
J.S. Norrizah
Faculty of Applied Science, Universiti Teknologi MARA, 40450, Shah Alam, Selangor, Malaysia
M. Yaseer Suhaimi
Faculty of Applied Science, Universiti Teknologi MARA, 40450, Shah Alam, Selangor, Malaysia
A. Rohaya
Faculty of Applied Science, Universiti Teknologi MARA, 40450, Shah Alam, Selangor, Malaysia
N.A.R. Nik Roslan
Faculty of Applied Science, Universiti Teknologi MARA, 40450, Shah Alam, Selangor, Malaysia
Biotechnology
Year: 2012 | Volume: 11 | Issue: 4 | Page No.: 238-242







ABSTRACT
Hedyotis corymbosa is a weedy herb belongs to Rubiaceae family. It has many medicinal properties. This plant is well known to contain bioactive medicinal compounds: Ursolic Acid (UA) and Oleanolic Acid (OA). There are vast studies that have been conducted to extract bioactive triterpenes such as ursolic and oleanolic acid from the natural plant of Hedyotis corymbosa. The objective of this study was to analyze the effect of different concentrations of methyl jasmonate on biomass and triterpenes (UA and OA) production in cell suspension culture of H. corymbosa. Cell suspension of Hedyotis corymbosa cultures were successfully induced using 2,4-D plant hormone and then were elicited with methyl jasmonate at different concentrations. The addition of methyl jasmonate elicitor at different concentrations (10, 50, 100 and 200 μM) cause a reduction in the growth of H. corymbosa cell suspension. It was found that cell suspension biomass decreased 52% in the presence of 10 μM methyl jasmonate concentration after 12 days of culture. However, both ursolic acid (ursa-9(11), 12-dien-3-ol acid) and oleanolic acid (olean-12-en-28-oic acid) productions were significantly increased in the presence of 200 and 100 μM methyl jasmonate, respectively, compared to control. In conclusion, the application of different concentrations of methyl jasmonate did not show any significant increase to cell suspension growth but were able to enhance the triterpenes (UA and OA) productions compared to the control. The highest UA (8.4 mg) and OA (17 mg) yields were obtained at 200 and 100 μM methyl jasmonate, respectively, on the third weeks of culture.
PDF Abstract XML References Citation
Received: December 07, 2011;
Accepted: August 11, 2012;
Published: October 09, 2012
How to cite this article
J.S. Norrizah, M. Yaseer Suhaimi, A. Rohaya and N.A.R. Nik Roslan, 2012. Ursolic Acid and Oleanolic Acid Productions in Elicited Cell Suspension Cultures
of Hedyotis corymbosa. Biotechnology, 11: 238-242.
DOI: 10.3923/biotech.2012.238.242
URL: https://scialert.net/abstract/?doi=biotech.2012.238.242
DOI: 10.3923/biotech.2012.238.242
URL: https://scialert.net/abstract/?doi=biotech.2012.238.242
INTRODUCTION
Hedyotis corymbosa belongs to the family Rubiaceae, commonly known in Malaysia as “rumput mutiara”. H. corymbosa exhibited potent hepatoprotective effects and there have been many studies to develop new drugs for hepatitis from this plant (Sadasivan et al., 2006). Many plant species from genus Hedyotis have long been recognized as medicinal herbs in the traditional art of healing and their derivatives continue to be important components of contemporary phytopharmaceuticals. H. corymbosa has been studied for its triterpenes constituents (Kim et al., 2001; Lu et al., 2000) in natural plant extraction. H. corymbosa has also been reported to contain Ursolic Acid (UA) and Oleanolic Acid (OA) (Khastgir et al., 1960). These two bioactive triterpenes: UA and OA have many high medicinal properties. UA and OA were reported to have anti-inflammatory effects, protecting liver, relieving pain and anti-tumor action by enhancing the production of nitric oxide and tumor necrosis factor-α (Qi et al., 2006; Ramachandran et al., 2008; Shehab et al., 2011). More pharmacology effect of UA is being found gradually such as restraining ulcer, controlling blood fat and treating diabetes (Jamil et al., 2007). Meanwhile, OA had been reported to increase the antioxidant component in the liver and maintain hepatic glutathione, this plays important role in protecting against liver injury (Liu, 1995).
