
Shrikant A. Patil
Trivedi Foundation, 14500 N Northsight, Blvd No. 309, Scottsdale, AZ 85260, USA
Gopal B. Nayak
Trivedi Foundation, 14500 N Northsight, Blvd No. 309, Scottsdale, AZ 85260, USA
Siddhivinayak S. Barve
Kelkar Education Trust`s Scientific Research Center, KET`s V.G. Vaze College Campus, Mithagar Road, Mulund (E), Mumbai 400081, India
Rashmi P. Tembe
Kelkar Education Trust`s Scientific Research Center, KET`s V.G. Vaze College Campus, Mithagar Road, Mulund (E), Mumbai 400081, India
Rummana R. Khan
Kelkar Education Trust`s Scientific Research Center, KET`s V.G. Vaze College Campus, Mithagar Road, Mulund (E), Mumbai 400081, India
Biotechnology
Year: 2012 | Volume: 11 | Issue: 3 | Page No.: 154-162







ABSTRACT
Pogostemon cablin is a known aromatic plant which is cultivated for its essential oil widely applicated in perfumery and cosmetic industries. In the present study, the effect of biofield treatment was studied on the growth of P. cablin. For this study an in vitro culture system was set up in two groups, viz., control and treatment, each of which was derived from three different explant sources, namely leaf, node and petiole. Further these in vitro plantlets were hardened and transferred to external environment. The stomatal cells and epidermal hair growth were also studied at various morphogenetic stages. The study revealed that a single spell of biofield energy treatment produced significant increase in growth in treated group throughout all the morphogenetic phases from in vitro to in vivo level. A remarkable increase in stomatal cells and epidermal hair was also seen in treated group.
PDF Abstract XML References Citation
Received: March 01, 2012;
Accepted: June 15, 2012;
Published: July 03, 2012
How to cite this article
Shrikant A. Patil, Gopal B. Nayak, Siddhivinayak S. Barve, Rashmi P. Tembe and Rummana R. Khan, 2012. Impact of Biofield Treatment on Growth and Anatomical Characteristics of Pogostemon cablin (Benth.). Biotechnology, 11: 154-162.
DOI: 10.3923/biotech.2012.154.162
URL: https://scialert.net/abstract/?doi=biotech.2012.154.162
DOI: 10.3923/biotech.2012.154.162
URL: https://scialert.net/abstract/?doi=biotech.2012.154.162
INTRODUCTION
Patchouli (Pogostemon cablin Benth.) is an herbaceous plant native to South and South East Asia (Maheshari et al., 1993). The leaves of the plant yield aromatic oil which is highly valued in perfumery and aromatherapy (Hasegawa et al., 1992). Indonesia is the largest producer of patchouli oil (Department of Biotechnology, 2009). However, this supply of oil is irregular due to the adverse weather conditions. The world consumption of patchouli oil is 1500 tons out of which Indian perfumery market imports over 150 tons of oil annually, as the current production in the country is only around 10 tons. India’s available infrastructure and environment present us the opportunity to gain a major part of the world market for patchouli oil production. To achieve this goal a number of enhancement factors were studied to improve the quality and quantity of patchouli oil to make it readily acceptable for the perfumery industry.
In recent years, many healing modalities have come into picture based on the individual-environment energy interactions. A number of terms were coined for these energy modalities out of which “Biofield” was the term accepted worldwide (Rubik, 2002). Many attempts were made to study the biological effects of these fields on proteins, antibiotics, water, in vitro cells, bacteria, plants and animals (Chein et al., 1991; Schwartz and DeMattei, 1991; Benor, 1990; Rein, 1995; Chen et al., 2002; Mo et al., 2003; Creath and Schwartz, 2004; Baldwin and Schwartz, 2006).
So, far researchers have worked upon the healing effect of these biofields in different medical streams such as hematology, immunology, wound healing ability and other clinical applications (Krieger, 1976; Wirth, 1990; Olson et al., 1997; Movaffaghi et al., 2006). However, until now not much work has been done to study the effect of biofields on the plants in in vitro system. It has been well known that plants are complex multicellular organisms that are highly sensitive to initial assaying of effects and testing of new therapies.
