
N. K. Bajaj
Department of Animal Reproduction, Gynaecoloy and Obstetrics, College of Veterinary Science and Animal Husbandry, (Madhya Pradesh Pashu Chikitsa Vigyan Vishwa Vidhyalaya, Jabalpur), Rewa, (M.P.), India
Neelesh Sharma
Division of Clinical Veterinary Medicine and Jurisprudence, Faculty of Veterinary Science and Animal Husbandry, Sher-E-Kashmir University of Agricultural Sciences and Technology of Jammu, R.S. Pura, Jammu-181102, (J and K), India
LiveDNA: 91.1660
Current Research in Dairy Sciences
Year: 2011 | Volume: 3 | Issue: 1 | Page No.: 1-24







ABSTRACT
Embryonic mortality denotes the death of fertilized ova and embryos upto end of implantation. Early embryonic mortality accounts for majority of reproductive failures with a mortality rate upto 40% of fertilized eggs. PGF2α secreted by endometrial cells by activation of specific receptors by oxytocin of luteal origin causes demise of corpus luteum. In bovines embryonic trophoblast secretes interferonι which constitutes major signals for maternal recognition of pregnancy having antiluteolytic effect and luteoprotective agents like PGE2 around day 8-9. Abnormal luteal function can be due to short luteal phase or abnormal luteal phase. Lack of exposure of the uterus to progesterone and estrogen prior to first postpartum ovulation decreases the concentration of progesterone and up regulation of oxytocin receptors which causes premature secretion of PGF2 α from uterus resulting in short luteal phase in most of cows. Milk, plasma and saliva progesterone concentrations, PAG test and ultrasonography can be used as diagnostic tool to know the embryonic mortality. Various treatment regiemes including use of hCG, GnRH, PMSG, interferon and supplementing Omega-3 fatty acids can be tried.
PDF Abstract XML References Citation
Received: August 07, 2010;
Accepted: September 07, 2010;
Published: April 29, 2011
How to cite this article
N. K. Bajaj and Neelesh Sharma, 2011. Endocrine Causes of Early Embryonic Death: An Overview. Current Research in Dairy Sciences, 3: 1-24.
DOI: 10.3923/crds.2011.1.24
URL: https://scialert.net/abstract/?doi=crds.2011.1.24
DOI: 10.3923/crds.2011.1.24
URL: https://scialert.net/abstract/?doi=crds.2011.1.24
INTRODUCTION
The success of diary enterprise is dependent on the milk production of dairy cattle and buffaloes (Bajaj et al., 2006a). Production is directly related to the reproductive phenomenon. The reproductive efficiency is affected by very well known factors like fertilization failure and embryonic mortality the later being more significant (Bajaj et al., 2006b). Embryonic death/mortality denote the death of fertilized ova and embryos upto the end of implantation (Jainudeen and Hafez, 2000). Mortality is more common during the early than the late embryonic period, i.e., from day 8th to 16th at the hatching of blastocysts and initiation of elongation and commencement of implantation without affecting cycle lengths. Early embryonic mortality is a major source of embryonic and economic loss with mortality rate upto 40% in animal production through repeat breeding and increased cost of artificial insemination (Sreenan and Diskin, 1986; Zavy, 1994; Bajaj, 2001), extended calving intervals and prolonged dry period resulting in reduced life time milk production (Roche et al., 1981) and reduced net calf crop (Maurer and Chenault, 1983). Earlier, it was believed that the bovine conceptus was resrobed, but transrectal ultrasound examination (Kastelic et al., 1991) had demonstrated that the conceptus and its breakdown products apparently is eliminated by expulsion through the cervix, which either goes unnoticed or appears as a vulval discharge of clear mucus.
When, the interestrus or the interovulatory intervals are extended in bred animals, it usually indicates embryonic loss that occurred around the period of Corpus Luteum (CL) maintenance (Van Cleef et al., 1991; Humblot, 2001). Measurement of progesterone in blood suggested that embryonic death at the time of CL maintenance delayed luteolysis and extended interestrus interval (Humblot, 2001). Therefore, when embryonic death precedes luteolysis, luteal regression is delayed by at least 3 days after the end of pregnancy (Kastelic et al., 1991). However, when luteolysis precedes and probably causes embryonic death, return to estrus are dependent upon the stage of follicle development. Humblot (2001) suggested that luteolysis and return to estrus prior to day 24 might be linked with early embryonic death (early embryonic loss) but if the CL is maintained and returns to estrus are delayed beyond day 24 and 24-49 (late embryonic loss). Pregnancy losses detected after day 50 known as fetal losses.
The cost of losses to farmers has been estimated as high as 250 million year-1 in the UK alone (Peters and Ball, 1995) and $1.4 billion in the USA (Hansela et al., 1976). Although, it is clearly not possible to extrapolate these figures to the whole world as national agricultural economics vary so widely, it can be seen that global cost of embryo mortality in a total cattle population (Bos taurus and Bos indicus) of approximately 1.28x1012 (FAO, 1994) is a large amount clearly justifying the relatively modest sums invested in research into its cause.
EMBRYO AND NORMAL EMBRYONIC DEVELOPMENT
An embryo is a product of fertilization characterized by growth and differentiation leading to the establishment of different organ systems that make up the individual.
Fertilization in the bovines occurs at ampullo-isthamus junction of the fallopian tube (Uterine tube) and the embryonic period can be divided into zygote, cleavage, morula, blastocyst, implantation and postimplantation. The zygote being the first structure formed as a result of successful fertilization cleaves mitotically into 2, 4, 8 and 16 cell stages. The cleaved embryo enters the uterine horn at the morula (mass of cells within the zona pellucida) stage about 4-5 days after fertilization. The morula then develops into blastocyst (having distinct blastocoele, trophoblast and embryonic disc) at days 6-7. The zona pellucida ruptures, resulting in hatching of the embryo after 9-10 days. The hatched blastocyst begins a process of elongation from about day 12-13, which is accompanied by the secretion of embryonic interferons. Early attachment (apposition) of the conceptus to the endometrium takes place from about day 19 and actual adhesion occurs by day 21-22.
Maternal recognition of pregnancy: Short (1969) firstly coined the term maternal recognition of pregnancy which means that the luteal function is maintained in early pregnancy and that the normal luteolytic mechanism is inhibited. In bovines mononuclear cells of trophectoderm secretes specific proteins (bovine trophoblastic proteins, later recognized as Interferon tau) (Thatcher et al., 2001) having anti-luteolytic effect (Danet-Denosyers et al., 1994) and luteal protective agents like PGE2 around day 8-9 (Bazer et al., 1994). Interferon-tau (IFNι) mediates its antiluteolytic effect by inhibiting expression of endometrial oxytocin receptors and by transduction mechanism after oxytocin-receptor binding on the endometrial cells thus inhibiting the episodic release of PGF2α (Demmers et al., 2001). Whenever, development of embryo is compromised or underdevelopment of trophectoderm, premature luteolysis occurs.
The ovine and bovine IFNι molecules have about 80% amino acid sequence homology whereas there is about 50% homology between IFN-α and IFNι (Roberts et al., 1992). Both ovine and bovine IFNι are secreted by the conceptus coincident with the blocking of luteolysis (Roberts, 1989).
Hernandez-Ledezma et al. (1992) indicated that bovine trophoblastic protein-1 (bTP-1/IFNι) production begins at the expanded blastocyst stage on day 8-9 just prior to the rupture of zona pellucida and hatching. They also suggested that embryonic mortality between day 14 and 19 is caused by luteal failure and occurs because certain embryos develop more slowly than normal and do not produce enough IFNι to prevent luteolysis and maintain the pregnancy.
Intrauterine infusion of recombinant ovine or bovine IFNι in on-pregnant cows extended oestrus cycle by abolishing oxytocin induced PGF2α secretion on day 17 and hence proved to be more effective in preventing embryonic mortality than IFNα (Meyer et al., 1995).
Martal et al. (1997) have reported many deleterious cytokines such as TNF-α, IFN-γ IL-2 and beneficial cytokines such as TGF-β, LIF, CSF-1, GMCSF, IL-1, IL-3, IL-6, IL-10 and IFNι are involved in embryo survival in ruminants and other species.
Kerbler et al. (1994) reported that an increased concentration of progesterone in cattle during early luteal phase is associated with enhanced embryonic production of the anti-luteolytic signal, IFNι. The CL that is maintained during maternal recognition of pregnancy is capable to convert PGF2α to its metabolite (PGFM) (Silva et al., 2000).