Plant tissue culture techniques may provide continuous and reliable source of natural product and an alternative to intact plant tissues for the production of high valuable secondary metabolites such as UA and OA. These days biosynthesis of secondary metabolites from plant tissue cultures is very important considering its medicinal values (Sarin, 2005). Thus, due to the importance of UA and OA, production of these triterpenes using cell suspension cultures is of great practical value. One of the most effective strategies to improve the production of secondary metabolites is using elicitation. Elicitors are substances that act as defense stimulator or stress-induced responses in plants. The elicitor such as methyl jasmonate is widely distributed as regulator signal molecules derived from linolenic acid in plants. Jasmonate act on growth, senescence and as a predominant signal in wound-induced defiance (Binns et al., 2001). Jasmonate are lipid based compounds that exerts numerous effects on plant growth and development (Biondi et al., 2002; Aftab et al., 2011). Jasmonic acid and its methyl ester (methyl jasmonate) have been reported to play an important role in a signal transduction process that regulates defense genes in plants (Farmer and Ryan, 1990; Tachinbana et al., 2007; Sharma et al., 2011). The objective of this study was to analyze the effect of different concentrations of methyl jasmonate on biomass and triterpenes (UA and OA) production in cell suspension culture of H. corymbosa.
MATERIALS AND METHODS
This research was conducted in Plant Tissue Culture and Natural Product Laboratories from 2009 to December 2010.
Cell suspension induction: Suspension culture was induced by transferring 2.0 g of friable and compact callus into 250 mL Erlenmeyer flasks containing 30 mL of liquid MS (Murashige and Skoog, 1962) medium supplemented with 30 g sucrose, 0.1 g myo-inositol and 0.5 mg 2,4-D plant hormone. The cultures were placed on a gyratory shaker at 120 rpm. Subculturing was performed on a 14 days cycle, inoculating approximately 2.0 g of callus to 30 mL of fresh medium. Larger agglomerates from the friable callus culture were separated from the suspension under sterile conditions through a conical metal sieve (diameter of holes: 850 μm) during the subculturing procedure. H. corymbosa cells were separated from the medium by vacuum filtration. The cells were weighed every 3 days until 4 weeks.
Preparation of elicitor: Methyl jasmonate was dissolved in 70% (w/v) ethanol and was prepared as a stock solution. Further dilutions were done using distilled water. Solution was filtered through a microfilter (0.2 μm) before being dispensed into cell cultures at various concentrations (10, 50, 100 and 200 μM).
Extraction process: The cell suspensions were air dried at room temperature for 48 h. The cell suspensions were soaked in the beaker with 250 mL of 80 % methanol. The beaker was covered with aluminum foil to prevent the methanol from evaporating. After 48 h the methanol was filtered and concentrated in vacuum using a rotor vapor at 40°C to get the methanol extract. The procedure was repeated three times to obtain concentrated extracts.
Gas chromatography-mass spectrometry (GC-MS) analysis: The analysis was done according to Razborsek et al. (2008) with some modification. The dried extract was dissolved in 1.0 mL analytical grade methanol. The solution was filtered through a 0.45 μm filter and the clear filtrate was used for GCMS analysis to quantify ursolic acid and oleanolic acid. The separation was carried out using an Agilent Technologies Inc., USA model 6890 GC, equipped with a HP-5MS capillary column; the dimension of the column was 30 mx0.25 mm I.D., 0.25 μm film thickness. The carrier gas was helium at a flow rate of 0.9 mL min-1 with a constant linear velocity of 35 cm sec-1. The oven temperature program was: Initial 105°C (min-0.8), then 12°C min-1 to 200°C (0.1 min), 7°C min-1 to 290°C (6 min), 25°C min-1 to 320°C (10 min). Injector temperature was set at 290°C. Samples were injected in splitless mode. The injection volume was 1 μL. MS detection was carried on an HP quadrupole mass selective detector model 5973. MS was operated in the positive ion mode, with an ionization voltage of 70 eV. The MS data were obtained in full scan mode (mass range 50-750 amu).
Statistical analysis: The data presented is an average of at least 6 replicates from 3 individual experiments. The results in this experiment are quantified and analyzed by using two-ways analysis of variance (ANOVA) from SAS version 9.0 (SAS Institute Inc., USA). All data was expressed as Mean±SEM (standard error mean) and statistical significance is accepted at p<0.05.