The current research was undertaken to study the effect of biofield treatment on P. cablin in vitro. The research was focused on the effect of energy transfer to P. cablin plants in in vitro system and its effect on growth in different morphogenetic phases.
MATERIALS AND METHODS
Plant material: The current study was carried out from January 2011-January 2012. The in vitro plantlets of P. cablin established in our plant tissue culture laboratory were used as an explant source for the present study. The leaf, node and petiole from these in vitro plantlets were used as explants for clonal propagation. In vitro initiation from these plantlets was done in triplicate at intervals of one week.
Media: Leaf, node and petiole explants obtained were cultured on Murashige and Skoog’s Medium (MS) (Murashige and Skoog, 1962) supplemented with 3% (w/v) sucrose, pH 5.7. The medium was solidified using 0.8% (w/v) agar (Bacteriological grade, Marine Chemicals, Cochin, India). All the chemicals used were from Qualigens and of analytical grade. About 20 and 50 mL media was dispensed in Pyrex glass Borosil®culture tubes (25x150 mm) and culture bottles, respectively. The tubes and bottles were covered with tight fitting plastic caps. Both the culture tubes and bottles were autoclaved at 1.05 kg cm-2 for 15 min. The cultures were maintained at 25±2°C under 16 h photoperiod provided by the cool-white fluorescent tubes.
Biofield treatment modality: The biofield treatment was given to the cultures via Single Treatment method (ST). In ST the cultures were subjected to biofield treatment only once at the initiation phase. In addition to this, respective sets of control cultures that were devoid of any treatment were also maintained. Thus, two groups of cultures obtained from leaf, node and petiole explant viz., control and treated, were maintained for further study.
Establishment of in vitro cultures
Shoot initiation: Leaf, node and petiole explants obtained from in vitro plantlets were inoculated on full strength basal MS medium in glass tubes. After one week in initiation medium the treated group of cultures was subjected to a spell of treatment via ST method. The cultures were maintained on shoot initiation medium for a month. After a month they were transferred to shoot multiplication medium. At the end of the month the total number of shoots produced per explant in each set was scored and the data was analyzed by paired t-test.
Shoot multiplication: The initiated shoots from shoot initiation medium were transferred to shoot multiplication medium that is full strength basal MS medium in glass bottles. The cultures were maintained on shoot multiplication medium for five months and were subcultured after every month. The total biomass of cultures and multiplication rate was scored at the end of each month. The data was analyzed by paired t-test.
Plantlet regeneration: After about five months in shoot multiplication medium the regenerated shoots of about 2 cm in height were transferred to rooting medium which is half strength basal MS medium in glass bottles. The cultures were maintained on rooting medium for two months and were subcultured after every month. At the end of rooting phase the parameters studied were, rooting response, days required for root initiation, regeneration rate, multiplication rate, number of roots per shoot, length of root, length of shoot, number of nodes, internodal distance and total number of plantlets regenerated at the end of rooting phase. The data was analyzed by paired t-test.
Hardening of rooted plantlets: The rooted plantlets of the control and treated groups were washed under running tap water to remove traces of agar. They were treated with 1% (w/v) systemic fungicide Bavistin® for 1 min. Then they were potted in potting mixture containing soil and compost (2:1), thoroughly drenched with water. The potted plants were covered with polycaps to prevent desiccation and were kept in the green house where the humidity was 70-80% and the temperature was 26±2°C. After about ten days the polycaps were removed and the plants were sprayed with water. After another ten days the plants were repotted in bigger polythene bags containing a mixture of soil, sand and compost (3:2:1). The plants were maintained in these polybags for 20 days in the green house. They were watered and sprayed with fungicide Bavistin® and fertilizer Biozyme® alternately after every 4 days. Then the plants were transferred to the shade house. Here, they were maintained for 15 days under similar conditions, after which they were transferred to open sunlight.
