
T. A. El-Shahawy
Department of Botany, National Research Center, Dokki, Cairo, Egypt
L. K. Balbaa
Department of Botany, National Research Center, Dokki, Cairo, Egypt
K. G. El-Rokiek
Department of Botany, National Research Center, Dokki, Cairo, Egypt
International Journal of Agricultural Research
Year: 2007 | Volume: 2 | Issue: 5 | Page No.: 411-425







ABSTRACT
A pot trial was conducted to study the potential effect of Glutathione (GSH) and Ascorbic acid (ASA) either applied alone or in combination at different rates of concentration (50-250 and 25-100 mg L-1, respectively) on controlling weeds and yield productivity of marjoram (Origanum majorana L.). The two compounds and their combinations were applied as a post-emergence application after 30 days from sowing. Applying the two compounds alone at different concentrations significantly inhibited the broad and narrow-leaved weeds by up to 56 and 60%, respectively. The combined effect of the two compounds was more efficient in this regard (up to 100% weed reduction) than the individual application. Marjoram growth and development was seriously affected (up to 63% growth induction) at all levels of studied criteria. The effect was more pronounced on increasing the elementary components of total sugar, total nitrogen and oil yield (up to 91%); irrespective of the rate and sort of application. The in vitro study of the allelopathic influence of marjoram oil extracts against seed germination (%) and seedling root and shoot length of certain broad and narrow-leaved weeds (e.g., Amaranthus cruentus and Echinochloa crus-galli) revealed of more superiority of the treated plants rather than the untreated one. GLC analysis also showed a significant increasing in the terpene (oxygen-containing monoterpenes in particular) and phenolic substances in the treated plants. It has been suggested that applying GSH or ASA either alone or in combination might have a direct and/or indirect impact on increasing the allelopathic capacity of marjoram plants by increasing their content of the terpene and phenolic substances, which could be the key factor of the allelopathic influence of marjoram plants in their vicinity.
PDF Abstract XML References
How to cite this article
T. A. El-Shahawy, L. K. Balbaa and K. G. El-Rokiek, 2007. The Ability of Glutathione and Ascorbic Acid to Increasing Growth Vigor and Enhancing the Allelopathic Capability of Marjoram (Origanum majorana L.) Against Growth Retardatory Weeds. International Journal of Agricultural Research, 2: 411-425.
DOI: 10.3923/ijar.2007.411.425
URL: https://scialert.net/abstract/?doi=ijar.2007.411.425
DOI: 10.3923/ijar.2007.411.425
URL: https://scialert.net/abstract/?doi=ijar.2007.411.425
INTRODUCTION
With increasing emphasis on more effective and economically sounder medicines, the world nowadays has given special attention to the medicinal and aromatic plants as an excellent source for secondary metabolites that can be used directly or after certain modifications in controlling a wide range of human diseases. There is a considerable amount of research of plant-containing bioactive agents, many of which have established pharmacological activity and some of others have proved useful as natural pesticides (Jacobson, 1982; Green et al., 1987; Duke et al., 2002). Marjoram (Origanum majorana L.) is one of such ambitious plants that currently received considerable attention for its aromatic and pharmacological properties.
Origanum species have been used in medicine and as a spicy since antiquity, mainly because of their highly content of essential oils, which consist of considerable amounts of terpene derivatives i.e., Monoterpene Hydrocarbons (MTHC) and Oxygen-Containing Monoterpenes (OCMT) as well as carvacrol and thymol phenolic derivatives (Sarer et al., 1982). The crude drug is an oriental folk medicine for intestinal function controlling, stomachaches, indigestion, toothache, asthma, rheumatism, headache and as an antispasmodic agent of various agonists and Ca2+ (Simon et al., 1984). However, the importance of marjoram and its components has recently been emerged from containing highly biocidal activity toward a wide range of mono and dicotyledonous weeds, that phenomenon called allelopathy. Hydroquinone is amongst the most powerful allelopathic agents identified in marjoram that provide efficiency under lab and field conditions for controlling weeds in many crops species (Assof et al., 1988). The compound was strongly qualified to be developed as natural herbicide, but its cytotoxic effect has precluded between achieving this task. Duke (1991) reported that of the hundreds of known plant-derived monoterpenes, camphor, 1,8-cineole and pulegone were the most suppressive compounds.
Glutathione (GSH), a thiol containing-tripeptide, is widely distributed in all plants and animal cells including human bodies. The compound plays a pivotal role in protecting plants from the damage effect of the oxidative stress caused by extremes of temperature, draught, herbicides, or air pollutants (Noctor and Foyer, 1998). In concert with Ascorbic Acid (ASA), GSH acts by scavenging free radicals and hydrogen peroxide in a mechanism by which intercellular biological structures and hence its activity are protected against potential damaging effects of peroxide free-radical mediated attacks (Hausladen and Alscher, 1993). Hausladen and Alscher (1993) described that the major function of GSH in the protection of cells against the toxic effect of the free radicals is to keep the free radical scavenger ASA in its reduced form and hence active form. Conceivably, the capacity of GSH to participate in the redox regulation in plant cells is, to a great extent, depends on its absolute concentration and the ratio of GSH/GSSG, the latter is the oxidized form of GSH (May et al., 1998; Foyer et al., 2001).