RESULTS AND DISCUSSION
Effect of different concentration of methyl jasmonate on biomass production: Study on biomass profile based on dry weight in suspension culture revealed that highest yield was obtained in twelve days of culture period (Fig. 1). From the first day to twelve days, suspension cells were found in their exponential phase. Beginning the eighteen days, the culture exhibited the death phase and the weight decreased rapidly. Meanwhile, detection and quantification showed that both UA and OA were present in suspension culture (Fig. 2, 3). None of the methyl jasmonate concentrations tested turned to attain growth higher than the control culture. The biomass gained in 10 μM methyl jasmonate was 36.1±0.12 g at 12 days of culture, which was 52% lower than control. However, among the methyl jasmonate treatments, 10 and 200 μM methyl jasmonate were found the best concentrations for cell suspension growth. The biomass gained in 10 and 200 μM methyl jasmonate were almost the same started from 18 days till 28 days of culture. Cell suspension culture of Salvia officinalis could obtain a maximal growth and biomass when cultivated without any elicitor (Bolta et al., 2000).
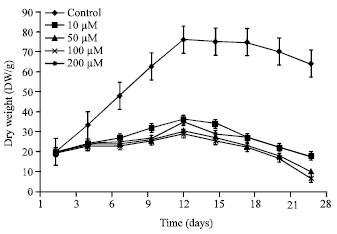 | |
| Fig. 1: | Effects of different concentrations of methyl jasmonate on Hedyotis corymbosa cell suspension culture growth |
Higher concentration of methyl jasmonate suppressed the biomass production of H. corymbosa cell suspension. Meanwhile, a decrease of biomass produced was observed in methyl jasmonate concentration higher than 10 μM. Suppression of growth by methyl jasmonate has also been reported by Yu et al. (2002) who revealed that biomass of Panax ginseng decreased with increasing jasmonic acid concentration. Jasmonic acid elicitation also decreased the biomass of Uncaria tomentosa cell suspension culture (Feria-Romero et al., 2005; Flores-Sanchez et al., 2002).
Cell suspension culture growth characteristics and triterpenes production: The content of the H. corymbosa triterpenes decreased as the culturing periods increased. Results revealed that suspension culture contain high amount of UA compared to the OA (Fig. 2, 3). The highest UA (8.4 mg) and OA (17 mg) yields were obtained at 200 and 100 μM methyl jasmonate, respectively, on the third weeks of culture.
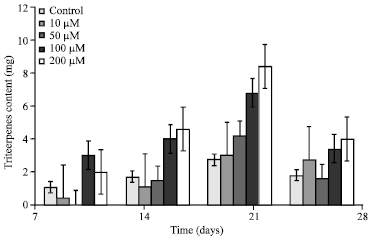 | |
| Fig. 2: | Effects of methyl jasmonate on triterpenes (UA) production in cell suspension cultures of H. corymbosa cultured in MS medium |
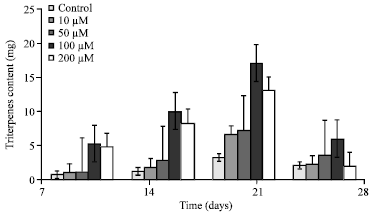 | |
| Fig. 3: | Effects of methyl jasmonate on triterpene (OA) production in cell suspension cultures of H. corymbosa cultured in MS medium |
Methyl jasmonate at 100 and 200 μM appear to be a suitable concentration for UA enhancement in cell suspension cultures of H. corymbosa. Cell suspension supplemented with methyl jasmonate at 10 μM give a lower yield of ursolic acid. Methyl jasmonate are considered to be involved in a part of the signal transduction pathway that induces particular enzymes catalyzing biochemical reaction to form defense compounds of low molecule weight in plants such as polyphenols, alkaloids, quinines, terpenoids and polypeptides (Gundlach et al., 1992; Mueller et al., 1993; Seo et al., 1993; Morimoto et al., 1998). The role of methyl jasmonate in protecting plants from various stresses has been reported for several plants (Nadarajah, 2001; Parthier et al., 1992; Creelman and Mullet, 1997; Norastehnia and Nojavan-Asghari, 2006). Yu et al. (2002) concluded that methyl jasmonate triggered the enzyme activities for the synthesis of triterpenoid saponins, protopanaxadiol ginsenosides in Panax ginseng. In Uncaria tomentosa cell cultures, formation of UA and OA is mediated by jasmonic acid as a secondary messenger in response to elicitation (Feria-Romero et al., 2005). OA content at day 21 was enhanced 78 and 71% in cell suspension contains 100 and 200 μM, respectively more than control. However, all the methyl jasmonate concentrations were able to enhance the production of OA higher than the control. Similar pattern of OA production enhancement after 20 days of cultivation was showed in cell suspension of Calendula officinalis cultivated with MS media supplemented with 2,4-D hormone (Grzelak and Janiszowska, 2002). On the other hand, OA yield was significantly influenced by the addition of methyl jasmonate at any concentrations. Premjet et al. (2002), reported that methyl jasmonate also able to enhance the production of podophyllotoxin in the Juniperus chinensis suspension cultures. Several studies also showed that an increase in sucrose contain in the media can enhance the secondary metabolites production including UA and OA (Feria-Romero et al., 2005; Rothe et al., 2001; Smeekens, 2000). Higher accumulation of UA content was observed in a single cells fraction of Salvia officinalis than in cell aggregates (Bolta et al., 2003). This study came to the same suggestion with Yoon et al. (2000) that, methyl jasmonate is a substance that involve in the downstream step of the triterpenoid induction pathway. The present study showed that methyl jasmonate is suitable to be used as an elicitor to enhanced UA and OA in the H. corymbosa cell suspension culture.