Microscopic study of stomata and epidermal hairs: For this study plants were selected from 6 different morphogenetic stages, namely a month old in vitro rooted plantlet, primary hardened plants with polycaps, primary hardened plants without polycaps, secondary hardened plants, plants kept in the shade house and plants kept in sunlight. For studying stomata, the lower epidermis was taken into consideration whereas for epidermal hair study the upper epidermis was used. The slides for the study were prepared using finger nail polish imprints as follows. A small area of the leaf lamina about 1 cm2 was selected. This area was then painted with transparent finger nail polish and was allowed to dry completely for 5 min. After this a clear cellophane tape about 3 cm in length was pressed on the nail polish imprint. Then it was gently pulled off and the imprint was carefully mounted on a clean and grease free microscopic slide. The excess cellophane tape was removed with the help of a sharp blade. The slides were observed under a compound microscope initially under low magnification (10x) and then under high magnification (40x) for further study. For calculation of Stomatal index three random microscopic fields were selected under high magnification (40x). The epidermal cells and stomatal cells in each of these fields were counted and the Stomatal index was calculated using the following formula:
The epidermal hairs were studied using the photographs taken by CCD camera (Watec Digital, WAT-202D, W73-27235, Aegis Electronic Group Inc., Arizona, US) attached to the microscope (MKOW OPTIK, Scientific India, Ambala Cantt., Haryana, India) under high magnification (40x).
RESULTS
Shoot initiation: Three different explants were used for in vitro studies in P. cablin, namely leaf, node and petiole. Shoot initiation was seen in all the three explants in both control and treated groups after six days of initiation (Fig. 1, 2a-c). However, at the end of initiation phase, the mean number of shoots in ST was comparable to its respective control (Table 1). In treated cultures the mean number of shoots obtained from leaf explant was 7.97 which was close to its control where the mean number was 7.08. Similarly in nodal explant the mean number of shoots in control and treated cultures was 10.85 and 9.57, respectively. On the contrary, in case of petiole explant, the mean number of shoots in treated was less that is 7.9 as compared to 8.73 in its respective control.
Shoot multiplication: Multiple shoot proliferation was studied over a period of five months. At the end of the first month, treatment showed increase in biomass comparable to the control (Fig. 1d-f). Later at the end of the second month, the treatment showed a significant increase in biomass in cultures derived from leaf and petiole explants. The increase in biomass in treated cultures was 2.23 and 2.4 g compared to 2.12 and 2.08 g in control cultures. However, at the end of the third month the control group showed significant increase in biomass in cultures derived from leaf, node and petiole explants over the treated group.
| Table 1: | Data collected at the end of shoot initiation phase in Pogostemon cablin |
 | |
| Data recorded after a month in shoot initiation medium (n = 30) | |
Further at the end of the fourth month ST was seen effective for all the three explants, leaf, node and petiole, with an increase in biomass which was 3.06, 3.12 and 3.04 g, respectively (Table 2) (Fig. 2d-f). Similar results were obtained at the end of the fifth month where ST was seen effective in all the three explants with an increase in biomass up to 3.17, 3.07 and 3.17 g compared to its control. Thus, it was observed that in the entire multiple shoot proliferation phase the cultures from all the three explants showed increase in biomass in the treated group.
Plantlet regeneration: Plantlet regeneration was studied over a period of two months. Both control and treated cultures showed 100% rooting (Fig. 1g-i). It was seen that treated cultures required less days for root initiation as compared to their respective control. The number of days required for rooting in treated cultures was 8 days whereas control cultures showed root initiation at the end of 10 days (Table 3). The plantlet multiplication rate in both control and treated cultures was found to be identical. However, the plantlet regeneration rate in cultures treated with ST was significantly high (Fig. 2g-i). It was as high as 2.76, 4.58 and 1.41 in treated group when compared with 1.0 in control group for all the three explants. On an average the treatment shows 3 fold increase in total number of plantlets regenerated when compared to its respective control (Table 3). Further, treated cultures showed significant increase in mean number of roots produced per shoot which was 17 as compared to 11 in control cultures (Table 4). The length of roots, length of shoots, total number of nodes, internodal distance and length of entire plant in treated group were comparable to the control group (Table 4).
