
M. M.M. Raja
Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020, Tamilnadu, India
A. Raja
Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020, Tamilnadu, India
M. M. Imran
Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020, Tamilnadu, India
A. M.I. Santha
Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020, Tamilnadu, India
K. Devasena
Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020, Tamilnadu, India







ABSTRACT
Enzyme metabolism is a fundamental biological process that is vital for the survival of all species. Their specific function is to catalyze chemical reactions. Enzymes have found wide and diverse applications at which enzymes increase the rate of reactions which approach to equilibrium. Enzymes play critical role in the metabolic activities of all living organisms whether humans, animals, plants or microorganisms and are widely applied in microbialtechnology and their diagnosis processes. Abnormality of the enzyme metabolism system leads to a number of metabolic diseases. It is shown that many diseases associate with many components of the enzyme metabolism systems are now widely applied in clinical examinations as special markers for diseases. An interesting discovery suggesting that new roles of enzymes as a potential link that associates to prevent metabolic disorder. The aim of this review is to discuss the diverse diagnostic application of biotechnological enzymes for the purpose of teaching, education and research.
PDF Abstract XML References Citation
Received: August 07, 2010;
Accepted: November 15, 2010;
Published: February 22, 2011
How to cite this article
M. M.M. Raja, A. Raja, M. M. Imran, A. M.I. Santha and K. Devasena, 2011. Enzymes Application in Diagnostic Prospects. Biotechnology, 10: 51-59.
DOI: 10.3923/biotech.2011.51.59
URL: https://scialert.net/abstract/?doi=biotech.2011.51.59
DOI: 10.3923/biotech.2011.51.59
URL: https://scialert.net/abstract/?doi=biotech.2011.51.59
INTRODUCTION
Enzymes are produced by cellular anabolism, the naturally occurring biological process of making more complex molecules from simpler ones. Source organisms include bacteria, fungi, higher plants and animals (White and White, 1997). Enzymes may be extracted from a given source organism by a number of different methods (Nielsen et al., 1996). Most of the organisms that produce commercial enzymes are fungi. These organisms are molds Rhizopus oryzae, Aspergillus niger, Rhizomucor meihei, blights such as Endothia parasitica and yeasts such as Saccharomyces and Candida sp.
In cells and organisms most reactions are catalyzed by enzymes, which are regenerated during the course of a reaction. These biological catalysts are physiologically important because they speed up the rates of reactions that would otherwise be too slow to support life (Porcelli et al., 2010). Enzymes increase reaction rates, sometimes by as much as one million-fold, but more typically by about one thousand fold. Catalysts speed up the forward and reverse reactions proportionately so that, although the magnitude of the rate constants of the forward and reverse reactions is are increased, the ratio of the rate constants remains the same in the presence or absence of enzyme (Qijun et al., 2010). Since the equilibrium constant is equal to a ratio of rate constants, it is apparent that enzymes and other catalysts have no effect on the equilibrium constant of the reactions they catalyze.
Enzymes increase reaction rates by decreasing the amount of energy required to form a complex of reactants that is competent to produce reaction products. This complex is known as the activated state or transition state complex for the reaction. Enzymes and other catalysts accelerate reactions by lowering the energy of the transition state. The free energy required to form an activated complex is much lower in the catalyzed reaction (Jiang et al., 2007). The amount of energy required to achieve the transition state is lowered; consequently, at any instant a greater proportion of the molecules in the population can achieve the transition state. The result is that the reaction rate is increased.
While it is clear that enzymes are responsible for the catalysis of almost all biochemical reactions, it is important to also recognize that rarely, if ever, do enzymatic reactions proceed in isolation. The most common scenario is that enzymes catalyze individual steps of multi-step metabolic pathways, as is the case with glycolysis, gluconeogenesis or the synthesis of fatty acids. As a consequence of these lock-step sequences of reactions, any given enzyme is dependent on the activity of preceding reaction steps for its substrate (Abir et al., 2005). In humans, substrate concentration is dependent on food supply and is not usually a physiologically important mechanism for the routine regulation of enzyme activity. Enzyme concentration, by contrast, is continually modulated in response to physiological needs. Three principal mechanisms are known to regulate the concentration of active enzyme in tissues: (1) Regulation of gene expression controls the quantity and rate of enzyme synthesis. (2) Proteolytic enzyme activity determines the rate of enzyme degradation. (3) Covalent modification of preexisting pools of inactive proenzymes produces active enzymes.