Such system of GSH/ASA are thought to be has a direct correlation on preserving seed's viability of plants, e.g., silver maple (Acer saccharinum L.), when exposed to hard conditions of high temperature as in desiccation process for food industry. Pukacka and Ratajczak (2005) have a confirming view of that since they noticed remarkable changes in GSH/ASA profiles and associated enzymes in embryo axis and cotyledons of those seeds subjected to high temperatures whereof invite them to declared that such changes in GSH/ASA are inevitable to counteracting the production of reactive oxygen species e.g., O2 and H2O2 during desiccation. The ability of maintaining redox homeostasis in cell is the reason behind this authority (May et al., 1998). Rather than their significant role as antioxidants, the GSH and ASA have more functional positions on regulating and controlling the physiological and biochemical processes within living cells. In this context, Lappartient and Touraine (1996) reported that GSH regulates sulphar uptake at root level, which in tern is used by GSH-s transferase in the detoxification of exobiotics and in controlling cellular heavy metal concentration (Marrs, 1996). Sanchez-Fernandez et al. (1997) assumed that GSH takes indispensable part in redox control of cell deviation. On the other hand, of the many functions of ASA have implicated, photoprotection and regulation of photosynthesis are the most prominent functions ascribed to the compound (Forti and Elli, 1995). Additionally, Patil and Lall (1973) confirmed that the ASA is involved in managing certain enzymatic functions and have a place in biosynthesis of nucleic acids and protein. ASA plays an important role in preserving the activity of the enzymes that contain prosthetic transition ions (Padh, 1990). Further, ASA can also work on maintaining α-tocopherol in the reduced form and therefore links GSH to the dominant free radical scavenger in membranes (Hess, 1994).
Allelopathy is the chemical interaction of one plant upon another including inhibitory and stimulatory effects (Rice, 1984). Allelopathy is quit useful to be used in different agricultural practices. Of its important are weed control, intercroppings and nutrient recycling (Rizvi and Rizvi, 1987). Allelopathy has also opened the door for new group of natural herbicides to come to light. Isolation and identification of the allelopathic responsible components have taken a large stride in this regard. The herbicidal selectivity, acting on new sites of action, low environmental stability and safety to whole forms of life all are of its great attributes that raising men's attention to use them as an alternative source of synthetic herbicides. Improving the allelopathic ability within crop populations has attracted the others by using several strategies and techniques such as plant breeding, genetic manipulation and/or shooting with exotic compounds (Olofsdotter et al., 2002; Yongqing, 2005).
Of the most important heading, exotic compounds and their potential to enhance allelopathic activity of the crops, many researchers have stated that synthetic compounds can be used successfully to elicit secondary metabolites that confer resistance to plant pests. For instance, application of low levels of diphenyl ether herbicides cause huge increases in tissue production of phytoalexins (Kogan and Paxton, 1983). Similarly, certain none phytotoxic compounds such as chitosan and ALA are very phytotoxic inducers (Pearce and Ride, 1982; Rebeiz et al., 1988a, b). Recently, Armero et al. (2001) found significant accumulation of phytoalexins and constitutive isoflavonoids upon GSH treatment which are of great interest in deterring pathogen attacks (Graham and Graham, 1991). Similar response was obtained with using salicylic and jasmonic acids (Kauss et al., 1994; Kauss and Jeblick, 1996). The phenolic compounds is not only viable in defense mechanisms against insect or pathogen pests, but also have from the phytotoxic properties that was invited researchers to considered them in herbicides discovery strategies (Sene et al., 2000; Wu et al., 2001). Numerous reports have established on the herbicidal efficiency of the phenolic derivatives and their ability to be developed as natural herbicides (Shettel and Balke, 1983). For example, the halogenated benzoic acid herbicides e.g., dicamba, cholramben and picloram are in fact of matter analogues of benzoate which is a phenolic plant product (Duke, 1992).
Marjoram has a great future in Egypt. It is widely used between public as a folk medicine. The crop has recently been given special attention between pharmacological and food companies to be used as a basic source for producing drugs or as spices. Thus, the aim of the present work is to study the influence of glutathione and ascorbic acid on the development of marjoram plants and their biocidal activity against wide range of weeds associated with such plants.
