes, vegetation and land cover gets disturbed however, species composition can be predicted by local environmental conditions (Ali and Kauser, 2006). Although members of Sapindaceae are not commonly cultivated, they can be successfully propagated. Investigating potential sites for cultivation is therefore an important first step towards the conservation of the representative species. In this study, geo-referenced Sapindaceae locality data and spatial environmental data were used in MAXENT, the species distribution modelling framework to predict potential sites for cultivation in Western Africa.
MATERIALS AND METHODS
Species data: A total of 153 Sapindaceae growing localities (without duplicates) were assembled from diverse sources. About 32% of the localities came from recent fieldwork while 68% were herbarium records. In order to reduce the effect of temporal bias and to match the species data with the environmental climatic data (produced over the period 1950-2000), herbarium records collected before 1950 were eliminated.
Environmental data: Growing potential was assessed by characterising the climate at the presence locations. Twenty spatial datasets from the WorldClim database (at 5' resolution) (Hijmans et al., 2005) were used. The Worldclim dataset included altitude and 19 bioclimatic variables derived from temperature and rainfall. Although 21 spatial datasets were used in the beginning, some were eliminated during modelling due to their low contribution to the final model. In agricultural systems, abiotic factors are more limiting than biotic factors (such as competition or dispersal); therefore no biotic factors were included in this study. Also, it was assumed that at a continental scale, climatic variables were much more limiting than human population.
Species distribution modelling: For each plant sample, GPS coordinates of the collection sites were recorded and subjected to modelling using the Maximum Entropy Programme (MAXENT) (Phillips and Dudik, 2008). Maxent predicts the potential species distribution by estimating the probability distribution of maximum entropy across a specified region, subject to a set of constraints that represent the missing information (lack of absence data) about the target distribution (Phillips et al., 2006). The Maxent method is currently considered to be the most accurate approach to modelling presence-only data (Elith et al., 2006; Pearson et al., 2007).
Model validation: Model performance was evaluated using several methods. First, model performance was determined by assigning a subset of the presence records for training and using the remaining records to test the resulting model. A good model should predict correctly the presence of Sapindaceae in the test locations. As performance can vary depending upon the particular set of data selected for building the model and for testing it, 10 random partitions of the presence records were made to assess the average behaviour of Maxent, following Phillips et al. (2006). Each partition was created by randomly selecting 75% of the total presence records to build the model and the remaining 25% of presence records were used for testing. However, the full set of presence records were used to build the final model to obtain the best estimate of the species distribution (for all records model, West Africa model). Second, Receiver Operating Characteristic (ROC) analysis was used to evaluate how well the Maxent model compared to a random prediction. The area under the ROC curve (AUC) serves as a measure of model performance in terms of sensitivity versus specificity. The sensitivity for a particular threshold is the fraction of all positive instances that are classified as present and specificity is the fraction of all negative instances that are classified as not present. The value of the AUC is typically between 0.5 (random) and 1.0. The closer the AUC value to 1, the better the model performance. Moreover, the success of the model was also evaluated by visually examining how well the mapped probability values matched the presence records. A good model should produce regions of high probability that cover the majority of presence records and areas of low probability should contain few or no presence points.
RESULTS
Potential distribution of Sapindaceae in West Africa: All records model, the resulting Maxent cultivation potential map for Africa predicted members of Sapindaceae to occur in most of West Africa. For the West Africa model, the cultivation potential map predicted Sapindaceae to grow in most of the Sahel from Senegal to south-west Cameroon and in southern Ghana-Nigeria (Fig. 1).
Model performance: All the ten generated training/testing models showed a high level of performance when compared to random (where the AUC would be 0.5). Training gained in this model ranged from 2.139 - 2.668 (Fig. 2) while the AUC value was 0.974 (Fig. 3). Test AUC values was lower but close to training AUC i.e., 0.960 (Fig. 3) while the test gained was 2.117 (Fig. 4). The training/testing models correctly predicted most of the test locations in all models and standard deviation is 0.007.