Enzyme synthesis and proteolytic degradation are comparatively slow mechanisms for regulating enzyme concentration (Griffith et al., 2004) with response times of hours, days or even weeks. Proenzyme activation is a more rapid method of increasing enzyme activity but, as a regulatory mechanism, it has the disadvantage of not being a reversible process. Proenzymes are generally synthesized in abundance, stored in secretory granules and covalently activated upon release from their storage sites. Examples of important proenzymes include pepsinogen, trypsinogen and chymotrypsinogen, which give rise to the proteolytic digestive enzymes. Likewise, many of the proteins involved in the cascade of chemical reactions responsible for blood clotting are synthesized as proenzymes. Other important proteins, such as peptide hormones and collagen, are also derived by covalent modification of precursors.
Another mechanism of regulating enzyme activity is to sequester enzymes in compartments where access to their substrates is limited. For example, sequestering these enzymes within the lysosome controls the proteolysis of cell proteins and glycolipids by enzymes responsible for their degradation.
In contrast to regulatory mechanisms that alter enzyme concentration, there is an important group of regulatory mechanisms that do not affect enzyme concentration, are reversible and rapid in action and actually carry out most of the moment to moment physiological regulation of enzyme activity. These mechanisms include allosteric regulation, regulation by reversible covalent modification and regulation by control proteins such as calmodulin. Reversible covalent modification is a major mechanism for the rapid and transient regulation of enzyme activity. The best examples, again, come from studies on the regulation of glycogen metabolism where phosphorylation of glycogen synthase and glycogen phosphorylase kinase results in the stimulation of glycogen degradation while glycogen synthesis is coordinately inhibited. Numerous other enzymes of intermediary metabolism are affected by phosphorylation, either positively or negatively (Shinoda and Itoyama, 2003). These covalent phosphorylations can be reversed by a separate sub-subclass of enzymes known as phosphatases. Recent research has indicated that the aberrant phosphorylation of growth factor and hormone receptors, as well as of proteins that regulate cell division, often leads to unregulated cell growth or cancer. The usual sites for phosphate addition to proteins are the serine, threonine and tyrosine R group hydroxyl residues.
CLASSIFICATION OF ENZYME AND CO ENZYMES
This is a special commission of the International Union of Biochemistry (IUB) that made recommendations for the classification and naming of enzymes and for the definitions of the mathematical constants used in enzymology. The recommendations were first published in 1964 and were published in revised form in 1972, 1978 and 1984. This is the systematic arrangement and the naming of enzymes that is based on the 1972 recommendations of the Enzyme Commission of the International Union of Biochemistry (Stenesh, 1989). Reactions and the enzymes that catalyze them form 6 classes, each having 4-13 subclasses. The enzyme name has 2 parts. The first Soetan et al. 383 names the substrate or substrates. The second, ending in-ase, indicates the type of reaction catalysed. A number composed of four figures denotes each enzyme. The first figure denotes one of the six main divisions: oxidoreductases, transferases, hydrolases, lyases, isomerases and ligases. The second figure denotes the subclass and the third figure denotes the sub-subclass. The last figure denotes the serial number of the enzyme in its sub-subclass. The enzyme number is preceded by the abbreviation E.C. For example, E.C.2.7.1.1 denotes class 2 (a transferase), subclass 7 (transfer of phosphate), subsubclass 1 (an alcohol is the phosphate acceptor). The final digit denotes hexokinase, or ATP: D-hexose 6 -Phos-photransferase, an enzyme catalyzing phosphate transfer from ATP to the hydroxyl group on carbon 6 of glucose. There are 6 classes of enzymes as follows:
Oxidoreductases: These enzymes are involved in oxidations and reductions of their substrates e.g., alcohol dehydrogenase, lactate dehydrogenase, xanthine oxidase, glutathione reductase, glucose-6-phosphate dehydrogenase.