According to Spencer and Bazer (2004) establishment and maintenance of pregnancy results from signaling by the conceptus and requires progesterone produced by the Corpus Luteum (CL). Trophoblastic hormones in most of the mammals maintain progesterone production by acting directly or indirectly to maintain the CL. In domestic animals (ruminants and pigs), trophoblastic hormones maintain progesterone production by acting on the endometrium to prevent uterine release of luteolytic PGF2α.
DEATH OF THE EMBRYO AND TIME OF OCCURRENCE
Embryo death before day 13 of gestation results in return to estrus at the normal interestrus period. Death after day 13 extends the interestrus period beyond the generally accepted figure of 18-24 days.
Linares (1981) concluded that Early Embryonic Mortality (EEM) rather than fertilization failure was the major cause of repeat breeding in heifers as long as estrus detection and insemination had been properly performed. EEM is more important than fertilization failure in parous females in their relative contributions to reduced reproductive efficiency (Maurer and Chenault, 1983). According to Maurer and Chenault (1983), the critical period of embryo demise is day 7 of gestation when the morula develops into a blastocyst.
Markette et al. (1985) studied the incidence of embryonic loss following embryo transfer during the first week of development and concluded that between 50 and 75% of the embryos of poor quality are eliminated during the first 3 weeks.
Sreenan and Diskin (1986) reported that 20-25% of inseminations fail during the embryonic period, i.e., between day 1 and 42, most of the losses have occurred before day 25, the most substantial losses occurring between day 8 and 13 (8-9%) and days 14 and (13-15%).
Dunne et al. (2000) carried out studies on embryo and fetal loss in beef heifers between day 14, 30 of gestation and full term by measuring embryo survival rates which were 68, 76 and 71.8%, respectively. This led to the conclusion that most embryo losses in heifers occurs before day 14 after insemination.
CAUSES OF EARLY EMBRYO DEATH
Survival of embryo is affected by nutrition, temperature and heat stress, time of insemination, genital infections (Bajaj et al., 2006c), uterine environment and asynchrony (Bajaj, 2001), maternal age, genetic factors, immunological and endocrine factors. Endocrine factors/causes play an important role in embryonic death (Bajaj, 2001). To understand the influence of endocrine causes on embryo survival and mortality one should be familiar with the structural composition and functioning of corpus luteum and interaction between different reproductive hormones affecting luteal lifespan.
Cells of corpus luteum and their function: On the basis of morphology and biochemical properties corpus luteum is composed of two distinct steroidogenic cell types (Hoyer and Niswender, 1985):
| • | Small luteal cells (follicular origin): These cells derived from theca interna of preovulatory follicles (Priedkalns et al., 1968) |
| • | Large luteal cells (granulosa cell): These cells derive from granulosa cells (O’shea, 1987). Small luteal cells are able to differentiate into large luteal cells as the cycle progresses (Hansel and Dowd, 1986). Farin et al. (1988) postulated that conversion of small luteal cells to large luteal cells occurs only during the early part of oestrus cycle (Fig. 1) |
| • | Stem cells of corpus luteum: These different cell types contribute to circulating progesterone (P4) in different manner. Luteinizing Hormone (LH) is the major luteotropin in domestic ruminants (Niswender et al., 1985) and cattle and there is marked difference in the response to LH by large and small luteal cells (O’shea, 1987) |
Harrison et al. (1987) reported that small luteal cells possess higher number of LH receptors than large luteal cells. Niswender et al. (1985) found that 20% of the progesterone (P4) in the ovarian vein in mid-cycle is secreted by small luteal cells while nearly 80% of P4 is secreted by large luteal cells which have only few functional receptors for LH. O’shea (1987) indicated that on per cell basis large luteal cells produce more progesterone than small cells.
Luteal function during early pregnancy: Plasma and milk progesterone concentration rise similarly in early luteal phase in pregnant and non-pregnant animals but the higher concentrations are maintained in pregnant cows for duration of pregnancy which are essential for mantainence of pregnancy. Anti-luteolytic substances secreted by embryo around day 13 are probably responsible for differences in progesterone patterns between pregnant and nonpregnant animals.
 | |
| Fig. 1: | Conversion of luteal cells |
Lamming et al. (1989) found that milk progesterone concentrations in pregnant and nonthpregnant cows rise indifferently until day 9, which later on diverged and the concentrations in pregnant cows remained higher. They also reported a significant dip in progesterone concentrations in pregnant animals on day 11, followed by a rise which reflects a rescue effect of the corpus luteum by the embryo.
Shelton et al. (1990) reported that the rate of rise in progesterone concentration was lower in the postovulatory period in cows identified for sub-fertility than in pregnant and nonpregnant heifers.
Role of oxytocin, oxytocin receptors (OTR) in luteal function: Wathes and Lamming (1995) have reported that during luteal regression, pulses of oxytocin stimulate synthesis and pulsatile release of PGF2α following an increase in endometrial oxytocin receptors (OTR). Oxytocin receptor synthesis and PGF2α release are inhibited by interferon production by the conceptus during early pregnancy. They also reported that OTRs are present during anoestrus, oestrus and late luteal phase and during most of pregnancy while the plasma oxytocin concentration causes parallel changes in plasma progesterone. These concentrations are basal at oestrus and rise from about day 2 of the cycle peaking around day 9 and falling from about day 12-13 before the onset of luteolysis (Wathes et al., 1993). The pattern is similar in the pregnant animal and plasma concentrations fall from 12-13 days after inseminations.
Plasma concentrations of PGF2α are generally low for most of the cycle but pulsatile release starts at day 13 and the pulse frequency increases until luteolysis (day 17). In nonpregnant ewes most oxytocin pulses occur in association with PGF2α whereas, in pregnancy most pulses are not associated with PGF2α (Hooper et al., 1986). This pattern might be similar in cattle.
Progesterone blocks the increase in OTR for about the first 10-12 days of the luteal phase, but the mechanism of its prolonged effect, followed by upregulation, is still unclear. McCracken et al. (1984) found that increasing progesterone concentration in the luteal phase down regulates the progesterone receptor for about 10 days. But there was no evidence whether the progesterone receptors increase prior to the OTR increase or the treatment with progesterone reduces its own receptor (Wathes and Lamming, 1995).
Interferon, oxytocin receptors interaction and luteal function: During early pregnancy the rise in OTR concentration is inhibited probably by the action of embryo derived IFN. The mechanism of action of IFN may involve suppression of both oestradiol and oxytocin receptors probably at the transcriptional level (Bazer et al., 1994). Bovine Interferon-alpha (bIFNα) has been shown to stimulate progesterone production by luteal cells in vitro, without affecting oxytocin output (Luck et al., 1992). Imakawa et al. (1993) reported that maternal granulocyte macrophage colony stimulating factor might be involved in stimulating embryo IFN production.
Types of abnormal luteal function: Abnormal luteal function is associated with reduced pregnancy rates (Hommeidaa et al., 2004). Two distinct type of abnormal luteal function have been reported (Troxel and Kesler, 1984):
Type-I (short luteal phase): It is observed after a period of sexual rest and when breeding is initiated for the first time. In this condition short life span of corpus luteum (6-12 days) is observed.
Type-II (inadequate luteal phase): It is observed at any stage during the reproductive life. Life span of corpus luteum is of more than 14 days but with depressed plasma progesterone.
Mechanisms contributing to reduced luteal function: Mechanism contributing to reduced luteal function can be classified into three main categories:
| • | Deficiencies in the maturational process within the preovulatory follicle and/or inadequacies of ovulatory stimulus |
| • | Short comings in the support of the CL once they have formed |
| • | Premature activation of the luteolytic process |
Follicular deficiencies may reflect in the form of subnormal progesterone concentration and short lived CL may be the consequence of premature activation of the luteolytic process.
WHY IS THE FIRST POSTPARTUM OVULATIONS RESULTS IN SHORT CYCLES?
During early postpartum period when the reproductive system is recovering to its normal anatomical and functional status, especially ovaries (ovarian rebound), presence of dormant follicles in ovaries and adequate pattern of LH secretion leads to first postpartum ovulation after few weeks (Perea and Inskeep, 2008) but the fertility in the first postpartum oestrus is poor as compared with subsequent cycles. During normal luteal phase of estrous cycle progesterone (P4) causes increase in estrogen concentration to promote synthesis of progesterone (P4) receptors (Zollers et al., 1993) and to inhibit synthesis of uterine receptors for oxytocin (OTR) through the mid-luteal phase (Kieborz-Loos et al., 2003). But lack of priming of uterus for progesterone followed by estrogen prior to first ovulation causes decrease in progesterone receptors and up regulation of oxytocin receptors which results in premature PGF2α secretion (Cooper et al., 1991). During the transition period from morula to blastocyst, i.e., during first 2 weeks of insemination (Inskeep, 2002) maternal recognition of pregnancy is prevented by premature release of PGF2α from uterus and luteal origin resulting in early demise of CL (Cooper et al., 1991; Inskeep, 2004). Since, the PGF2α is secreted prematurely the exogenous supplementation of progesterone is also ineffective in prevention of embryonic death.