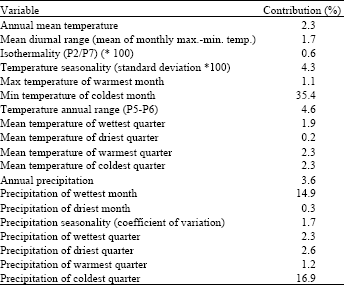
Variable contribution: The model indicated that the presence of members of Sapindaceae was mainly associated with minimum temperature of the coldest month (35.4%), precipitation of the coldest quarter (16.9%) and precipitation of wettest month (14.9%) (Table 1).
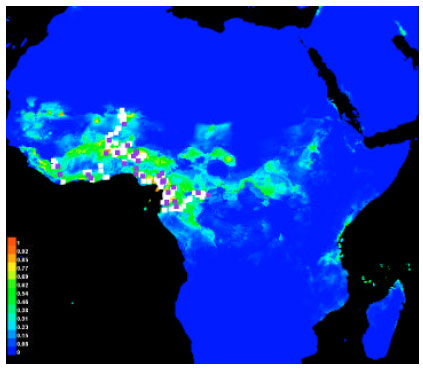 | |
| Fig. 1: | Species distribution: Training vs. test locations, Green: The typical condition of where species are found, Blue: Low probability of suitable condition, Red: High probability of suitable condition, White dots: The presence locations used for training, while violet dots show test locations |
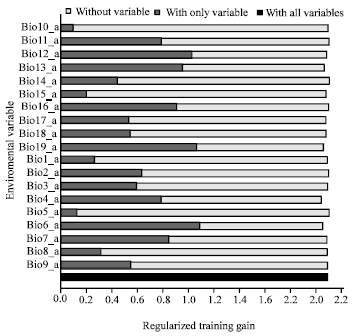 | |
| Fig. 2: | Jack-knife of regularized training gain for Sapindaceae |
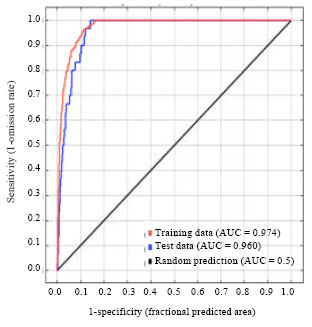 | |
| Fig. 3: | Sensitivity vs. specificity test on Sapindaceae |
Maxent’s jackknife test of variable importance also suggested that the variables which contributed the most to the model were good predictors as they had the most information not contained in other variables and they could describe the distribution of Sapindaceae on their own (without the other variables).
| Table 1: | Variable contributions to the MAXENT model |
 | |
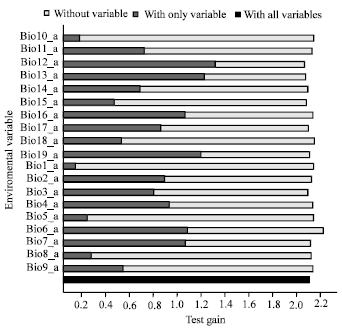 | |
| Fig. 4: | Jack-knife of regularized test gain for Sapindaceae |
DISCUSSION
Although members of the family Sapindaceae have been recorded to be widely distributed in Africa, their occurrence is being threatened by high rate of deforestation and agricultural practices leading to loss of forest and threatened status of the family as recorded in the IUCN R.L. (2008). MAXENT, a machine-learning method based on the principle of maximum entropy, are used to predict distribution for each species of butterfly under current and future climates (Sofy et al., 2012). Data from this model is used to test the efficiency of the modeling program through the evaluation of the area under the Receiving Operator Curve (AUC) (Elith et al., 2006; Phillips et al., 2006).
Species distribution modelling with climatological data analysis using nineteen (19) standard environmental variables from the WorldClim database revealed that the species distribution is most affected by the minimum temperature of the coldest month (bio 6) with a variable contribution of 35.4%. This is followed by precipitation of coldest quarter (bio 19) and precipitation of the wettest month (bio 13) with 16.9 and 14.9% contribution, respectively. These results support the location of Sapindaceae mostly in the tropics rather than the temperate region.