Transferases: These enzymes catalyze the transfer of a particular group from one substrate to another e.g., aspartate amino transferase (AST), alanine aminotransferase (ALT), hexokinase, phosphoglucomutase, hexose- 1-phosphate uridyltransferase, ornithine carbamoyl transferase etc.
Hydrolases: These enzymes bring about hydrolysis e.g., glucose- 6 -phosphatase, pepsin, trypsin, esterases, glycoside hydrolases etc.
Lyases: These are enzymes that facilitate the removal of small molecule from a large substrate e.g., fumarase, argino succinase, histidine decarboxylase.
Isomerases: These enzymes are involved in isomerization of substrate e.g., UDP-glucose, epimerase, retinal isomerase, racemases, triose phosphate isomerase.
Ligases: These enzymes are involved in joining together 2 substrates e.g., alanyl-t-RNA synthetase, glutamine synthetase, DNA ligases.
Many enzymes require a coenzyme which functions as group transfer reagents. Many enzymes that catalyze group transfer and other reactions require, in addition to their substrate, a second organic molecule known as a coenzyme, without which they are inactive. Coenzymes expand the repertoire of the catalytic capabilities of an enzyme far beyond those offered by the functional groups alone of the amino acids that constitute the bulk of the enzyme. Coenzymes that are tightly associated with an enzyme through either covalent bonding or non covalent forces are often referred to as prosthetic groups. Coenzymes that are freely diffusible generally serve as continually recycled carriers of hydrogen, flavin adenine dinucleotide (reduced) (FADH), hydride nicotinamide adenine dinucleotide (reduced) (NADH) and nicotinamide adenine dinucleotide phosphate (reduced) (NADPH), or chemical units such as acyl groups (coenzyme A) or methyl groups (folates), shuttling them between their points of generation and consumption. These latter coenzymes can thus be considered as second substrates. Enzymes that require coenzymes include those which catalyze oxidoreductions, group transfer and isomerization reactions and reactions that form covalent bonds (IUB classes 1, 2, 5 and 6). Lytic reactions, including the hydrolytic reactions catalyzed by the digestive enzymes, do not require coenzymes.
Coenzymes can be classified according to the group whose transfer they facilitate based on the above concept, coenzymes may be classified as follows: Coenzymes involved in transfer of groups other than hydrogen: Biotin, CoA-SH, cobamide (B12) coenzymes, folate coenzymes, pyridoxal phosphate, lipoic acid, sugar phosphates, thiamine pyrophosphate and Coenzymes involved in transfer of hydrogen. Nicotinamide adenine dinucleotide (oxidized) (NAD+), Nicotinamide adenine dinucleotide phosphate (oxidized) (NADP+). Flavin mononucleotide (FMN), Flavin Adenine Dinucleotide (FAD), lipoic acid, coenzyme Q (Murray et al., 2000).
ENZYME APPLICATION IN MICROBIAL TECHNOLOGY
There are many enzymes employed in biotechnological operations. These include:
SI Endonuclease: This enzyme is isolated from Aspergilus cryzae and it acts exclusively on ssDNA or RNA. It can break supercoiled DNA because it contains ss bubbles. It can also be used to distinguish supercoiled from both non-supercoiled, covalent circles and nicked circular DNA, both of which are resistant to the enzyme.