Theoretically, CL could be influenced by two tropic stimuli:
| • | Increasing the CLs secretory activity |
| • | Prolonging CLs life span |
Deficiencies prior to ovulation: Proper maturation of preovulatory follicle is essential as follicle after maturation results in CL formation (Garverick et al., 1988). Gonadotropin treatment prior to the induction of ovulation results in improved functioning of resulting CL (Sheffel et al., 1982). Changes within maturing follicle may influence functional CL in the form of decreased cell numbers, cell sizes or proportions of large to small luteal cells (O’shea et al., 1984) or deficiency of receptors to luteotropin could result in the CL failing to recognize or respond luteotropin (Rutter and Randel, 1984). Increased concentrations of preovulatory LH could promote secretion of P4 via cells of granulosa or thecal origin (Fitz et al., 1982), with the evidence favouring an effect of LH on the small luteal cells.
Why are the mature follicles scarce at the time of sexual rest?: During the period of anoestrum or sexual rest No. of mature follicles is less due to low plasma LH concentration (Nett, 1987). This was partly the result of low pituitary reserves of LH during the early postpartum period consequently, the deficiencies were considered to include inadequate release of LH, either for several days prior to ovulation, or in short coming (duration and/or peak concentration) of preovulatory surge of LH.
Hormonal mechanisms involved in follicular maturation: The final stages of follicular maturation are associated with marked increase in the frequency of LH episodes (Karsch et al., 1983). The frequency of LH increases from one per 3 to 12 h from luteal regression to 1 to 2 immediately per LH peak (McLeod et al., 1982). During ovarian acyclicity these LH frequency were low (Savio et al., 1990). Thus, during the early postpartum period in dairy cows, the pulsatile release of LH was insufficient to promote the final maturation of follicles (Hunter and Southee, 1987). To support follicular maturation a pulsatile delivery of LH to the ovary may be preferable to constant exposure (McNatty et al., 1981). However, widely differing patterns of LH stimulation during the pre-ovulatory period have resulted in normal follicular maturation and some CL function (Keisler et al., 1985). PGF2α may also be involved in follicular development in early postpartum cows (Guilbault et al., 1987). Accordingly, either the quantity or pattern of PGF2α release is inappropriate prior to ovulation resulting in subnormal CL function.
Why LH pulses infrequent during early postpartum?: Lowered pituitary concentration of LH is one of the limitations to the re-establishment of oestrous cycles rather than alterations in the sensitivity of the organ to GnRH (Moss et al., 1985). Nett (1987) suggested that deficiencies of the hypothalamic hypophysial axis could result in a reduced secretion of LH during the early part of the postpartum period.
Nett (1987) concluded that during the postpartum period the hypothalamus although, contained sufficient stores of GnRH to stimulate the anterior pituitary. Once the pituitary stores of LH had been replenished, then pulses in the secretion of LH could increase in frequency to culminate in the first postpartum oestrus (Humphrey et al., 1983). Ovarian acyclicity thus appeared to be the result of a failure of follicle development, possibly due to an inadequate frequency of LH pulses reflecting inadequate pulsatile release of GnRH (Wise et al., 1989).
Feedback response to estrogen: Peters (1984) has proposed that although, the positive feedback mechanism of E2 in triggering the preovulatory LH release may be functional, recovery of maximal activity may continue over an extended period. Therefore, the hypothalamic-pituitary axis would need to recover responsiveness to increasing plasma concentrations of E2 before the first ovulation can occur.
Parfet et al. (1986) stated that the absence of oestrous cycles in suckled cows near 30 days postpartum was not due to deficiencies in:
| • | Ovarian follicular development |
| • | Anterior pituitary concentration of LH and FSH or in vitro releasibility of LH or |
| • | Pituitary receptors for GnRH |
In the properly fed cow, at about one month after calving, the hypothalamic-pituitary axis is fully able to support resumption of ovarian cycles, but the suckling stimulus inhibits the release of LH (Williams, 1989).
Is FSH deficit during early postpartum period?: FSH plays a permissive role in the onset of ovarian cycles postpartum. Deficiency of FSH during the late luteal to early follicular phase could inhibit the development and function of the preovulatory follicle (Fortune and Quirk, 1988).
FSH concentrations are lower over the last four days before the first preovulatory LH surge induced by weaning in cows. FSH concentrations were also lower compared to the same period prior to second preovulatory LH surge (Remirez-Godinez et al., 1982). The former preceded short lived CL, while the later preceded normal luteal function.
Lack of follicular maturation in the postpartum cow: Nett (1987) used a model to explain the inadequate maturation of follicles in the postpartum, anoestrous cows. For low concentrations of LH during early lactation he suggested that during pregnancy the high concentrations of P4 and E2 resulted in a prolonged negative feedback on the hypothalamic-hypophyseal pituitary axis. Accordingly, the synthesis of LH was inhibited and pituitary stores become depleted so that basal release of LH was reduced. After parturition, a two phase recovery of the hypothalamic-pituitary gonadal axis occurred with the first phase (lasting 2-5 weeks) characterized by infrequent release of GnRH (one pulse/4-8 h). Once pituitary stores of LH had been replenished then the amplitude of the LH pulses was sufficient to stimulate follicular growth. This denoted the start of second phase of the recovery process, during which the increased circulating concentration stimulated growth of ovarian follicles, which in turn produced E2. At this point of time, the frequency of release of GnRH also increased with a consequent increase in the frequency of LH pulses. The final stages of follicular development ensued and culminated in the first ovulation.
Intrafollicular receptors for LH and the role of estrogen (E2): Even if luteotropin secretion is sufficient to support normal CL function, then short lived or inadequate CL may not recognize LH. This might be due to insufficient LH receptors possibly within the maturing follicle and in the subsequent CL. Both E2 and FSH are necessary for production of LH receptors before luteinization of granulosal cells (Richards, 1980).
According to Fortune and Quirk (1988), estrogen (E2) also acts within the follicle to regulate its development and function. They proposed that as the preovulatory follicle mature the initial action of E2 of positive feedback is on its own production, via increased androgen synthesis.
During preovulatory growth, follicles become more responsive to LH and acquire an increased ability to synthesize E2. This increased responsiveness to basal levels of LH was due to increased LH receptors in the granulosal cells.
Estrogen (E2) and luteal lifespan: Larson (1987) reported that administration of hCG (to induce ovulation) leads to CL with a normal lifespan in cows with high plasma E2 than in cows with low E2. Inskeep et al. (1988) reported largest preovulatory follicle possessed decreased numbers of receptors for LH in granulosa and theca cells in cows which were predicted to have short luteal phases than in Norgestomet pretreated cows which were expected to show normal luteal lifespans.
Garcia-Winder et al. (1987) and Inskeep et al. (1988) found a higher concentration of E2 in largest follicle and CL with normal life span in cows implanted with synthetic progestogen (Norgestomet). Evidences from number of sources support that estradiol within the preovulatory follicle is associated with function of resulting CL.
Deficiencies at the time of ovulation/postovulation
Could abnormalities of preovulatory LH surge result in a short lived Cl or defective CL?: Troxel and Kesler (1984) stated that the magnitude and duration of GnRH induced LH surge appeared to be associated with enhanced CL function and lifespan. Cruz and Kesler (1988) reported that cows with normal luteal function had a greater GnRH induced LH release than cows with short luteal phases.
Shirar et al. (1989) emphasized the importance of duration of the preovulatory LH release. They observed that progestogen treatment, prior to GnRH, changed the pattern of LH release and possibly thereby improved luteal function.
Can the reduced lifespan or lowered secretory activity of the first CL be attributed to suboptimal luteotropic support?: Manns et al. (1984) reported the presence of FSH receptors in luteal cells of cows. Walters and Schallenberger (1984) suggested that since 97% of separate FSH pulses during the mid-luteal phase in cow were associated with P4 pulses, it is possible that FSH could be the principal hormone that stimulates P4 secretion. They maintained that this did not exclude a luteotropic action of LH in addition to a stimulatory action of FSH on release of P4 in the cow. Pekala et al. (1983) proposed that there might be an interaction between oxytocin (from the CL) and LH in regulating P4 synthesis, with oxytocin increasing the response to LH.