MATERIALS AND METHODS
Two greenhouse experiments were conducted during two successive seasons in the period of (2003-2005) at the screening house of National Research Center, Egypt. Marjoram (Origanum majorana L.) seeds were obtained from Medicinal and Aromatic Plant Section, Agriculture Research Center, Ministry of Agriculture, Egypt. The seeds were sown in a bed of 24% sand, 47% silt and 29% clay on 10-15th October in the two successive seasons. After three months of sowing, the seedlings (10 cm height) were transplanted into the main site of the experiment which consists of 102 pots (30x40 cm/each) filled with silty clay soil; the pot of each included 5 seedlings. The pots were infested simultaneously with certain broad and narrow leaf weeds such as pigweed (Amaranthus cruentus L.), beet (Beta vuligaris L.), stinkgrass (Eragrostis cilianensis L.) and bluegrass (Poa annua L.). The plants were watered and fertilized according to the recommendations. A serial concentration of glutathione (50, 100, 250 mg L-1) and ascorbic acid (25, 50 and 100 mg L-1) were prepared by dissolving in 5% aqueous acetone solution. The two compounds were applied as a post-emergence, either alone or in combination (under the same concentrations) after 40 days from transplanting; distilled water and hand weeding were used as controls. Six replicates were used for each treatment in a completely randomized design.
The plant samples and other observations on growth and development of marjoram and associated weeds were estimated as follows:-
Marjoram Development
Two cuts were taken during marjoram growth. The first and second cuts were taken after 3 and 6 months of foliar application, respectively (e.g., bloom stage). The plant samples were used to determine several vegetative characteristics including plant height (cm), number of branches/plant and fresh and dry weights of shoot biomass (g).
Determination of the total carbohydrates was conducted in the dried herb of treated and untreated plants according to Smith et al. (1964) and estimated calorimetrically by phenolic-sulphuric acid technique (Dubois et al., 1956). The total nitrogen content was estimated using the Micro-Kjeldahl methods (Allen, 1953) and Markham apparatus (Markham, 1942). The total oil yield (mg/plant) and its percent were determined immediately in the fresh herb by using steam hydro-distillation technique established by Egyptian Pharmacopoeia (1984). Half of the extracted oil of the first cutting was used for the qualitative and quantitative analysis of the oil components including monoterpenes and phenols. The remainder oil extract was re-dissolved in acetone (10%) solution and examined in vitro for their biological activity at different rates of concentration (0.5, 1.0 and 2.5 mg mL-1).
The qualitative and quantitative analysis of the putative compounds of the essential oil including Monoterpene Hydrocarbons (MTHC), Oxygen-Containing Monoterpenes (OCMT) and phenolic substances were carried out using Hewlet Packard GC apparatus, Agilent Technologies (mode 6890 hp), connected to PC furnished with Chrom-Card Software. The analysis was performed by inject 1 μL of oil samples and authentic compounds at 250°C (inlet oven temperature) onto capillary Zebron glass column (Hp-5; 30x0.32 mm) with film thickness (0.25 μm) consist of 5% pheny-95% dimethy-polysiloxane. The chromatograph was operated in a constant flow rate of nitrogen (30 mL min-1) with an oven temperature programme: 2 min isothermal at 70°C, then at 4°C min-1 up to 190°C and finally held isothermally for 13 min. The oil components were detected on a flam ionization detector at 280°C. For increasing measurement sensitivity, an attention time degree of 4 was selected and chart speed was adjusted at 0.50 cm min-1. Chromatograms were recorded and the peak areas and retention times were used in identifying and determining the chemical constitutes of marjoram oil in corresponding to the pure standard. The relative amount of each compound was expressed as a percentage from the total mixture.
Weeds Development
Weed samples were taken after three weeks of foliar application within the two successive seasons by hand pulling of all grown weeds of each treatment (3 pots only of each). The weeds were classified into broad and narrow leaf weeds and each group was subjected to fresh and dry weight (g/pot) determination.
The biological activity of marjoram crude oil extracts was examined in vitro at different concentrations on the seed germination and seedling growth of pigweed (Amaranthus cruentus L.) and barnyard-grass (Echinochloa crus-galli L.) weeds, by placing 20 seeds/each into 7 cm diam. Petri dishes containing filter paper (Whatman No.1). The aqueous solutions were added at rate of (2.5 mL/dish) at the beginning of the screening, followed by 1 mL 5 days later. Three replicates were used for each treatment. An extra three dishes were used as control. The dishes were arranged in a completely randomized design. The seed germination (%), root and shoot elongation (cm) of the treated and untreated plants were estimated within 15 days of the experiment. The data were subjected to standard analysis of variance at 5% probability according to Snedecor and Cochran (1967). The biological activity of marjoram extract was estimated using equations of Itokawa et al. (1982).