Restriction endonucleases: Restriction endonucleases are enzymes that cut DNA molecules at specific positions. It recognizes ‘foreign’ (unmodified) DNA at a specific site, and degrades it by internal cleavage. Most commonly used are the type II restriction enzymes, which cut within the recognition site. Some examples are EcoRI from E. coli, HindIII from Haemophilus influenzae, BamHI from Bacillus amyloliquefaciens, PstI from Providencia stuartii, SmaI from Serratia marcescens, Sau3A from Staphylococcus aureus, AluI from Arthrobacter luteus, TagI from Thermus aquaticus and HpaII from Haemophilus parainfluenza.
DNA polymerase I: It is primarily known as a ‘repair’ polymerase, which fills in single-stranded gaps. It is also involved in repair of the gaps formed on the lagging strand during replication. It also possesses both 5 I --- 3 I and 3 I --- 5 I exonuclease activity.
Klenow fragment of E. coli DNA polymerase 1: This enzyme is used for sequencing DNA using the Sanger Dideoxy System, filling the 3 recessed termini of restriction enzyme treated DNA and also used for labeling the termini of DNA fragments. The enzyme is also used for second strand cDNA synthesis in the cDNA procedures. ‘Klenow fragment’ is a proteolytic cleavage of DNA polymerase I. It leaves a fragment that is devoid of 5 I ---3 I exonuclease activity.
DNA polymerase III: This is the main ‘replication’ polymerase.
Helicase: It unwinds DNA, for example in conjugal plasmid transfer
S1 nuclease: This enzyme degrades single-stranded DNA
T4 DNA polymerase: This enzyme is used in the labeling of DNA fragments for use as hybridization probes.
RNA Polymerase: This enzyme synthesizes RNA, using a DNA template.
Primase: This is a special RNA polymerase, which makes a short primer required for DNA synthesis.
Replicase: This is RNA-directed RNA polymerase used in replication of some RNA viruses.
Ribonuclease (RNase): This enzyme degrades RNA molecules.
Rnase H: This is a specific RNAse which cuts RNA--- DNA hybrids. It is involved in replication of Col EI-like plasmid
Exonuclease: This is an enzyme that removes nucleotides from the ends of DNA fragments. A 5I ----3I exonuclease removes nucleotides from the 5I end, while a 3I ----5I exonuclease removes nucleotides from the 3Iend. This enzyme recognizes the terminal 5 I -phosphate of dsDNA for its exonuclease activity. Its primary use is the removal of protruding 5I terminus from dsDNA which is needed for the terminal transferase tailing of DNA (Daini, 2000).
Exonuclease III: This enzyme is used for generating linear template DNA for the dideoxysequencing technique and generating staggered ends on dsDNA due to its 3I ---- 5I exonuclease activity.
Ligases: It seals single-stranded gaps (nicks) in double-stranded DNA. It is also used for the formation of recombinant DNA molecules in gene cloning. T4 DNA Ligase: This enzyme catalysis the formation of a Phosphodiester bond between 3I-OH and 5I- phosphate ends in DNA using DNA molecules with cohesive ends as substrate.
Alkaline phosphatase: In gene cloning, this enzyme is used to remove phosphate groups from the 5I end of DNA molecules and also used as a reporter gene for identification of secretion signals.
Polynucleotide kinase: It transfers a phosphate group from ATP to the 5I OH end of DNA or RNA. T4 Polynucleotide kinase is an enzyme is isolated from T4 infected E. coli and catalyzes the transfer of-phosphate of ATP to a 5I -OH end in DNA or RNA. It is also used for the labeling of 5I termini of DNA for Maxam and Gilbert DNA sequencing and the phosphorylation of DNA lacking 5I P termini.
Reverse transcriptase: This is RNA-directed DNA polymerase. It synthesizes DNA (complementary DNA) using mRNA template. For example Reverse transcriptase is an enzyme coded for by avian myeloblastosis virus which catalyses the synthesis of cDNA from an RNA template. It can also be used for the labeling of termini of DNA with extended 5I ends.
Topoisomerase: This is a class of enzymes that alters the conformation of DNA, for example by changing the degree of winding or super coiling.
Transposase: This enzyme catalysis the initial steps in transpositions.
Terminal transferase: This adds nucleotides to the 3I end of DNA, without requiring a template strand.