Mallory et al. (1986) reported premature luteal regression or reduced CL function if the luteotropic support from pituitary is disrupted during the formation of CL (±day 1 of the cycle).
Deficiencies in luteotropic support: Deficiency in luteotropic support might be due to:
| • | Short lived CL |
| • | Inadequate CL |
Short lived CL: Several studies have been carried out on insufficient luteotropic stimulus to maintain lifespan of subsequent CL during early postpartum period. Injection of microencapsulated GnRH promote normal luteal function by elevating LH concentration (Roberts et al., 1989).
Inadequate CL: If it is accepted that luteotropin may not be sufficient to realize normal luteal function, then the question arises that when should the endogenous supplies need to be augmented.
Pearson and Lishman (1989) demonstrated that a luteotropic stimulus provided early in the induced cycle (day 3-5) improved luteal function to a greater extent than when PMSG was provided after day 5.
Progesterone concentration on day 10 after AI were higher in pregnant buffaloes than in buffaloes that showed embryonic mortality not associated with infectious agent. Similarly higher progesterone concentrations on day 20 of AI were found in pregnant buffaloes as compared to non-pregnant ones and buffaloes that showed embryonic mortality (Campanile et al., 2005).
LH receptors within the CL and luteal function: Increased numbers of receptors for LH, on the granulosal cells of the bovine CL, could be a major factor in regulation of secretion of P4 by the ensuing CL (McNeilly et al., 1981). They also believed that there may not be a simple relationship between the binding of LH to luteal cells and the secretion of P4. Consequently, the failure of the short lived CL to recognize LH was not a factor that caused early luteal regression (Rutter et al., 1985).
LH receptors are not deficient in the inadequate CL (Braden et al., 1989). Hunter et al. (1988) concluded that lack of gonadotropin receptors was probably not a fundamental cause of premature regression of the short lived CL.
The role of the uterus and PGF2α in subnormal luteal function
Presence of PGF2α during early lactation: Thatcher et al. (1980) have demonstrated increased jugular plasma concentrations of prostaglandin metabolite (PGFM) during the early postpartum interval in milked dairy cows. Odde et al. (1980) suggested that premature regression of CL, which appeared to function normally for approximately 7 days after induced ovulation, was due to the luteolytic effect of PGF2α needs consideration.
Oxytocin and the release of PGF2α: Luteolysis occur due to positive mechanism between luteal oxytocin and uterine PGF2α which occurs at specific time of the oestrus cycle. Progesterone suppresses PGF2α secretion when the concentration of progesterone receptors is high by inhibiting oxytocin in early part of oestrus cycle but as the cycle progresses these progesterone receptors decreases resulting in secretion of estrogen and oxytocin receptors. This result in uterine responsiveness to oxytocin which further facilitates positive mechanism between oxytocin and endometrial PGF2α. Various research studies conducted on normal oestrus cycles suggests that progesterone regulates the PGF2α release by timing the initial peaks of secretion early in the oestrus cycle and later on by modulating the PGF2α secretion until the corpus luteum regresses completely.
Increase in oxytocin concentrations in association with increase concentration of PGFM in cows with short luteal phases has also been reported by Peter et al. (1989). Zollers et al. (1989) compared short and normal luteal phases in beef cattle and found oxytocin induced premature release of PGF2α on day 5 in short cycle which was not seen in normal cycles but the oxytocin induced PGF2α release was similar to that observed at luteolysis on day 16 of normal cycle. Lesser No. of progesterone receptors as compared to more No. of endometrial oxytocin receptors were reported on Day 5 after estrus in postpartum beef cows expected to have short luteal phase than in cows with normal luteal phase (Zollers et al., 1993). Evidences indicate involvement of oxytocin in release of PGF2α in cows with short luteal cycles.
Research studies on anoestrus cows have suggested that progesterone treatment results in development of larger preovulatory follicles containing more esrtadiol 17-β than in control or short cycled cows and greater concentration of LH receptors in granulosa and theca layer in largest preovulatory follicle in postpartum cows pretreated with norgestomet than controls (Inskeep et al., 1988). Increase in concentration of estradiol is associated with induction of endometrial progesterone receptors (Ing and Tormesi 1997). Cooper et al. (1991) and Kieborz-Loos et al. (2003) reported lower concentration of estradiol secretion by preovulatory follicle in short cycled cows will result in less No. of progesterone receptors in uterine endometrium and the first rise in progesterone and early PGF2α secretion with short luteal phases.
Oxytocin administration during the early part of the cycle has short luteal lifespan in cycling cows and it was suggested that oxytocin functioned via elevated levels of uterine PGF2α (Milvae and Hansel, 1980) in a manner similar to that of an IUD inserted early in the cycle in the ewes. Suckling has same effect via the pituitary release of oxytocin (Troxel and Kesler, 1984). When the CL releases oxytocin, it may cause PGF2α release which in turn regresses the CL (Schams et al., 1985). An imbalance in luteal levels of oxytocin was proposed as a cause of the eventual demise of the CL. Peter et al. (1989) were able to detect parallelism between concentration of PGFM and oxytocin in early postpartum dairy cows. Inskeep (1995) reported oxytocin to be an important factor that plays an important role in embryonic death of cows with short luteal phases. While McCracken et al. (1984) had earlier reported that luteal oxytocin and uterine prostaglandin facilitate luteolysis through reciprocal positive feedback mechanism. Zollers et al. (1993) reported that concentration of endometrial oxytocin receptors are greater at day 5 after estrus in cows with short lifespan of CL than in cows receiving progesterone and are expected to have normal lifespan.
Niswender et al. (2007) concluded that luteal PGF2α is involved in luteolysis in sheep. According to him uterine PGF2α initiates increased secretion of PGF2α by corpus luteum and it also initiates mobilization of Ca++ in large luteal cells and secretion of oxytocin in large luteal cells. Oxytocin secreted by large luteal cells acts on small luteal cells and triggers mobilization of Ca++ in small luteal cells. Increase in intracellular levels of calcium leads to apoptotic death of the large and small luteal cells.
Negating the effect of PGF2α: Involvement of PGF2α in premature regression of the CL is supported by the observation that treatment with substances which block the action of PGF2α increased the functional lifespan of CL destined to be short lived in dairy cows (Dobson et al., 1987).
Removing the source of PGF2α (experimentally): Strong evidence that a premature release of PGF2α from the uterus could be responsible for the early demise of the first induced CL in cows has been provided by Copelin et al. (1987). They demonstrated that the CL formed after early weaning of calves did not possess an inherently short lifespan because removal of the uterus did not result in premature luteolysis. Wright et al. (1988) showed that the previously gravid uterus had to be present to induce premature regression of the first CL, subsequent to induced abortion in heifers.
Demonstrating the release of PGF2α: Peter et al. (1989) showed increased concentrations of metabolite of PGF2α (15 keto-13, 14 dihydro [PGFM] PGF2α) were detected early in the cycle when, the CL was short lived. Schrick et al. (1993) found greater concentrations of PGF2α with lower embryo quality in uterine flushing of cows with short lived CL than normal luteal phases on day 6 and concluded that detrimental uterine environment was major cause of lowered fertility in cows with short luteal phases. Buford et al. (1996) concluded that inhibition of prostaglandin synthetase and lutectomy prevents detrimental effects of endogenous PGF2α on the early embryonic stage in postpartum cows whereas lutectomy alone was effective in doing so in non-lactating cycling cows. Therefore, the lower survivability of embryos in postpartum cows might to be due to the combined effect of uterine PGF2α secreted on Days 4 through 9 (Cooper et al., 1991) and luteal secretion of PGF2α in response to the uterine signal (Buford et al., 1996). Hockett et al. (2004) reported that PGF2α might interfere with process of compaction of morula stage of embryo resulting in lower quality and viability of embryos.
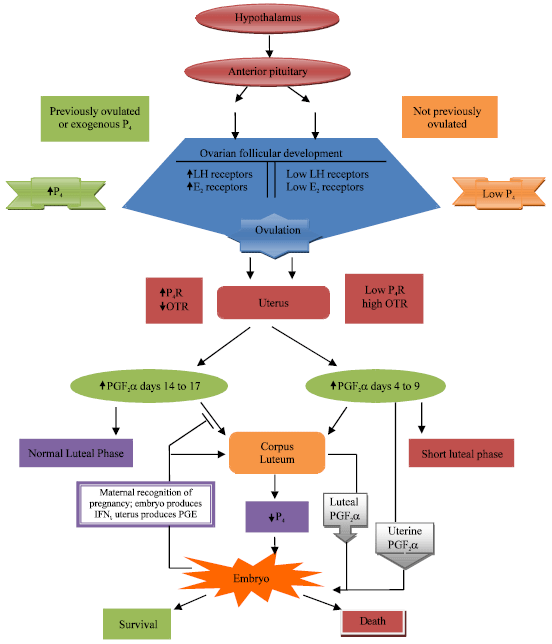 | |
| Fig. 2: | Sequence of endocrinal events conducive to embryonic death or survival in short and normal estrous cycles (Perea and Inskeep, 2008) |
This might occur as alterations in gap junctions and cell adhesion molecules, induction of apoptosis mechanisms or alterations of gene transcription during embryonic development in compaction morula stage of embryo (Scenna et al. 2004).