RESULTS
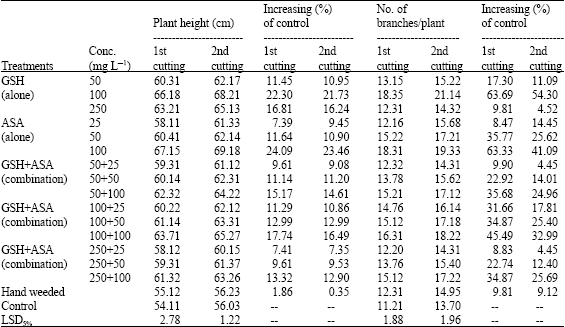
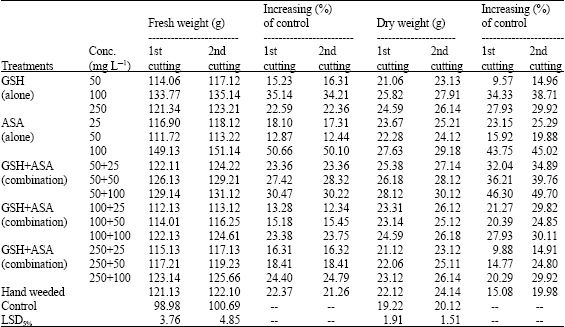
It was obvious that applying the two compounds either alone or in combination significantly increased the mass vegetative growth of marjoram plants including fresh and dry weights, as well as plant height and number of branches/plant (Table 1). The number of branches per plant is the criteria that recorded the highest response (up to 63.33% growth increasing) of all treatments in comparison with plant height (up to 24.09%) and fresh and dry weights (up to 50.66 and 49.70%, respectively). Of applying the two compounds alone, it was noted in the time that ASA provided good results in enhancing marjoram growth at the different concentrations (e.g., ascending results, increased as the concentration increased), applying GSH was better only at the lowest (50 mg L-1) and moderate (100 mg L-1) concentrations.
| Table 1: | Effect of GSH, ASA and their combinations on marjoram growth. (Combined analysis for the two successive seasons) |
 | |
| Table 1: | Continued |
 | |
At the highest concentration of the compound (250 mg L-1), the efficiency was retreated to register the closest value to the control.
Application of the two compounds together in tank mixture was fractionally more efficient than the case described above. Paying no attention to the season of the study, sort and rate of application, the data revealed of up to 49.70% efficient in increasing fresh and dry weights of shoot biomass and up to 17.74 and 45.49% in plant height and number of branches per plants, respectively. Hand weeding caused number of branches/plant increased by up to 9.81% and fresh and dry weights by 9.88-22.37%, but insignificantly affected the plant height as compared with control or other treatments.
| Table 2: | Effect of GSH, ASA and their combinations on growth and development of associated weeds. (Combined analysis for the two successive seasons) |
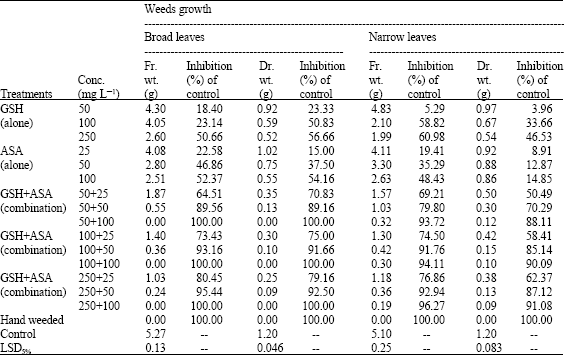 | |
On the other hand spraying GSH and ASA substantially suppressed occurring weeds flora including broad (pigweed, A. cruentus L.; beet, B. vuligaris L.) and narrow-(stinkgrass, E. cilianensis; bluegrass, P. annua L.) leaved weeds, either they applied alone or in combination; irrespective of the rate of application (Table 2). Applying GSH and ASA alone significantly inhibited the fresh and dry weights of shoot biomass of broad-leaved weeds by up to 56.66% and grassy one by up to 60.98%. However, the combined application treatments were so far more effective against both types of weeds, reducing their growth respect to the control by up to 100%. Hand weeding is of great interest. It is completely eradicated both types of broad and narrow leaf weeds, recording of that the best results overall the others.
In general, the efficiency of GSH, ASA or their combinations in suppressing weeds growth was increased as the concentration increased. This was more pronounced with the combined application rather than the singlet one. In this context, the treatments containing the highest level of GSH (250 mg L-1) recorded the best results amongst the others for controlling weeds infested marjoram, which in most cases entirely killed weeds within 20 days of foliar application. Of those containing the lowest and moderate concentrations of GSH, the response was feeble to a certain (50.49-100% weed reduction), but still remains more efficient than the individual application of either of the two components as compared with control.
The data in Table 3 seem to be clear that applying GSH, ASA or their combinations significantly increased the allelopathic ability of marjoram plants at all levels of application. In vitro evidences revealed that the plants subjected to GSH/ASA treatments have given the most powerful essential oils toward suppressing seed germination (%) and seedling growth of the two assayed weeds as compared with controls. With no exception, such inhibitory effect was increased as the concentration increased. Some degree of fluctuation was observed between the different treatments of oil extracts. The oil extracts obtained after foliar application by GSH alone at different rates was found to be more efficient in inhibiting the seed germination percent (9.74-100%) and root and shoot elongation (11.55-100%) of either broad or narrow-leaved weeds than those obtained after foliar application by ASA. Applying oil extracts obtained after foliar application by GSH/ASA-mixed components was far more effective in comparison with the controls.