Terminal deoxynucleotidyl transferase: This enzyme is isolated from calf thymus and catalyses the addition of dNTP to the 3 I -OH of DNA molecules. One of the primary uses of terminal transferase is the tailing of vectors and cDNA with complementary bases, thus permitting the cloning of the cDNA fragments. It can also be used for labeling of 3I ends of DNA fragments.
IMMOBILIZED ENZYME
Since the second half of the last century, numerous efforts have been devoted to the development of insoluble immobilized enzymes for a variety of applications these applications can clearly benefit from use of the immobilized enzymes rather than the soluble counterparts, for instance as reusable heterogeneous biocatalysts, with the aim of reducing production costs by efficient recycling and control of the process (Reetz et al., 2000) as stable and reusable devices for analytical and medical applications as selective adsorbents for purification of proteins and enzymes as fundamental tools for solid-phase protein chemistry and as effective micro devices for controlled release of protein drugs (Fig. 1).
This is an enzyme that is physically confined while it carries out its catalytic function. This may occur naturally, as in the case of particulate enzymes, or itmay be produced artificially by chemical or by physical methods (Stenesh, 1989).
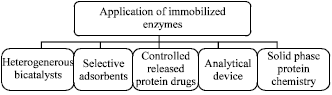 | |
| Fig. 1: | Range of application of immobilized enzymes |
In the chemical methods, the enzyme is linked covalently to a support. These methods include attachment of the enzyme to a water-insoluble support, incorporation of the enzyme into a growing polymer chain, or cross linking of the enzyme with a multifunctional low molecular weight reagent (Van Roon et al., 2002). In the physical methods, the enzyme is not linked covalently to a support. These methods include adsorption of the enzyme to a water-insoluble matrix, entrapment of the enzyme within either a water-insoluble gel or a microcapsule, or containment of the enzyme within special devices equipped with semi permeable membrane (Soares et al., 2001) . Expensive enzymes can be recovered and used again. The enzyme can also be used in a variety of configurations of bioreactors that permit continuous operation (Coombs, 1992).
ENZYME IMMUNE SYSTEM
Enzymes become part of the immune system (Alpha-2-macroglobulin), working with it and facilitating its function. The modulation of growth factor binding properties of alpha 2-macroglobulin by enzyme therapy. Ingestion of proteinases was found to trigger the formation of intermediate forms of alpha-2- macroglobulin displaying high affinity of transforming growth factor- beta (TGF-β). They observed maximum binding of TGF- β 1 - 2 h after bolus ingestion and steadily leveled off with time. They concluded that intestinal absorption of proteinase triggers the formation of TGF-β binding species of alpha 2 macroglobulin in blood mediated by this process of high concentrations of TGF- β might be reduced via enhanced clearance of alpha 2-macroglobulin TGF- β complexes. Thus, proteinase therapy may have beneficial effects in treatment of fibrosis and certain cancers accompanied by excessively high TGF- β concentrations. The oral therapy with proteolytic enzymes decreases excessive TGF-beta levels in human blood. In the study, they stated that therapy with oral proteolytic enzymes (OET) with combination drug products containing papain, bromelain, trypsin and chymotrypsin have been shown to be beneficial in clinical settings such as radiotherapyinduced fibrosis, bleomycin pneumotoxicity and immunosuppression in cancer, all of which are nowadays knowned to be accompanied by excessive transforming growth factor-beta (TGF- β) production. They concluded that their results support the concept that OET is beneficial in diseases characterized in part by TGF- β1 overproduction.