Perea and Inskeep (2008) described the sequential endocrine events conducive to embryonic death or survival in short and normal estrous cycles (Fig. 2). According to them after parturition, the hypothalamic-pituitary axis stimulates progressive ovarian follicular development, estradiol production and expression of LH receptors in follicle. Without a previous cycle or exposure to progesterone (P4)/progesterone priming followed by estrogen (E2), uterine progesterone receptors (P4R) are low and increase in progesterone after ovulation initiates immediate secretion of uterine PGF2α which causes luteolysis and, if the animal has been mated, it is embryotoxic. Exposure of the uterus to progesterone followed by estrogen prior to first postpartum ovulation up regulates uterine progesterone receptors and contributes to establishment of progesterone dominance of the uterus. As the estrus cycle progresses progesterone binding to its receptors delays uterine secretion of PGF2α. If the animal is not mated/pregnant luteolysis is initiate by uterine secretion of PGF2α that starts around day 14 to 17 and decreases progesterone production. If the animal is pregnant, the embryo produces Interferon-tau (IFNι) while the uterus increases PGE and the corpus luteum is maintained allowing the embryo to survive. The whole process is known as maternal recognition of pregnancy.
DIAGNOSIS
Early embryonic deaths before regression of CL are indistinguishable from fertilization failure in that both cow and ewe return to estrus at the normal time. Death of one embryo in twin ovulating ewes may be undetected as pregnancy will continue. Several methods are used to determine embryonic mortality in cattle. The main being:
Examining embryos: Examining embryos collected by in vivo flushing of reproductive tract at different days after breeding.
Determining progesterone in blood, milk and saliva: Determination of progesterone in blood and milk, 21 days after oestrus is the most common method used in the pregnancy diagnosis of ruminants until the early nineties (Zoli et al., 1992; Karen et al., 2003). Prvanovic et al. (2009) in his study on monitoing of early pregnancy and embryonic mortality using blood progesterone concluded that it is impossible to determine embryonic mortality alone on the basis of progesterone profile while pregnant and non-pregnant cows can be easily distinguished 21 day post AI. They also concluded that it is very easy and accurate to distinguish non-pregnant cows from cows that have suffered early embryonic mortality.
CL dysfunction in human reproduction is identified by either of two criteria namely estimation of plasma or salivary P4 levels or endometrial biopsy. Two or three plasma samples could be processed for the assay. Progesterone levels should not be less than 10 ng/ml during the luteal and mid luteal phase.
Pregnancy associated glycoprotein (PAG) test: The main advantage of Pregnancy Specific Proteins (PSP) for pregnancy diagnosis in cattle is their ability to prove the existence of placentation and the presence of live, vital embryos, while progesterone only proves the existence of corpus luteum. The most commonly used pregnancy specific protein for pregnancy diagnosis in cows is PAG (pregnancy-associated glycoprotein). Pregnancy-associated glycoprotein has been found in the serum of pregnant cattle and used as a pregnancy marker (Perenyi, 2002). As pregnancy failure occurs, PAG concentrations drop and disappear from maternal blood. The Pregnancy-Associated Glycoproteins (PAG) are synthesized by the monothand binucleate cells of the ruminant's trophectoderm. Apart of it is released into maternal blood circulation which can be assayed by RIA and ELISA. RIA methods are very precise for measuring PAG concentrations in the maternal blood and milk of the ruminants. The sensitivity and specificity of this method are very high. The results are encouraging and use of milk and in blood for PAG test is helpful in detection of embryonic mortality in the ruminants (Sousa et al., 2008). Prvanovic et al. (2009) in his study on monitoing of early pregnancy and embryonic mortality using PAG test concluded that embryonic mortality between 18-24 days after AI was evident from drastic decrease in PAG seven and half to nine days later and using PAG for pregnancy diagnosis enables us to prove the existence of live, vital embryos in utero 24 days after conception.
Ultrasound examination: Transrectal ultrasonography has been used to detect early pregnancy and to determine embryo/fetal death in recent years (Kahn, 1992; Romano and Magee, 2001). It is advantageous as it is a safe technique with no effects on embryo/fetus viability (Kahn, 1992; Ball and Logue, 1994; Baxter and Ward, 1997). It is advantageous over palpation per rectum pregnancy diagnosis in earlier diagnosis of pregnancy/non-pregnancy, determination of embryo/fetus viability, determination of number of embryos, reduction of misdiagnosis (false negatives and false positives) and reduction of potential iatrogenic embryo/fetus attrition (Romano and Magee, 2001). In research studies maximum sensitivity and negative predictive values were obtained from day 29 on in cows and from day 26 on in heifers.
TREATMENT TO IMPROVE PREGNANCY RATES
Various methods have been tried using different preparations for improving pregnancy rates by reducing embryonic mortality. They are as under:
Supplementing progesterone/progestogen: Research studies have reported that low concentration of progesterone can result in the development of a stronger luteolytic signal and hence it might be concluded that cows with lower plasma concentrations are apparently more prone to embryo loss. Macmillan and Peterson (1993) found that the conception rates to first insemination were increased when the CIDR device was inserted 6-8 days after insemination. Broadbent et al. (1992) recorded a significant increase in conception rate among cattle when Crestar ear implants (Norgestomet) were given on day 7.
Peters and Ball (1995) stated that progesterone supplementation might also cause suppression of endogenous luteotropic support due to increased negative feedback. While, Mann and Lamming (1999) demonstrated that supplemental progesterone was beneficial to fertility increasing conception rates when administered prior to day 6 after AI in lactating dairy cows.
Use of human chorionic gonadotropin (hCG): An alternative approach to increase the progesterone levels is by use of Human Chorionic Gonadotropin (hCG) to enhance the production of progesterone by the animals own corpus luteum. Administration of human chorionic gonadotropin (hCG) induces ovulation with the subsequent formation of a functional accessory CL which in turn increases progesterone and may enhance embryo survival. Studies have supported that conception rate is better in cows with three follicular waves after insemination as compared to cows with two follicular waves and hCG induction of three-wave cycles may also contribute to higher pregnancy rates. Thatcher and Collier (1986) and associates have indicated that the injection of hCG (2000 IU) i.m.; (1000 IU) i.v. 5 days after oestrus induces ovulation of the first wave dominant follicle and formation of accessory corpus luteum and increases plasma progesterone levels during the luteal phase. Santos et al. (2001) demonstrated that injecting 3300 IU of hCG in lactating cows 5 days after AI resulted in increased number of CL and higher plasma progesterone concentrations. Conception rates on days 28, 42 and 90 were improved by hCG treatment. The findings of Santos et al. (2001) were supported by findings of Nishigai et al. (2002), they administered hCG on day 6 and the pregnancy rates were increased (67.5%) with formation of accessory corpora lutea as compared to control cows (45.0%). or cows receiving hCG on day 1 (42.5%) after Lopez-Gatius et al. (2002) reported that cows having an additional spontaneous CL were eight times less prone to fetal loss than those with a single CL.
PMSG (pregnant mare serum gonadotropin) administration: Hirako et al. (1995) in their study on luteotropic effect of PMSG in cattle and reported significant increase in progesterone concentration on administration of 500 IU of PMSG on day 7 after estrus. They concluded that PMSG treatment increases progesterone secretion and luteal function without excessive follicular development.
Gonadotropin releasing hormone (GnRH) treatment: Administration of GnRH (250 μg or greater) at the time of insemination increases pregnancy rates by 12.5% and effect was more pronounced in repeat breeder cows (22.5%) (Morgan and Lean, 1993). Administration of GnRH at oestrus increases serum progesterone levels and the proportion of large luteal cells in the corpus luteum (Mee et al., 1995).
MacMillan et al. (1986) reported an enhancement of the conception rate in dairy cows when GnRH (10 μg Buserelin) was injected on day 11 after breeding by AI. Injecting Buserelin at 3 days interval from day 12 (luteal phase) increases progesterone concentrations and maintains at luteal levels till the injections are continued, i.e., until day 48 after the preceding oestrus (Thatcher et al., 1989). This indicates that buserelin exerts a continued luteotropic or antiluteolytic effect under these circumstances.