| Table 3: | In vitro herbicidal effects of oil extracts obtained after greenhouse treatment with GSH and ASA against seed germination characteristics of certain broad and narrow-leaved weeds |
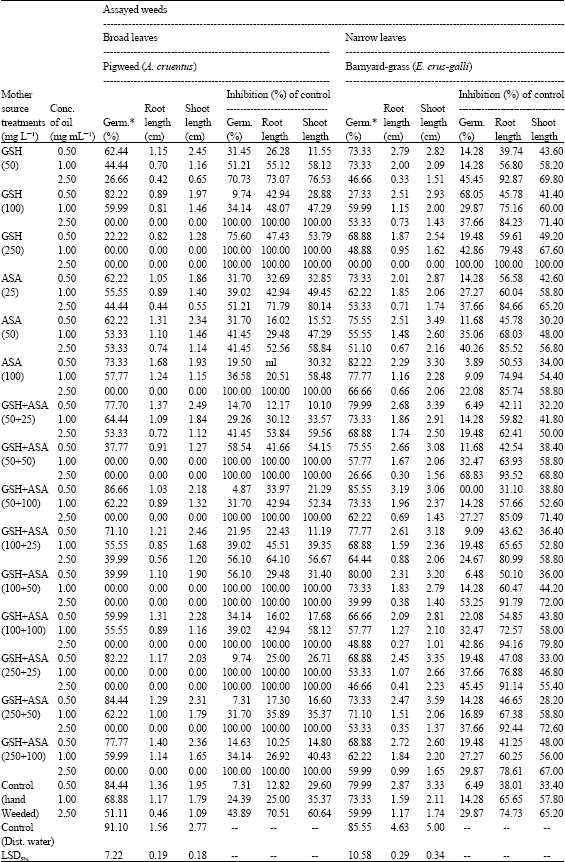 | |
| *Germ. = Germination | |
It caused complete growth failure in many cases of in vitro investigation. However, it seems to be clear that the herbicidal influence over both types of weeds was a species-specific to a great extent, since the data revealed of more sensitivity toward the broad leaf weed (pigweed) than the grassy (barnyard-grass) one.
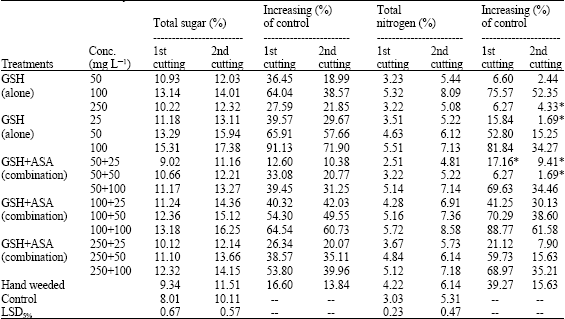
The chemical constituents of marjoram plants and their influence by GSH and ASA are shown in Table 4 and 5. This composes the effect on the elementary components of total sugar (%), nitrogen (%), total oil (%) and oil yield (mg/plant), as well as the chemical constituents of the essential oil including terpenoids (MTHC; OCMT) and phenols. Superficially, the results have taken similar trend as much the influence on marjoram growth. On total sugar, nitrogen and oil contents, applying ASA alone showed good results (2.85-91.13% increasing over control) at all of its concentrations (Table 4).
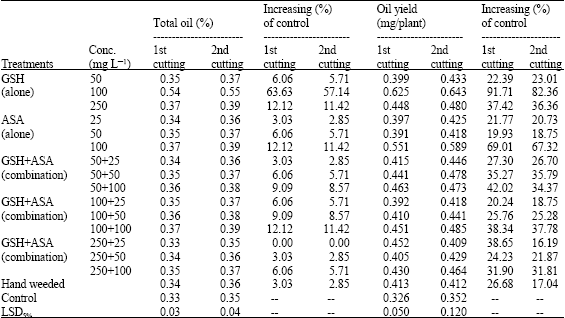
| Table 4: | Potential effect on the elementary components of total sugar, total nitrogen and oil yield of marjoram plants. (Combined analysis for the two successive seasons) |
 | |
| Table 4: | Continued |
 | |
| * Inhibition | |
effect in this regard fluently increased as the concentration increased. Again, GSH has showed superiority at the lowest and moderate concentrations only, unlike the highest concentration which mostly was ineffective. Applying the two compounds in combination was fluctuated in their efficiency in increasing such components of total sugar, nitrogen or oil yield. The best results (11.42-88.77% increasing over control) were obtained with applying GSH plus ASA at 100 mg L-1 for both of them. On the internal scene of the susceptibility, no much significant difference were observed between the different studied criteria in their influencing by GSH/ASA treatments.