ENZYMES AS MARKERS FOR DISEASE
Some enzymes are found only in specific tissues or in a limited number of such tissues. For example, lactase dehydrogenase (LDH) has 2 different forms, called isozymes, in heart and skeletal muscle. Two forms differ slightly in amino acid composition and Can be separated on the basis of charge as a result. Since LDH is a tetramer of four subunits, it too can exist in 5 different forms depending on the source of the subunits. An increase of any form of LDH in the blood indicates some kind of tissue damage. A heart attack can usually be diagnosed with certainty if there is an increase of LDH from heart. Also, there are different forms of Creatine Kinase (CK), an enzyme that occurs in the brain, heart and skeletal muscle. Appearance of the brain type can indicate a stroke or a brain tumour, whereas the heart type indicates a heart attack. After a heart attack, CK shows up more rapidly in the blood than LDH. Monitoring the presence of both enzymes extends the possibility of diagnosis, which is useful, since a very mild heart attack might be difficult to diagnose. An elevated level of the isozyme from heart in blood is a definite indication of damage to the heart tissue (Drolet et al., 2007). Another useful enzyme assayed is acetyl cholinesterase (AChE), which is important in controlling certain nerve impulses. Many pesticides affect this enzyme, so farm workers are often tested to be sure that they have not received inappropriate exposure to these important agricultural toxins. There are several enzymes that are typically used in the clinical laboratory to diagnose diseases. There are highly specific markers for enzymes active in the pancrease, red blood cells, liver, heart, brain, prostate gland and many of the endocrine glands. Since these enzymes are relatively easy to assay using automated techniques, they are part of the standard blood test veterinary and medical doctors are likely to need in the diagnosis and treatment/management of diseases.
ENZYME AS DRUG OR ANTIBIOTICS
New antibiotics that are active against resistant bacteria (Raja et al., 2010) have lived on earth for several billion years. During this time, they encountered in nature a wide range naturally occurring antibiotics or drugs. To survive, bacteria developed antibiotic resistance mechanism (Hoskeri et al., 2010). Enzymes as drugs have two important features that distinguish them from all other types of drugs. First, enzymes often bind and act on their targets with great affinity and specificity. Second, enzymes are catalytic and convert multiple target molecules to the desired products. These two features make enzymes specific and potent drugs that can accomplish therapeutic biochemistry in the body that small molecules cannot. These characteristics have resulted in the development of many enzyme drugs for a wide range of disorders (Vellard, 2003).
ENZYMES IN THE DIAGNOSIS OF PATHOLOGY
The measurement of the serum levels of numerous enzymes has been shown to be of diagnostic significance. This is because the presence of these enzymes in the serum indicates that tissue or cellular damage has occurred resulting in the release of intracellular components into the blood (Machetti et al., 1998). Hence, when a physician indicates that he/she is going to assay for liver enzymes, the purpose is to ascertain the potential for liver cell damage. Commonly assayed enzymes are the amino transferases: alanine transaminase, ALT (sometimes still referred to as serum glutamate-pyruvate aminotransferase, SGPT) and aspartate aminotransferase, AST (also referred to as serum glutamate-oxaloacetate aminotransferase, SGOT); lactate dehydrogenase, LDH; creatine kinase, CK (also called creatine phosphokinase, CPK); gamma-glutamyl transpeptidase, GGT. Other enzymes are assayed under a variety of different clinical situations. Many enzymes are involved in the clinical diagnoses of various diseases in human and veterinary medicine (Nielsen et al., 1996). These enzymes facilitate or enhance rapid diagnoses of these diseases. These enzymes could be classified into many classes. They are:
Alkaline phosphatase: Alkaline phosphatases were the earliest serum enzymes to be recognized to have clinical significance, when in the 1920s, it was discovered that they increase in bone and liver diseases . Since then, they have been the subject of more publications than any other enzyme (Lone et al., 2003). Alkaline phosphates are a group of isoforms which hydrolyse many types of phosphate esters, whose natural substrate or substrates are unknown. The term ‘>alkaline’= refers to the optimal alkaline pH of this class of phosphatases in vitro. In both humans and animals, the major sources of ALPs are the liver, bone, kidney and placenta. In humans, it is involved in bone and hepatobiliary diseases. ALPs are also of diagnostic importance in animal diseases. Total serum ALP activity has diagnostic value in the hepatic and bone diseases in dogs and cats. It is of little value in hepatic diseases of horses and ruminants because of the broad range of reference values against which the patients’ values must be compared. The range of serum ALP value in goats may be 10-fold with no evidence of hepatic damage. Values within the individual are fairly constant for sequential evaluation.