Also, from the AUC curve generated, it can be concluded that this present model is more reliable as it predicts the species distribution better than a typical random model as shown in Fig. 3. This makes it useful for future projections. Recommendations for different conservation strategies include in situ conservation in Protected Areas; ex situ conservation in seed banks; and conservation through ‘sustainable utilization’.
ACKNOWLEDGMENTS
Special thanks to the Head of Molecular Systematics Laboratory, Department of Botany, University of Lagos for providing enabling environment for this study. Also, we are grateful to the Explorer’s Club USA for providing the exploration funds.
REFERENCES
- Adesegun, S.A., H.A.B. Coker and M.T. Hamann, 2008. Antifungal triterpenoid saponins from Lecaniodiscus cupanioides. Res. J. Phytochem., 2: 93-99.
CrossRefDirect Link - Ali, S.M. and S. Kauser, 2006. Plant communities analysis of selected urban flora of Islamabad. J. Applied Sci., 6: 177-182.
CrossRefDirect Link - Antwi, S., O.N.K. Martey, K. Donkor and L.K.N.A. Okine, 2009. Anti-diarrhoeal activity of Blighia sapida (Sapindaceae) in rats and mice. J. Pharmacol. Toxicol., 4: 117-125.
CrossRefDirect Link - Elith, J., C.H. Graham, R.P. Anderson, M. Dudik and S. Ferrier et al., 2006. Novel methods improve prediction of species' distributions from occurrence data. Ecography, 29: 129-151.
CrossRefDirect Link - Hijmans, R.J., S.E. Cameron, J.L. Parra, P.G. Jones and A. Jarvis, 2005. Very high resolution interpolated climate surfaces for global land areas. Int. J. Climatol., 25: 1965-1978.
CrossRefDirect Link - Muanya, C.A. and O.A. Odukoya, 2008. Lipid peroxidation as index of activity in aphrodisiac herbs. J. Plant Sci., 3: 92-98.
CrossRefDirect Link - Pearson, R.G., C.J. Raxworthy, M. Nakamura and A.T. Peterson, 2007. Predicting species distributions from small numbers of occurrence records: A test case using cryptic geckos in Madagascar. J. Biogeogr., 34: 102-117.
CrossRef - Pendota, S.C., D.S. Grierson and A.J. Afolayan, 2008. An ethnobotanical study of plants used for the treatment of eye infections in the Eastern Cape province, South Africa. Pak. J. Biol. Sci., 11: 2051-2053.
CrossRefPubMedDirect Link - Phillips, S.J. and M. Dudik, 2008. Modelling of species distributions with Maxent: New extensions and a comprehensive evaluation. Ecography, 31: 161-175.
Direct Link - Phillips, S.J., R.P. Anderson and R.E. Schapire, 2006. Maximum entropy modeling of species geographic distributions. Ecol. Modell., 190: 231-259.
CrossRefDirect Link - Ripa, F.A., M. Haque and I.J. Bulbul, 2010. In vitro antibacterial, cytotoxic and antioxidant activities of plant Nephelium longan. Pak. J. Biol. Sci., 13: 22-27.
CrossRefDirect Link - Sofy, A.R., A.A. Mousa, A.M. Soliman and K.A. El-Dougdoug, 2012. The Limiting of climatic factors and predicting of suitable habitat for citrus gummy bark disease occurrence using GIS. Int. J. Virol., 8: 165-177.
CrossRefDirect Link - Watson, L. and M.J. Dallwitz, 2008. The grass genera of the world: Descriptions, illustrations, identification and information retrieval; including synonyms, morphology, anatomy, physiology, phytochemistry, cytology, classification, pathogens, world and local distribution and references. The Grass Genera of the World.
- Sofidiya, M.O., O.A. Odukoya, A.J. Afolayan and O.B. Familoni, 2007. Survey of anti-inflammatory plants sold on herb markets in Lagos Nigeria. Int. J. Bot., 3: 302-306.
CrossRefDirect Link