CONCLUSION
In conclusion, the application of different concentrations of methyl jasmonate did not show any significant increased to cell suspension growth of Hedyotis corymbosa but were able to enhance the triterpenes (UA and OA) productions compared to the control. The highest UA (8.4 mg) and OA (17 mg) yields were obtained at 200 and 100 μM methyl jasmonate, respectively, on the third weeks of culture.
ACKNOWLEDGMENTS
The authors would like to thank RMI, University Technology MARA (UiTM) and the Ministry of Higher Education for the grant No. 600-IRDC/ST/FRGS.5/3/1146.
REFERENCES
- Aftab, T., M.M.A. Khan, M. Idrees, M. Naeem, Moinuddin and N. Hashmi, 2011. Methyl jasmonate counteracts boron toxicity by preventing oxidative stress and regulating antioxidant enzyme activities and artemisinin biosynthesis in Artemisia annua L. Protoplasma, 248: 601-612.
CrossRefPubMedDirect Link - Norastehnia, A. and M. Nojavan-Asghari, 2006. Effects of methyl jasmonate on the enzymatic antioxidant defense system in maize seedlings subjected to paraquat. Asian J. Plant Sci., 5: 17-23.
CrossRefDirect Link - Binns, S.E., I. Inparajah, B.R. Baum and J.T. Arnason, 2001. Methyl jasmonate increases reported alkamides and ketoalkene/ynes in Echinacea pallida (Asteraceae). Phytochemistry, 57: 417-420.
CrossRefDirect Link - Biondi, S., S. Scaramagli, K.M. Oksman-Caldentey and F. Poli, 2002. Secondary metabolism in root and callus cultures of Hyoscyamus muticus L.: The relationship between morphological organisation and response to methyl jasmonate. Plant Sci., 163: 563-569.
CrossRefDirect Link - Bolta, I., D. Barievi, B. Bohanec and S. Andrenek, 2000. A preliminary investigation of ursolic acid in cell suspension culture of Salvia officinalis. Plant Cell Tissue Organ Culture., 62: 57-63.
CrossRefDirect Link - Bolta, Z., D. Baricevic and P. Raspor, 2003. Biomass segregation in sage cell suspension culture. Biotechnol. Lett., 25: 61-65.
PubMed - Creelman, R.A. and J.E. Mullet, 1997. Biosynthesis and action of jasmonates in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 48: 355-381.
CrossRefPubMedDirect Link - Premjet, D., K. Itoh and S. Tachibana, 2002. Enhancement of podophyllotoxin production by biogenetic precursors and elicitors in cell suspension cultures of Juniperus chinensis. Pak. J. Biol. Sci., 5: 1267-1271.
CrossRefDirect Link - Feria-Romero, I., E. Lazo, T. Ponce-Noyola, C.M. Cerda-Garcia-Rojas, A.C. Ramos-Valdivia, 2005. Induced accumulation of oleanolic acid and ursolic acid in cell suspension cultures of Uncaria tomentosa. Biotechnol. Lett., 27: 839-843.
CrossRefPubMedDirect Link - Flores-Sanchez, I.J., J. Ortega-lopez, M.C. Montes Horcasitas and A.C. Ramos Valdivia, 2002. Biosynthesis of sterols and triterpenes in cell suspension cultures of Uncaria tomentosa. Plant Cell Physiol., 43: 1502-1509.
CrossRef - Grzelak, A. and W. Janiszowska, 2002. Initiation and growth characteristics of suspension cultures of Calendula officinalis cells. Plant Cell Tissue Organ Culture, 71: 29-40.
CrossRefDirect Link - Khastgir, H., S.K. Sengupta and P. Sengupta, 1960. Note on the constituents of the Indian medicinal plant Oldenlandia corymbosa Linn. J. Am. Pharm. Assoc., 49: 562-563.