Hardening of rooted plantlets: A mixture of soil and sand (2:1) was used for initial acclimatization of plants. The plants survived and acclimatized well in the above potting mixture. The survival percentage was 98, 95 and 90% for plants derived from leaf, node and petiole explant, respectively in both treated as well as control groups. Later the plants were maintained in sand, soil and compost (3:2:1) for further growth with 100% survival rate.
 | |
| Fig. 1(a-i): | Various morphogenetic stages of P. cablin in vitro in control group, (a) Shoot initiation from leaf explant (bar 1.67 cm), (b) Shoot initiation from node explant (bar 1.84 cm), (c) Shoot initiation from petiole explant (bar 2 cm), (d) Shoot multiplication from leaf explant (bar 1.11 cm), (e) Shoot multiplication from node explant (bar 1.14 cm), (f) Shoot multiplication from petiole explant (bar 1.09 cm), (g) Plantlet regeneration from leaf explant (bar 1.09 cm), (h) Plantlet regeneration from node explant (bar 1.08 cm) and (i) Plantlet regeneration from petiole explant (bar 1.04 cm) |
| Table 2: | Data collected throughout the shoot multiplication phase in Pogostemon cablin |
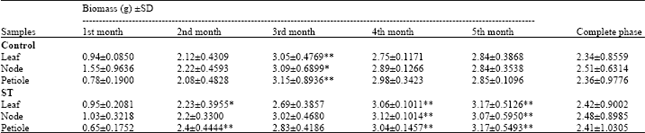 | |
| Data recorded after 1, 2, 3, 4 and 5 months in shoot multiplication medium (n = 30) *Significant and **Highly significant at p<0.05 when compared to control | |
On transferring to the shade house both the groups of plants showed vigorous growth. However, the plants from treated group showed more number of nodes which resulted in increase in height as compared to control group (Fig. 3a, c and e). Also the treated plants appeared healthy and the leaves were mature and deeply veined as compared to control plants (Fig. 3b, d and f). At the end of 3 months they were ready for field cultivation.
 | |
| Fig. 2(a-i): | Various morphogenetic stages of P. cablin in vitro in treated group, (a) Shoot initiation from leaf explant (bar 1.72 cm), (b) Shoot initiation from node explant (bar 1.7 cm), (c) Shoot initiation from petiole explant (bar 1.6 cm), (d) Shoot multiplication from leaf explant (bar 1.14 cm), (e) Shoot multiplication from node explant (bar 1.1 cm), (f) Shoot multiplication from petiole explant (bar 1.11 cm), (g) Plantlet regeneration from leaf explant (bar 1.14 cm), (h) Plantlet regeneration from node explant (bar 1.11 cm) and (i) Plantlet regeneration from petiole explant (bar 1.19 cm) |
| Table 3: | Data (non-statistical parameters) collected throughout the plantlet regeneration phase in Pogostemon cablin |
 | |
Microscopic study of stomatal index and epidermal hairs: It was observed that the SI in both control and treated plants showed a decrease when the plants were transferred from in vitro to in vivo phases. It decreased from 0.2970, 0.2932 and 0.2698 to 0.1913, 0.2099 and 0.2336 in plants derived from leaf, node and petiole explants respectively in control group (Table 5). A similar decrease in SI was seen from 0.3312, 0.3054 and 0.3088 to 0.2711, 0.2834 and 0.2862 in plants derived from leaf, node and petiole, respectively in treated group (Table 5).
 | |
| Fig. 3(a-f): | Three months old hardened plants of P. cablin, (a) Plants derived from leaf explant in control group (bar 1.12 cm), (b) Plants derived from leaf explant in treated group (bar 1.04 cm), (c) Plants derived from nodal explant in control group (bar 1.28 cm), (d) Plants derived from nodal explant in treated group (bar 1.05 cm), (e) Plants derived from petiole explant in control group (bar 1.29 cm), (f) Plants derived from petiole explant in treated group (bar 1.61 cm) |
| Table 4: | Data (statistical parameters) collected throughout the plantlet regeneration phase in Pogostemon cablin |
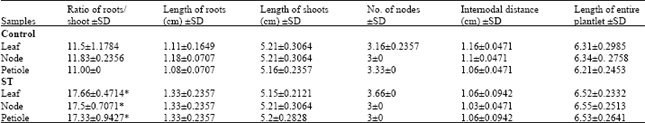 | |
| *Significant at p<0.05 when compared to control | |
Later with the progressive hardening process the SI showed a gradual increase in both control and treated groups. However, the SI in treated plants was high compared to the respective control plants (Fig. 4a, b). It was as high as 0.4330, 0.3314 and 0.3409 when compared to 0.3328, 0.3002 and 0.2476 in plants derived from leaf, node and petiole explants in treated and control groups, respectively (Table 5). Similarly, the epidermal hair study revealed that, in the initial phase of morphogenesis, the number of glandular hairs is very less; however, a gradual change from in vitro to in vivo phase results in increased number of glandular hairs.