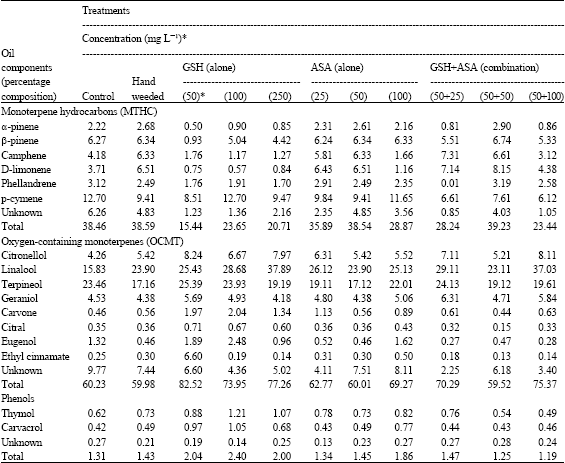
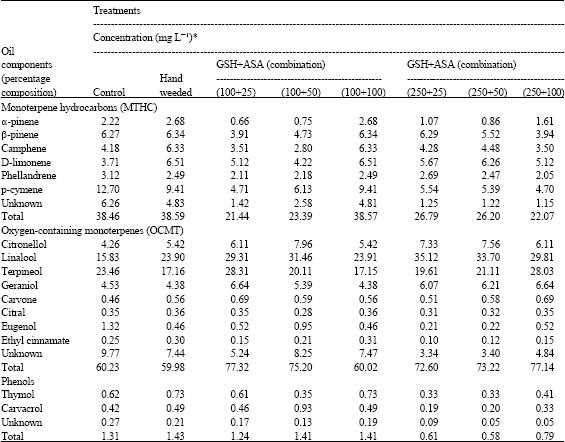
The chemical constituents of marjoram oil including MTHC (e.g., camphene, D-limonene, phellandrene, p-cymene, α-and β-pinene) and OCMT (e.g., citronellol, linalool, terpineol, geraniol, carvone, citral, eugenol and ethyl cinnamate) as well as phenols (e.g., thymol and carvacrol) were considerably affected after GSH/ASA treatments (Table 5). The compounds were seriously affected more than any chemicals or vegetative characteristics assessed before. The fluctuation between the treatments was so far clear in this regard, since many cases of positive and negative effects or even losing activity were recorded throughout. Applying GSH and ASA either alone or in combination at different concentrations resulted in increasing OCMT by up to 37%, in the same time that MTHC were reduced by up to 59.85% (data deduced from the total value). Despite of the group they are belong to, linalool, D-limonene and, to a certain, camphene were the most affected components recording of that up to 139.35% increasing over control. On the other hand, the effect on increasing phenolic compounds was more constant than the case of monoterpenes components (e.g., MTHC; OCMT) and the maximum value was estimated with applying GSH and ASA alone (up to 82%). Applying the two compounds in combination was less efficient at all of their applications.
| Table 5: | Potential effect on the chemical constituents of the essential oil |
 | |
| Table 5: | Continued |
 | |
DISCUSSION
The keynote of applying GSH, ASA or their combinations seems to be achieved in this study. Remarkable achievements were obtained in reducing weed growth and increasing yield productivity in response to applying GSH, ASA or their combinations. No much significant results were observed between GSH and ASA in this connection. However, in opposite with ASA the capability of GSH on increasing marjoram growth or other attributes of yield and its components was stopped at the lowest and moderate concentrations only. Irreversible results were obtained at the highest concentration.
Increasing of the vegetative growth parameters of marjoram plants including plant height, number of branches per plant and shoot biomass (Fr.; Dr. Wts) as well as raising yield and its constituents could be partially or totally attributable to the physiological roles of such components as growth regulators, which have been established by several researchers. The GSH has a multifarious crucial positions attached in total with enhancing growth and development of growing plants. These include managing roots uptake for nutrients (i.e., sulphur), controlling cellular heavy metal concentration, managing protein synthesis and regulating cell division process (Grill et al., 1989; Herschbach and Rennenberg, 1994; Lappartient and Touraine, 1996; Sanchez-Fernandez et al., 1997). Talaat and Aziz (2005) referred that increasing the vegetative growth as well as oil productivity are two of the most distinguish features of applying GSH in aromatic plants e.g., Matricaria species. Of the most important rather than being involved in the above-mentioned physiological activities, GSH in concert with ASA has more precious functions in activating plant defense mechanisms against oxidative stress initiated by extremes of temperature, herbicides, or environmental pollution (Hausladen and Alscher, 1993). Chen et al. (2003) added that the role of GSH in protecting cells against toxic effects of free radicals is to keep the free radical scavenger ASA in its reduced form and hence active form.