Creatine kinase: Creatine kinase isozymes are the most organ-specific serum enzymes in clinical use. They catalyse the reversible phosphorylation of creatine by ATP to form creatine phosphate, the major storage form of high-energy phosphate required by muscle. Creatine kinases are found in many parts of the body like the heart, brain, skeletal muscle and smooth muscle but they have their highest specific activity in the skeletal muscle (Aksenova et al., 2000) . In humans, Creatine kinase is associated with myocardial infarction and muscle diseases. Increase in Creatine kinase in cerebrospinal fluid has been associated with a number of disorders in dogs, cats, cattle and horses. The Creatine kinase are such sensitive indicators of muscle damage that, generally, only large increases in serum activity are of clinical significance.
Alanine aminotransferase: It was formerly known as Glutamic Puruvate Transaminase; (GPT). It catalyses the reversible transamination of Lalanine and 2-oxoglutarate to pyruvate and glutamate in the cytoplasm of the cell. ALT can be found in the liver, skeletal muscle and heart. The greatest specific activity of ALT in primates, dogs, cats, rabbits and rats is in the liver. It is a well established, sensitive liver-specific indicator of damage. However, ALT in the tissues of pigs, horses, cattle, sheep or goats is too low to be of diagnostic value ( Kikuchi et al., 1999). It is used as an indicator of hepatopathy in toxicological studies which use small laboratory rodents as well as dogs.
Aspartate aminotransferase: It wasformerly called Glutamic Oxaloacetic Transaminase; (GOT). It catalyses the transamination of L-aspartate and 2-oxoglutarate to oxaloacetate and glutamate. AST is found in skeletal muscle, heart, liver, kidney and erythrocytes and is associated with myocardial, hepatic parenchymal and muscle diseases in humans and animals. The pre-sence of AST in so many tissues make their serum level a good marker of soft tissue but precludes its use as an organspecific enzyme (Bittinger et al., 2003). Red blood cells contain a large amount of AST which leaks into plasma before haemolysis is seen.
Sorbitol dehydrogenase (SDH): It is also called L-iditol dehydrogenase; (IDH). It catalyses the reversible oxidation of D-sorbitol to D-fructose with the cofactor NAD. The plasma activity is low in dog and horse plasma but appreciably greater in cattle, sheep, and goat serum. Aside from the testes, it is found in appreciable amounts only in hepatocytes (El-Kabbani et al., 2004). As a result of this, an increase in plasma SDH is consistent with hepatocyte damage. SDH is liver specific in humans and all species of animals and hepatic injury appears to be the only source of increased SDH activity. Although SDH is liver specific in all species, the already established usage of ALT in dogs and cats has limited SDH as a diagnostic indicator of hepatocellular damage to horses, cattle, sheep and goats.
Lactate dehydrogenases(LDH): It catalyses the reversible oxidation of pyruvate to L(+) lactate with the cofactor NAD. The equilibrium favours lactate formation, but the preferred assay method is in the direction of pyruvate because pyruvate has an inhibitory effect on LDH. Lactate dehydrogenase has isoenzymes. LDH can be found in the heart, liver, erythrocyte, skeletal muscle, platelets and lymph nodes. In humans, it is involved in myocardial infarction, haemolysis and liver disease. LDH isoenzyme profiles were the first isoenzyme profiles used in clinical veterinary medicine in an attempt to detect specific organ damage. The introduction of more highly organ-specific procedures has resulted in LDH no longer being in common use in veterinary medicine (Murray et al., 2000)
Cholinesterase (ChE): Serum cholinesterase (ChE) activity is composed of two distinct cholinesterases. The major substrate is acetylcholine, the neurotransmitter found at the myoneural junction. Acetylcholinesterase (AChE; EC 3.1.1.7) found at the myoneural junction is the true ChE and is essential in hydrolyzing acetylcholine so that the junction can be reestablished and prepared for additional signals (Ellis, 2005). The myoneural junction AChE is also found in Red Blood Cells (RBC), mouse, pig, brain and rat liver. Only a small amount of AChE is found in plasma. The ChE of plasma is a pseudocholine sterase, butylcholinesterase (ButChE; EC 3.1.1.8), which hydrolyses butyrylcholine four times faster than acetylcholine and is also located in white matter of the brain, liver, pancrease and intestinal mucosa . Decreases in ButChE have been reported in humans with acute infection, muscular dystrophy, chronic renal disease and pregnancy, as well as insecticide intoxification.