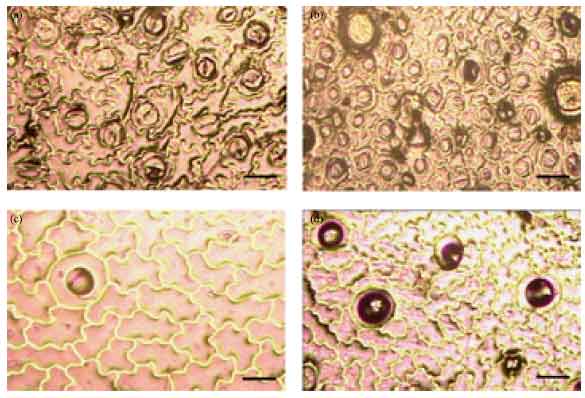 | |
| Fig. 4(a-d): | Study of stomata and glandular hair in P. cablin as seen under microscope with 40x magnification, (a) Stomata seen on the lower epidermis of leaf in control plants (bar 49.2 μm), (b) Stomata seen on the lower epidermis of leaf in treated plants (bar 50.6 μm), (c) Glandular hair seen on the upper epidermis of leaf in control plants (bar 49.5 μm), (d) Glandular hair seen on the upper epidermis of leaf in treated plants (bar 50.3 μm) |
| Table 5: | Stomatal index of plants of Pogostemon cablin in different morphogenetic phase |
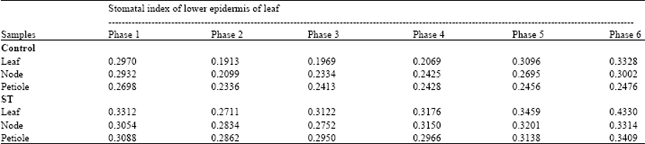 | |
| Phase 1: A month old in vitro rooted plantlet, Phase 2: Primary hardened plants with polycaps, Phase 3: Primary hardened plants without polycaps, Phase 4: Secondary hardened plants, Phase 5: Plants kept in shade house, Phase 6: Plants kept in sunlight | |
When control and treated groups were compared, it was observed that the upper epidermis of leaves of treated plants showed more glandular hairs (Fig. 4c, d). The increase in number of glandular hairs is evidently seen from the photographs taken.
DISCUSSION
The biofield energy transferred to in vitro cultures of P. cablin was observed to show significant effect on plant growth at different morphogenetic stages. In the initiation phase ST was found to be comparable to control. Though in multiple shoot proliferation phase the treatment showed varied effect on cultures derived from all the three explants in terms of biomass yield, a significant effect was seen in plantlet regeneration phase where total number of plantlets regenerated in treated group was nearly thrice than that of control group. There were certain changes which were seen in plants treated with ST. It was observed that morphologically treated plants were healthier as compared to control. Similarly 100% response, less days required for regeneration, increase in root and shoot number and increase in biomass were some of the evidently seen effects in treated cultures which were significant when compared with their respective control cultures.
Microscopic study of stomata and epidermal hairs further supported the superiority of treated plants over control. From in vitro to in vivo phases, a gradual decrease in light intensity results in decrease in stomatal index in all plants. Further as the light intensity increases during progressive hardening phases there is a gradual increase in stomatal index in all plants. Thus, stomatal index is found to be directly correlated to light intensity in surrounding conditions. This is in correlation to previous studies which showed that increase in light intensity results in increase in stomatal index (Lake et al., 2001). Study of epidermal hairs showed similar increases in treated plants in later phases of hardening. Also this number was high when compared to control plants. Increase in glandular hairs would directly contribute to increase in oil yield in treated plants. This increase in essential oil content as a result of biofield treatment will add to the economic value of the P. cablin plants which can be further subjected to large scale cultivation.