Talking about the other possibility of enhancing marjoram growth due to weed reduction is the second point of interest that should be deal in more details. For several years studying, it is well-knowing the biocidal activity of marjoram's oil in suppressing growth and development of agricultural pests including insect and microbial diseases (Balbaa, 1989). Lately, weeds are involved as a part of this. Such unique property undoubtedly gives marjoram plants great advantage in their vicinity to face any of these troublesome pests. The mechanism of that is to be release certain allelochemicals in the surrounding media using one or more of the allelopathic avenues well known as volatilization, root secretion, leaching or residues decomposition (Rice, 1984). For enhancing such allelopathic capability within marjoram plants, it was the resolution of using GSH, ASA or their combinations, which seems to be achieved through greenhouse trials and confirmed in vitro via lab screening test. The chromatography analysis of oil components might provide the proof of the rightness of this fact, since the data revealed of more increasing in the terpene (i.e., OCMT) and phenolic compounds well known by their highly allelopathic action. The compatibility between the results obtained under greenhouse and lab screening tests absolutely ruled out competition as one putative cause of suppressing weeds accompanying marjoram. This confirming the above discussed point that something more likely to be a chemical interaction than competition could be behind this effect, scientists called allelopathy. This is the reason why field or greenhouse experiments connected to allelopathy should be appended with lab screening tests (as in our present study)-simply is to be supported and easily separated from any other involved factor such as competition or interacting micro-organisms (Olofsdotter et al., 1997). Despite of that, particularly with what actually achieved from vigorously and great enhancement of mass vegetative growth of marjoram herb in response to GSH/ASA foliar application, it is not logic to completely ignore competition as one potential reason for weed frailer. Even its participant role can’t be trustable, but it may have a significant role beside allelopathy in what actually happened under pot trials.
Like the idea of using the natural non protein amino acid, δ-aminolevulinic acid (ALA), it was the convergence with our study on using GSH and/or ASA. May be the mode of action and enhanced components were differed, but they have the same function by end in eliciting the phytotoxic compounds or probably producing another phytotoxic agent was not exist before in the receiving plants. Leaving aside the eliciting effect on oil components, many of these discovered chemicals in our study have already established herbicidal activity against wide range of mono and dicot plants including crop and weed species. However, their benefits as pharmaceutical agents far outweigh all positive herbicidal activities (Franthworth, 1988). Couladis et al. (2003) revealed that limonene, α-pinene, β-pinene and germacrene D were the main allelochemicals in turpentine (Pistacia terebinthus L.) tree and thanks for this discovery several phytotoxic symptoms were found a way for good explanation in many allelopathic species (Asplund, 1968). So far similar results were obtained on Ruta graveolens, either on its composition of the essential oil or the allelopathic ability which estimated up to 77% growth reduction in radish seedlings (Feo et al., 2002). Likewise, eucalyptus (E. citriodora) oils, the richest in limonene and cineole monoterpenes, the results were obtained on killing Phaseouls aureu (Singh et al., 1991). However, the factor seems to be common amongst these is that limonene, α-pinene and camphene were the most publicized and effective constituents and having more of its favor is to be used as a basic source for natural herbicides (Asplund, 1968).
The great reduction in broad and narrow-leaved weeds either in greenhouse or lab screening tests intimated that using such derivatives alone or in mixture might act in inducing the most bioactive phytotoxic agents in marjoram plants forward to that level is able to suppress associated weeds. The phenolic and terpene compounds are on the head of such components which established herbicidal activity (Duke et al., 2002). This come in reality particularly if we know that the marjoram oils are consist of about 70% of phenols and the remainder is to be of terpenoids (Sarer et al., 1982). There are many supporting evidences of increasing the essential oil components within the aromatic plants (e.g., Matricaria chamomilla and Tagetes minuta) in response to the foliar application by GSH or ASA (Balbaa, 2002; Talaat and Aziz, 2005). Probably, such increasing could be for some components at the expense of the others as documented in our study, but in common it is a strong visual evidence of the ability of these compounds to interact in a way or another in the metabolic pathway of the terpene and phenolic compounds and hence increasing the allelopathic capacity. Graham and Graham (1996) found that applying GSH led to increase the phenolic polymers content in soybean plant, which ultimately raised the competitive status of the plants in their media or agro-ecosystems (Armero et al., 2001).
Of the allelopathic phenomenon and its own allelochemicals-responsible components, phenolic and terpenoid substances are in the preference as they have constantly discovered in the plants containing highly allelopathic properties, not only in living plants, but also in their decomposed residues (Rashid et al., 2005; Singh et al., 2005). In this regard, Lydon and Duke (1989) reviewed many research reports of plant phytotoxicity and have the same conclusion by end that when a total or partial purification of a plant-derived phytotoxin is obtained, the active compound, in the majority of cases, is a phenolic compound (Harrison and Peterson, 1986; Kato-Noguchi et al., 2002). Assof et al. (1988) found that the phenolic compound named hydroquinone is one of the most efficient naturally-occurring compounds in marjoram extract. The compound has been adequately screened for being used as a natural herbicide; however, the cytotoxic effect was disappointed (Khan and Ungar, 1986). Kil (1989) reported that hydroquinone was amongst wide range of phenolic substances that possess allelopathic activity against pitch pine (Pinus sp.) trees. On the practical level, the post-emergence application of hydroquinone at 11.2 kg ha-1 was far promising for controlling redroot pigweed when used at 2-3 leaf stage, with the advantage set above others no harmful effects were noticed on the target crops e.g., corn or soybean (Shettel and Balke, 1983).