Administration of hCG during luteal phase of the cycle induces ovulation of the dominant follicle (Price and Webb, 1989). Administration of GnRH on day 6 of the cycle resulted in ovulation in 75% animals with the formation of accessory corpora lutea (Webb et al., 1992). This not only induces additional progesterone secretion, but also downregulates oestradiol production.
Mann et al. (1995) reported that administration of buserelin on day 12 after insemination results in reduced plasma oestradiol concentrations and suppression of pulses of the PGF2α metabolite (13, 14-dihydro-15-keto-PGF2α (PGFM) from about day 13 onwards, confirming its antiluteolytic effect. Reduction in oestradiol concentrations at this time might inhibit the luteolytic mechanism and hence, pregnancy is maintained. They also concluded that GnRH treatment weakens rather than delays the luteolytic signal. The timing of buserelin/GnRH treatment appears critical since treatment at other times after insemination did not have any effect (MacMillan et al., 1986; Drew and Peters, 1994).
Interferons: Intrauterine administration of recombinant ovine/bovine IFNι resulted in extension of the oestrous cycle and abolished the oxytocin induced PGF2α secretion on day 17 (Meyer et al., 1995). IFNι is more effective than IFNα in preventing embryonic mortality as it has no side effects.
Omega-3 and reproductive performance: Research studies by workers (Mattos et al., 2000, 2002; Thatcher et al., 2001) have indicated that omega-3 fatty acids decreases secretion of PGF2α. Trials by research workers (Bonnette et al., 2001; Mattos et al., 2002; Petit and Twagiramungu, 2002; Ambrose and Kastelic, 2003) with natural sources of omega-3 fatty acids such as Eicosapentaenoic Acid (EPA), Dehydroascorbic Acid (DHA) and α-linolenic acids have indicated that these fatty acids are capable of decreasing the secretion of PGF2α and compliment the antiluteolytic action of IFNι thereby improve pregnancy rates. EPA and DHA also have anti-inflammatory and immunosuppressive effects that compliment the normal immunosuppressive and anti-inflammatory effects of progesterone and IFNι in early pregnancy.
CONCLUSIONS
EED is the major cause of reproductive and economic loss in cattle. Though many factors have been incriminated but luteal function plays an important role. Embryonic death occurs the time of maternal recognition of pregnancy, probably related to a failure of the IFNι secretory mechanism. Recent research, both in terms of physiological mechanisms and pharmacological treatments has mostly focused on the period of maternal recognition of pregnancy or the anti-luteolytic effect. Ovarian examination, blood/milk progesterone levels, PAG test and ultrasound appear to be the only practical tool presently available. hCG/GnRH/P4 supplementation have shown positive results. Supplementation of interferon as anti-luteolytic agent and supplementing Omega-3 has shown encouraging results.
REFERENCES
- Ambrose, D.J. and J.P. Kastelic, 2003. Dietary fatty acids and dairy cow fertility. Adv. Dairy Tech., 15: 35-47.
Direct Link - Baxter, S.J. and W.R. Ward, 1997. Incidence of fetal loss in dairy cattle after pregnancy diagnosis using an ultrasound scanner. Vet. Rec., 140: 287-288.
Direct Link - Bazer, F.W., T.L. Ott and T.E. Spencer, 1994. Pregnancy recognition in ruminants, pigs and horses: Signals from the trophoblast. Theriogenology, 41: 79-94.
CrossRefDirect Link - Bonnette, T.R., J.C. Whittier, T.E. Engle and P.D. Burns, 2001. Effect of fish meal supplementation on fertility in primiparous lactating beef cows. Proc. West Sec. Am. Soc. Anim. Sci., 52: 74-78.
Direct Link - Braden, T.D., H.R. Sawyer and G.D. Niswender, 1989. Functional and morphological characteristics of the first corpus luteum formed after parturition in ewes. J. Reprod. Fert., 86: 525-533.
PubMed - Buford, W.I., N. Ahmad, F.N. Schrick, R.L. Butcher, P.E. Lewis and E.K. Inskeep, 1996. Embryotoxicity of a regressing corpus luteum in beef cows supplemented with progestogen. Biol. Reprod., 54: 531-537.
CrossRefPubMedDirect Link - Campanile, G., G. Neglia, B. Gasparrini, G. Galiero and A. Prandi et al., 2005. Embryonic mortality in buffaloes synchronized and mated by AI during the seasonal decline in reproductive function. Theriogenology, 63: 2334-2340.
PubMed - Cooper, D.A., D.A. Carver, P. Villeneuve, W.J. Silvia and E.K. Inskeep, 1991. Effects of progestogen treatment on concentration of prostaglandins and oxytocin in plasma from posterior vena cava of post-partum beef cows. J. Reprod. Fert., 91: 411-421.
CrossRefDirect Link - Copelin, J.P., M.F. Smith, H.A. Garverick and R.S. Youngquist, 1987. Effect of the uterus on subnormal at luteal function in anoestrous beef cows. J. Anim. Sci., 64: 1506-1511.
PubMedDirect Link - Cruz, L.C. and D.J. Kesler, 1988. Relationship of GnRH induced LH and FSH profiles on the ovulation response and corpus luteum function of postpartum suckled beef cows. Proceedings of the 11th International Congress on Animal Reproduction and Artifical Insemination, June 26-30, A.J. Dublin, Ireland, pp: 18-18.
- Danet-Denosyers, G., C. Wetzels and W.W. Thatcher, 1994. Natural and recombinant bovine interferon-t regulates basal and oxytocin-induced secretion of prostaglandin-F2-alpha and E2 by epithelial cells and stromal cells in the endomertrium. Reprod. Fert. Dev., 6: 193-202.
PubMed - Demmers, K.J., K. Derecka and A. Flint, 2001. Trophoblast interferon and pregnancy. Reproduction, 121: 41-49.
CrossRef - Dobson, H., M.G.S. Alam and L.N. Kanchev, 1987. Effect of betamethasone treatment on luteal lifespan and the LH response to GnRH in dairy cows. J. Reprod. Fert., 80: 25-30.
Direct Link - Drew, S.B. and A.R. Peters, 1994. Effect of buserelin on pregnancy rates in dairy cows. Vet. Rec., 134: 267-269.
CrossRefDirect Link - Dunne, L.D., M.G. Diskin and J.M. Sreenan, 2000. Embryo and foetal loss in beef heifers between day 14 of gestation and full term. Anim. Reprod. Sci., 58: 39-44.
Direct Link - Farin, C.E., C.L. Moeller, H. Mayan, F. Gamboni, H.R. Sawyer and G.D. Niswender, 1988. Effect of luteinizing hormone and human chorionic gonadotropin on cell populations in the bovine corpus luteurn. Biol. Reprod., 38: 413-421.
CrossRefDirect Link - Fitz, T.A., M.H. Mayan, H.R. Sawyer and G.D. Niswender, 1982. Characterization of two steriodogenic cell types in the bovine corpus luteum. Biol. Reprod., 27: 703-711.
Direct Link - Fortune, J.E. and S.M. Quirk, 1988. Regulation of steroidogenesis in bovine prevoulatory folicies. 18th Biennial Symp. Anim. Reprod. J. Anim. Sci. Suppl., 66: 1-8.
Direct Link - Garcia-Winder, M., P.E. Lewis, E.C. Townsend and E.K. Inskeep, 1987. Effects of norgestomet on follicular development in postpartum beef cows. J. Anim. Sci., 64: 1099-1109.
Direct Link - Garverick, H.A., J.R. Parfet, C.N. Lee, J.P. Copelin, R.S. Youngquist and M.F. Smith, 1988. Relationship of pre and post-ovulatory gonadotropin concentrations to subnormal luteal function in postpartum beef cattle. J. Anim. Sci., 66: 104-111.
PubMedDirect Link - Hansela, W., R.W. Spaldinga, L.L. Larson, D.B. Laster, J.F. Wagner and R.K. Braun, 1976. Influence of human chorionic gonadotropin on pregnancy rates in lactating dairy and beef cows. J. Dairy Sci., 59: 751-754.
CrossRef - Hansel, W. and J.P. Dowd, 1986. New concepts of the control of corpus luteum function. J. Reprod. Fert., 78: 755-768.
PubMedDirect Link - Harrison, L.M., N. Kenny and G.D. Niswender, 1987. Progesterone production, L.H. receptors and oxytocin secretion by ovine luteal cell types on day 6, 10 and 15 of the oestrous cycle and day 25 of pregnancy. J. Reprod. Fert., 79: 539-548.