Till date, very few findings are available in which work was conducted to study the effect of biofields on living systems in vitro. Researchers have worked upon the effect of Reiki treatment on bacterial cultures with respect to its healing context (Rubik et al., 2006). Also the effect of music, noise and sound has been studied in seed germination assays (Creath and Schwartz, 2004). However, this is for the first time that attempts were made to study the effect of biofield treatment on the entire regeneration phase of a plant in in vitro system.
It was observed that a single spell of energy transfer in treated group is effective in enhancing the growth to a significantly higher rate as compared to the control group in P. cablin plants. However, these observations are in respect to the effect of biofield treatment at various morphogenetic stages at in vitro and acclimatization phases. Further these regenerated plantlets will be subjected to screening methods at morphological, phytochemical and molecular levels. These further studies may reveal a deeper impact of the biofield energy on plant systems at various levels.
REFERENCES
- Baldwin, A.L. and G.E. Schwartz, 2006. Personal interaction with a Reiki practitioner decreases noise-induced microvascular damage in an animal model. J. Altern. Complementary Med., 12: 15-22.
CrossRefPubMedDirect Link - Chein, C.H., J.J. Tsuei, S.C. Lee, Y.C. Huang and Y.H. Wei, 1991. Effect of emitted bioenergy on biochemical functions of cells. Am. J. Chi. Med., 19: 285-292.
CrossRefPubMedDirect Link - Chen, K.W., S.C. Shiflett, N.M. Ponzio, B. He, D.K. Elliott and S.E. Keller, 2002. A preliminary study of the effect of external qigong on lymphoma growth in mice. J. Altern. Complementary Med., 8: 615-621.
CrossRefPubMedDirect Link - Creath, K. and G.E. Schwartz, 2004. Measuring effects of music, noise and healing energy using a seed germination bioassay. J. Altern. Complementary Med., 10: 113-122.
CrossRefPubMedDirect Link - Hasegawa, Y., K. Tajima, N. Toi and Y. Sugimura, 1992. An additional constituent occurring in the oil from a patchouli cultivar. Flavor Fragrance J., 7: 333-335.
CrossRefDirect Link - Krieger, D., 1976. Healing by the laying-on of hands as a facilitator of bioenergetic change: The response of In-vivo human hemoglobin. Psychoenergetic Syst., 1: 121-129.
Direct Link - Lake, J.A., W.P. Quick, D.J. Beerling and F.I. Woodward, 2001. Plant development: Signals from mature to new leaves. Nature, 411: 154-154.
CrossRef - Mo, Z., K.W. Chen, W. Ou and M. Li, 2003. Benefits of external qigong therapy on morphine-abstinent mice and rats. J. Altern. Complementary Med., 9: 827-835.
CrossRefPubMedDirect Link - Movaffaghi, Z., M. Hasanpoor, M. Farsi, P. Hooshmand and F. Abrishami, 2006. Effects of therapeutic touch on blood hemoglobin and hematocrit level. J. Holistic Nursing, 24: 41-48.
CrossRefPubMedDirect Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Olson, M., N. Sneed, M. LaVia, G. Virella, R. Bonadonna and Y. Michel, 1997. Stress-induced immunosuppression and therapeutic touch. Altern. Ther. Health Med., 3: 68-74.
PubMedDirect Link - Rein, G., 1995. The In vitro effect of bioenergy on the conformational states of human DNA in aqueous solutions. Acupunct. Electrother. Res., 20: 173-180.
PubMedDirect Link - Rubik, B., 2002. The biofield hypothesis: Its biophysical basis and role in medicine. J. Altern. Complementary Med., 8: 703-717.
CrossRefPubMedDirect Link - Rubik, B., A.J. Brooks and G.E. Schwartz, 2006. In vitro effect of Reiki treatment on bacterial cultures: Role of experimental context and practitioner well-being. J. Altern. Complementary Med., 12: 7-13.
CrossRefPubMedDirect Link