Terpenoids are larger and more implication in plant-plant interaction than phenolic substances particularly mono and sesquiterpenoid groups. There are considerable amounts of research regarding the herbicidal potential of terpenes, suggesting that many of them are quite phytotoxic and quite enough to be developed as natural herbicides (Duke, 1991). Sherman et al. (1983) indicated that the commercial herbicide sold under the trade name of toxaphene or camphechlor for controlling primarily sicklepod (Cassia obtusifolia L.) weed in soybean is just about the chlorinated form of the camphene monoterpene derivative. The only other terpene-related herbicide is cinmethylin. It is originally generated from the monoterpene molecule 1,8-cineol (Grayson et al., 1987). Reynolds (1987) found that applying p-cymene at 51 μM in vitro caused 50% growth inhibition in radish seedling growth. Asplund (1968) found similar results with applying pinene and limonene, but at rates of 45; 30 μM, respectively. These might explain the current findings of our study on the herbicidal activity of marjoram-containing terpenes.
It has been suggested that using GSH, ASA or their combinations for the purpose of increasing yields productivity and controlling weeds in aromatic plants might open the door for new strategy of the cleaning agriculture and sustainable development via minimizing our reliance on synthetic agrochemicals.
REFERENCES
- Armero, J., R. Requejo, J. Jorrin, R. Lopez-Valbuena and M. Tena, 2001. Release of phytoalexins and related isoflavonoids from intact chickpea seedlings elicited with reduced glutathione at root level. Plant Physiol. Biochem., 39: 785-795.
Direct Link - Chen, K., H. Gong, G. Chen, S. Wang and C. Zhang, 2003. Up-regulation of glutathione metabolism and changes in redox status involved in adaptation of reed (Phragmites communis) ecotypes to draught-prone and saline habitats. Plant Physiol., 160: 293-301.
Direct Link - DuBois, M., K.A. Gilles, J.K. Hamilton, P.A. Rebers and F. Smith, 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem., 28: 350-356.
CrossRefDirect Link - Duke, S.O., F.E. Dayan, A.M. Rimando, K. Schrader, G. Aliotta, A. Oliva and J.G. Romagni, 2002. Chemicals from nature for weed management. Weed Sci., 50: 138-151.
CrossRefDirect Link - De Feo, V., F. de Simone and F. Senatore, 2002. Potential allelochemicals from the essential oil of Ruta graveolens. Phytochemistry, 61: 573-578.
PubMed - Harrison, H.F. and J.K. Peterson, 1986. Allelopathic effects of sweet potatoes (Ipomoea batatas) on yellow nutsedge (Cyperus esculentus) and alfalfa (Medicago sativa). Weed Sci., 347: 623-627.
CrossRefDirect Link - Kato-Noguchi, H., T. Ino, N. Sata and S. Yamamura, 2002. Isolation and identification of a potent allelopathic substance in rice root exudates. Physiol. Planta., 115: 401-405.
CrossRef - Markham, R., 1942. A steam distillation apparatus for micro-kjeldahl analysis. Biochem. J., 36: 790-791.
Direct Link - Noctor, G. and C.H. Foyer, 1998. Ascorbate and glutathione: Keeping active oxygen under control. Annu. Rev. Plant Physiol. Mol. Biol., 49: 249-279.
CrossRefPubMedDirect Link - Olofsdotter, M., L.B. Jensen and B. Curtois, 2002. Improving crop competitive ability using allelopathy-an example from rice. Plant Breed., 121: 1-9.
CrossRefDirect Link - Pukacka, A. and E. Ratajczak, 2006. Antioxidative response of ascorbate-glutathione pathway enzymes and metabolites to desiccation of recalcitrant Acer saccharinum seeds. J. Plant Physiol., 163: 1259-1266.
CrossRefDirect Link - Rashid, A., N.H. Furness, B.E. Ellis and M.K. Upadhyaya, 2005. Inhibition of seed germination and seedling growth by hound's-tongue (Cynoglossum officinale L.) seed leachate. Weed Biol. Manage., 5: 143-149.
Direct Link - Rice, E.L., 1984. Allelopathy. 2nd Edn., Academic Press, Netherlands, ISBN: 0125870558, Pages: 422.
Direct Link - Sherman, M.E., L. Thompson and R.E. Wilkison, 1983. Sicklepod (Cassia obtusifolia) management in soybeans (Glysine max). Weed Sci., 31: 622-627.
Direct Link - Shettel, N.L. and N.E. Balke, 1983. Plant growth response to several allelopathic chemicals. Weed Sci., 31: 293-298.
Direct Link - Singh, H.P., D.R. Batish, J.K. Pandher and R.K. Kohli, 2005. Phytotoxic effects of Parthenium hysterophorus L. residues on three Brassica species. Weed Biol. Manage., 5: 105-109.
CrossRef - Snedecor, G.W. and W.G. Cochran, 1967. Statistical Methods. 6th Edn., Oxford and IBH Publishing Co. Pvt. Ltd., New Delhi, India, Pages: 593.
Direct Link - Yongqing, M.A., 2005. Allelopathic studies of common wheat (Triticum aestivum L.). Weed Biol. Manage., 5: 93-104.
Direct Link