CrossRef - Hernandez-Ledezma, J.J., J.D. Sikes, C.N. Murphy, A.J. Watson, G.A. Schultz and R.M. Roberts, 1992. Expression of bovine trophoblast interferon in conceptuses derived in vitro techniques. Biol. Reprod., 47: 374-380.
CrossRefDirect Link - Hockett, M.E., N.R. Rohrbach and F.N. Schrick, 2004. Alterations in embryo development in progestogen-supplemented cows administered prostaglandin F2alpha. Prostaglandins Other Lipid Mediat., 73: 227-236.
PubMed - Hommeida, A., T. Nakao and H. Kubota, 2004. Luteal function and conception in lactating cows and some factors influencing luteal function after first insemination. Theriogenology, 62: 217-225.
CrossRefDirect Link - Hoyer, P.B. and G.D. Niswender, 1985. The regulation of steroidogenesis is different in the two types of bovine luteal cells. Can. J. Physiol. Pharmaciol., 63: 240-248.
PubMed - Humblot, P., 2001. Use of pregnancy specific proteins and progesterone assays to monitor pregnancy and determine the timing, frequencies and sources of embryonic mortality in ruminants. Theriogenology, 56: 1417-1433.
PubMed - Humphrey, W.D., C.C. Kaltenbach, T.G. Dunn, D.R. Koritnik and G.D. Niswender, 1983. Characterization of hormonal patterns in the beef cow during postpartum anoestrus. J. Anim. Sci., 56: 445-453.
Direct Link - Hunter, M.G. and J.A. Southee, 1987. Treatment with progesterone affects follicular steriodogenesis in anoestrus ewes. Anim. Reprod. Sci., 14: 273-279.
Direct Link - Hunter, M.G., J.A. Southee and G.E. Lamming, 1988. Function of abnormal corpora lutea in vivo after GnRH-induced ovulation in the anoestrous ewe. J. Reprod. Fert., 84: 131-137.
CrossRef - Imakawa, K., S.D. Helmer, K.P. Nephew, C.S.R. Meka and R.K. Christenson, 1993. A novel role for GM-CSF: Enhancement of pregnancy specific interferon production, ovine trophoblast protein-I. Endocrinology, 132: 1869-1871.
Direct Link - Ing, N.H. and M.B. Tormesi, 1997. Estradiol up-regulate estrogen receptor and progesterone receptor gene expression in specific ovine uterine cells. Biol. Reprod., 56: 1205-1215.
PubMed - Inskeep, E.K., 1995. Factors that affect fertility during oestrous cycles with short or normal luteal phases in post-partum cows. J. Reprod. Fert., 49: 493-503.
PubMed - Kahn, W., 1992. Ultrasonography as a diagnostic tool in female animal reproduction. Anim. Reprod. Sci., 28: 1-10.
Direct Link - Karsch, F.J., D.L. Foster, E.L. Bittman and R.R. Goodman, 1983. A role for estradiol in enhancing hormone pulse frequency during the follicular phase of the estrous cycle of the sheep. Endocrinology, 113: 1333-1339.
PubMed - Kastelic, J.P., D.L. Northey and O.J. Ginther, 1991. Spontaneous embryonic death on days 20 to 40 in heifers. Theriogenology, 35: 351-363.
PubMed - Kieborz-Loos, K.R., H.A. Garverick, D.H. Keisler, S.A. Hamilton, B.E. Salfen, R.S. Youngquist and M.F. Smith, 2003. Oxytocin-induced secretion of prostaglandin F2α in post-partum beef cows: Effects of progesterone and estradiol-17beta treatment. J. Anim. Sci., 81: 1830-1836.
Direct Link - Lamming, G.E., A. Darwash and H.L. Black, 1989. Corpus luteum function in dairy cows and embryo mortality. J. Reprod. Fert., 37: 245-252.
PubMed - Lopez-Gatius, F., P. Santolaria, J. Yaniz, J. Rutllant and M. Lopez-Bejar, 2002. Factors affecting pregnancy loss from gestation day 38 to 90 in lactating dairy cows from a single herd. Theriogenology, 57: 1251-1261.
CrossRefDirect Link - Luck, M.R., J.A. Shale and J.H. Pynae, 1992. Direct stimulation of bovine ovarian progesterone secretion by low concentration of α-interferon. Endocrinology, 135: R5-R8.
PubMed - Macmillan, K.L. and A.J. Peterson, 1993. A new intravaginal progesterone releasing device for cattle (DIDR-B) for estrous synchronization, increasing pregnancy rates and the treatment of postpartum anestrous. Anim. Reprod. Sci., 33: 1-25.
Direct Link - Mann, G.E., G.E. Lamming and M.D. Fray, 1995. Plasma estradiol and progesterone during early pregnancy in the cow and the effects of treatment with buserelin. Anim. Reprod. Sci., 37: 121-131.
Direct Link - Mann, G.E. and G.E. Lamming, 1999. The influence of progesterone during early pregnancy in cattle. Reprod. Domes. Anm., 34: 269-274.
Direct Link - Manns, J.G., G.D. Niswender and T. Braden, 1984. FSH receptors in the bovine corpus luteum. Theriogenology, 22: 321-328.
PubMed - Martal, J., N. Chene, S. Camous, L. Huynth and F. Lantier et al., 1997. Recent developments and potentialities for reducing embryo mortality in ruminants: The role of IFN-tau and other cytokines in early pregnancy. Reprod. Fer. Dev., 9: 355-380.
PubMed - Mattos, R., C.R. Staples and W.W. Thatcher, 2000. Effects of dietary fatty acids on reproduction in ruminants. Rev. Reprod., 5: 38-45.
CrossRefDirect Link - Mattos, R., C.R. Staples, J. Williams, A. Amorocho, M.A. McGuire and W.W. Thatcher, 2002. Uterine, ovarian, and production responses of lactating dairy cows to increasing dietary concentrations of menhaden fish meal. J. Dairy Sci., 85: 755-764.
CrossRef - Maurer, R.R. and J.R. Chenault, 1983. Fertilization failure and embryonic mortality in parous and non-parous beef cattle. J. Anim. Sci., 56: 1186-1189.
Direct Link - McCracken, J.A., W. Schramm and W.C. Okulicz, 1984. Hormone receptor control of pulsatile secretion of PGF2α from the ovine uterus during luteolysis and its abrogation in early pregnancy. Anim. Reprod. Sci., 7: 31-55.
Direct Link - McLeod, B.J., W. Haresign and G.E. Lamming, 1982. The induction of ovulation and luteal function in seasonally anoestrous ewes treated with small dose multiple injections of GnRH. J. Reprod. Fert., 65: 215-221.
CrossRef - McNatty, K.P., M. Gibb, M. Dobb, C. Dobson and D.C. Thurely, 1981. Evidence that change in functioning luteinizing hormone secretion regulates the growth of the preovulatory follicle in the ewe. J. Edor., 90: 375-389.
PubMed - McNeilly, A.S., M. Hunter, R.B. Land and H.M. Fraser, 1981. Inadequate corpus luteum function after induction of ovulation in anoestrous ewes by LH-RH or an LH-RH agonist. J. Reprod. Fert., 63: 137-144.
CrossRef - Meyer, M.D., P.J. Hansen, W.W. Thatcher, M. Drost and L. Badinga et al., 1995. Extension of corpus luteum lifespan and reduction of uterine secretion of prostaglandin F2a of cows in response to recombinant interferon-tau. J. Dairy Sci., 78: 1921-1931.
CrossRef - Milvae, R.A. and W. Hansel, 1980. Concurrent uterine venous and ovarian arterial prostaglandin F concentrations in heifers treated with oxytocin. J. Reprod. Fert., 60: 7-15.
PubMed - Morgan, W.F. and I.J. Lean, 1993. Gonadotrophin-releasing hormone treatment in cattle: A meta-analysis of the effects on conception at the time of insemination. Aust. Vet. J., 70: 205-209.
CrossRefDirect Link - Moss, G.F., J.R. Parfet, C. Marvin, R.D. Allrich and M.A. Diekman, 1985. Pituitary concentrations of gonadotropins and receptors for GnRH in suckled beef cows at various intervals after calving. J. Anim. Sci., 60: 285-293.
Direct Link - Nett, T.M., 1987. Function of the hypothalamic-hypophysial axis during the postpartum period in ewes and cows. J. Reprod. Fertil., 34: 201-213.
PubMed - Nishigai, M., H. Kamomae, T. Tanaka and Y. Kaneda, 2002. Improvement of pregnancy rate in Japanese Black cows by administration of hCG to recipients of transferred frozen-thawed embryos. Theriogenology, 58: 1597-1606.