CrossRefDirect Link - Kim, Y., E.J. Park, J. Kim, Y.B. Kim, S.R. Kim and Y.C. Kim, 2001. Neuroprotective constituents from Hedyotis diffusa. J. Nat. Prod., 64: 75-78.
CrossRefPubMedDirect Link - Liu, J., 1995. Pharmacology of oleanolic acid and ursolic acid. J. Ethnopharmacol., 49: 57-68.
CrossRefPubMedDirect Link - Lu, C.M., J.J. Yang, P.Y. Wang and C.C. Lin, 2000. A new acylated flavonol glycoside and antioxidant effects of Hedyotis diffusa. Planta Med., 66: 374-377.
CrossRefPubMedDirect Link - Morimoto, S., N. Tateishi, T. Matsuda, H. Tanaka and F. Taura et al., 1998. Novel hydrogen peroxide metabolism in suspension cells of Scutellaria baicalensis Georgi. J. Biol. Chem., 273: 12606-12611.
Direct Link - Mueller, M.J., W. Brodchelm, E. Spannagl and M.H. Zenk, 1993. Signalling in the elicitation process is mediated through the octadecanoid pathway leading to jasmonic acid. Proc. Natl. Acad. Sci., 90: 7490-7494.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Razborsek, M.I., D.B. Voncina, V. Dolecek and E. Voncina, 2008. Determination of oleanolic, betulinic and ursolic acid in Lamiaceae and mass spectral fragmentation of their trimethylsilylated derivatives. Chromatographia, 67: 433-440.
CrossRefDirect Link - Rothe, G., U. Garske and B. Drager, 2001. Calystegines in root cultures of Atropa belladonna respond to sucrose, not to elicitation. Plant Sci., 160: 1043-1053.
CrossRefDirect Link - Nadarajah, K., 2001. The signalling pathway involved in plant-pathogen interactions in Arabidopsis thaliana. Pak. J. Biol. Sci., 4: 508-510.
CrossRefDirect Link - Ramachandran, S., N. Rajendra Prasad, K.V. Pugalendi and V.P. Menon, 2008. Modulation of UVB-induced oxidative stress by ursolic acid in human blood lymphocytes. Asian J. Biochem., 3: 11-18.
CrossRefDirect Link - Sarin, R., 2005. Useful metabolites from plant tissue cultures. Biotechnology, 4: 79-93.
CrossRefDirect Link - Sadasivan, S., P.G. Latha, J.M. Sasikumar, S. Rajashekaran, S. Shyamal and V.J. Shine, 2006. Hepatoprotective studies on Hedyotis corymbosa (L.) Lam. J. Ethnopharmacol., 106: 245-249.
CrossRefPubMedDirect Link - Smeekens, S., 2000. Sugar-induced signal transduction in plants. Ann. Rev. Plant Physiol. Plant Mol. Biol., 51: 49-81.
CrossRef - Tachinbana, S., T. Muranaka and K. Itoh, 2007. Effect of elicitors and a biogenetic precursor on paclitaxel production in cell suspension cultures of Taxus cuspidata Var. nana. Pak. J. Biol. Sci., 10: 2856-2861.
CrossRefPubMedDirect Link - Qi, S., L. Ding, K. Tian, X. Chen and Z. Hu, 2006. Novel and simple nonaqueous capillary electrophoresis separation and determination bioactive triterpenes in Chinese herbs. J. Pharm. Biomed. Anal., 40: 35-41.
CrossRefDirect Link - Sharma, M., A. Sharma, A. Kumar and S.K. Basu, 2011. Enhancement of secondary metabolites in cultured plant cells through stress stimulus. Am. J. Plant Physiol., 6: 50-71.
CrossRefDirect Link - Shehab, N.G., A. Mahdy, S.A. Khan and S.M. Noureddin, 2011. Chemical constituents and biological activities of Fagonia indica burm F. Res. J. Med. Plant., 5: 531-546.
CrossRef - Yoon, H.J., H.K. Kim, C.J. Ma and H. Huh, 2000. Induced accumulation of triterpenoids in Scutellaria baicalensis suspension cultures using a yeast elicitor. Biotechnol. Lett., 22: 1071-1075.
CrossRefDirect Link - Yu, K.W., W.Y. Gao, E.J. Hahn and K.Y. Paek, 2002. Jasmonic acid improves ginsenoside accumulation in adventitious root culture of Panax ginseng C.A. Meyer. Biochem. Eng. J., 11: 211-215.
CrossRefDirect Link