Direct Link - Niswender, G.D., T.L. Davis, R.J. Griffith, R.L. Bogan and K. Monser et al., 2007. Judge, jury and executioner: The auto-regulation of luteal function. Soc. Reprod. Fert., 64: 191-206.
PubMed - Niswender, G.D., R.H. Schwall, T.A. Fitz, C.E. Farin and H.R. Sawyer, 1985. Regulation of luteal function in domestic ruminants: New concepts. Rec. Prog. Horm. Res., 41: 101-151.
PubMed - Odde, K.G., H.S. Ward, G.H. Kiracofe, R.M. McKee and R.J. Kittok, 1980. Short estrous cycles and associated serum progesterone levels in beef cows. Theriogenology, 14: 105-112.
PubMed - O`shea, J.D., 1987. Heterogeneous cell types in the corpus luteum of sheep, goats and cattle. J. Reprod. Fert., 34: 71-85.
PubMed - O`shea, J.D., R.J. Rodgers and P.J. Wright, 1984. Morphometric analysis and function in vivo and in vitro of corpora lutea from ewes treated with LHRH during seasonal anoestrus. J. Reprod. Fert., 72: 75-85.
CrossRef - Peters, A.R., 1984. Effect of exogenous oestradiol-17-B on gonadotropin secretion in postpartum beef cows. J. Reprod. Fert., 72: 473-478.
Direct Link - Price, C.A. and R. Webb, 1989. Ovarian response to hCG treatment during the oestrus cycle in heifer. J. Reprod. Fert., 86: 303-308.
CrossRef - Priedkalns, J., A.F. Weber and R. Zemuanis, 1968. Qualitative and quantitative morphological studies of the cells of the membrane granulose, theca interna and corpus luteum of the bovine ovary. Cell Tissue Res., 85: 501-520.
CrossRefDirect Link - Prvanovic, N., A. Tomaskovic, J. Grizelj, P. Kocila and M. Samardzija, 2009. Monitoring of early pregnancy and early embryonic mortality by ultrasound and determination of pregnancy-associated glycoproteins and progesterone in cows. Vet. Arch., 79: 259-267.
Direct Link - Remirez-Godinez, J.A., G.H. Kiracofe, R.R. Schalles and G.D. Niswender, 1982. Endocrine patterns in the postpartum beef cow associated with weaning: A comparison of the short and subsequent normal cycles. J. Anim. Sci., 55: 153-158.
Direct Link - Richards, J.S., 1980. Maturation of ovarian follicles: Actions and interactions of pituitary and ovarian hormones on follicular cell differentiation. Physicol. Rev., 60: 51-89.
PubMedDirect Link - Roberts, R.M., 1989. Conceptus, interferons and maternal recognition of pregnancy. Biol. Reprod., 40: 449-452.
CrossRefDirect Link - Roberts, A.J., C.F. Chang, A.V. Schally and J.J. Reeves, 1989. Luteinizing hormone concentrations in serum of postpartum beef cows injected with microencapsulated luteinizing hormone releasing hormone. J. Anim. Sci., 67: 2730-2734.
Direct Link - Roberts, R.M., J.C. Cross and D.W. Leaman, 1992. Interferons as hormones of pregnancy. Endocrine Rev., 13: 432-452.
CrossRefDirect Link - Roche, J.F., M.P. Bolandl and T.A. McGeady, 1981. Reproductive wastage following artificial insemination of heifers. Vet. Rec., 109: 401-404.
PubMed - Rutter, L.M. and R.D. Randel, 1984. Luteal competency during resumption of ovarian cyclicity in postpartum Brahman cows. Theriogenology, 21: 713-725.
PubMed - Rutter, L.M., T.D. Carruthers and J.G. Manns, 1985. The postpartum induced corpus luteum: Functional differences from that of cycling cows and the effects of progesterone pretreatment. Biol. Reprod., 33: 560-568.
CrossRef - Santos, J.E.P., W.W. Thatcher, L. Pool and M.W. Overton, 2001. Effect of human chorionic gonadotropin on luteal function and reproductive performance of high-producing lactating Holstein dairy cows. J. Anim. Sci., 79: 2881-2894.
Direct Link - Savio, J.D., M.P. Boland, N. Hynes and J.F. Roche, 1990. Resumption of follicular activity in the early in post-partum perioed in dairy cows. J. Reprod. Fert., 88: 569-579.
Direct Link - Schams, D.T., A.M. Kruip and R. Koll, 1985. Oxytocin determination in steroid producing tissues and in vitro production in ovarian follicles. Acta. Endocr. Copenh., 109: 530-536.
CrossRef - Schrick, F.N., E.K. Inskeep and R.L. Butcher, 1993. Pregnancy rates for embryos transferred from early postpartum beef cows into recipients with normal estrous cycles. Biol. Reprod., 49: 617-621.
Direct Link - Sheffel, C.E., B.R. Pratt, W.L. Ferrell and E.K. Inskeep, 1982. Induced corpora lutea in the postpartum beef cow. II. Effects of t reatment with progestogen and gonadotropins. J. Anim. Sci., 54: 830-836.
PubMed - Shirar, A., C. Meusnier, J. Paly, M.C. Levasseur and J. Martinet, 1989. Resumption of ovarian activity in post-partum ewes: Role of the uterus. Anim. Reprod. Sci., 19: 79-89.
Direct Link - Shelton, K., M.F.G. De Abreu, M.G. Hunter, T.J. Parkinson and G.E. Lamming, 1990. Luteal inadequacy during the early luteal phase of sub-fertile cows. J. Reprod. Fertil., 90: 1-10.
Direct Link - Silva, P.J., J.L. Juengel, M.K. Rollyson, G.D. Niswender, 2000. Prostaglandin metabolism in the ovine corpus luteum: Catabolism of prostaglandin F2α (PGF2α) coincides with resistance of the corpus luteum to PGF2α. Biol. Reprod., 63: 1229-1236.
CrossRefDirect Link - Sousa, N.M., J.F. Beckers and Z. Gajewski, 2008. Current trends in follow-up of trophoblastic function in ruminant species. Physiol. Pharmacol., 59: 65-74.
PubMedDirect Link - Spencer, T.E. and F.W. Bazer, 2004. Conceptus signals for establishment and maintenance of pregnancy. Reprod. Biol. Endoc., 5: 49-49.
PubMed - Thatcher, W.W., A. Guzeloglu, R. Mattos, M. Binelli, T.R., Hansen and J.K. Pru, 2001. Uterine-conceptus interactions and reproductive failure in cattle. Theriogenology, 56: 1435-1450.
Direct Link - Walters, D.L. and E. Schallenberger, 1984. Pulsatile secretion of gonadotrophins, ovarian steroids and ovarian oxytocin during the periovulatory phase of the oestrous cycle in the cow. J. Reprod. Fertil., 71: 503-512.
PubMed - Wathes, D.C. and G.E. Lamming, 1995. The oxytocin receptor, luteolysis and the mantainence of pregnancy. J. Reprod. Fertil. Suppl., 45: 53-67.
PubMed - Wathes, D.C., C.L. Gilbert and V.J. Ayad, 1993. Interactions between oxytocin, the ovaries and reproductive tract in the regulation of fertility in ewe. Ann. N. Y. Acad. Sci., 689: 396-410.
CrossRef - Webb, R., J.G. Gong, A.S. Law and S.M. Rusbridge, 1992. Control of ovarian function in cattle. J. Reprod. Fertil. Suppl., 45: 141-156.
PubMed - Williams, G.L., 1989. Modulation of luteal activity in postpartum beef cows through changes in dietary lipid. J. Anim. Sci., 67: 785-793.
Direct Link - Wright, J.M., G.H. Kiracofe and R.H. Beeman, 1988. Factors associated with shortened estrous cycles after abortion in beef heifers. J. Anim. Sci., 66: 3185-3189.
Direct Link - Zoli, A.P., L.A. Guibault, P. Delahaut, W.B. Ortiz and J.F. Beckers, 1992. Radioimmunoassay of a bovine pregnancy associated glycoprotein in serum: its aplication for pregnancy diagnosis. Biol. Reprod., 46: 83-92.
PubMed - Zoller, W.G., H.A. Garverick, M.F. Smith, R.J. Moffat, B.E.S. Fen and R.S. Youngquist, 1993. Concentrations of progesterone and oxytocin receptors in endometrial of postpartum cows expected to have a short or normal oestrous cycle. J. Reprod. Fert., 97: 329-337.
CrossRefDirect Link