Lipase: Serum pancreatic lipases (EC 3.1.1.3; triacylglycerol lipase) catalyse the hydrolysis of triglycerides preferentially at the 1 and 3 positions, releasing two fatty acids and a 2-monoglyceride. Lipase can be found in the pancrease and hepatobiliary tract and is involved in pancreatitis and hepatobiliary disease (Nduka, 1999).
Amylase: Amylases are calcium-dependent metalloenzymes that randomly catalyze the hydrolysis of complex carbohydrates, e.g., glycogen at the -1-4 linkages. The products of this action are maltose and limit dextrins. The enzyme is a Ca2+ metalloenzyme which requires one of a number of activator ions such as Cl-or Br-. Amylase can be found in the salivary glands, pancrease and ovaries and is used as a diagnostic aid for pancreatitis (Gupta et al., 2001).
Glutamyltransferase: This is a carboxypeptidase which cleaves C-terminal glutamyl groups and transfers them to peptides and other suitable acceptors. It is speculated that GGT is associated with glutathione metabolism (Kaneko, 1989). The major sources are the liver and kidney and are involved in hepatobiliary disease and alcoholism. Cholestatic disorders of all species examined result in increased serum GGT activity.
Trypsin: Trypsins are serum proteases which hydrolyse the peptide bonds formed by lysine or arginine with other amino acids. The pancreas as the zymogen trypsinogen, which is converted to tyrosine by intestinal enterokinase or trypsin itself, secretes them.
Glutathione peroxidases: These are metalloenzymes containing four atoms of selenium per molecule of enzyme. They catalyze the oxidation of reduced glutathione by peroxide to form water and oxidized glutathione. Because of the high concentration of selenium in glutathione peroxidases, there is a good direct correlation between the amount of red blood cell GPx activity and the selenium concentration of other organs (Chatterjea and Shinde, 2002). Other enzymes with disease diagnosis applications are acid phosphatase (ACP), found in prostate and erythrocytes and are used in diagnosis of prostate carcinoma. Aldolase (ALD), found in skeletal muscle and heart and involved in muscle disease. Glutamate dehydrogenase (GLDH), found in the liver is used to diagnose hepatic parenchymal disease. Hydroxybutyrate dehydrogenase (HBD), which is the heart form of lactate dehydrogenase is involved in myocardial infarctionJust as enzyme assay is used to diagnose diseases in humans and animals, it may also be applied to the investigation of diseases in plants. For example, it has been found that an injury (either mechanical or pathogenic) results in a marked, localized increase in the activity of glucose-6-phosphate dehydrogenase, but not of glucose phosphate isomerase, indicating diversion of glucose breakdown from glycolysis to the pentose phosphate pathway.
ENZYMES USED IN IMMUNOASSAYS
Enzymes may also be used as an alternative to radioisotopes as markers in immunoassays have been used for the determination of a variety of proteins and hormones. The role of enzymes in immunoassay used to replace radioisotopes as markers, since they are not hazardous to health and can be detected by techniques which are more generally available. Any enzyme with a sensitive and convenient assay procedure can be used for this purpose. Two common examples of enzyme immunoassay (EIA) procedures are enzyme-linked immunosorbent assay (ELISA) and Enzyme-Multiplied Immunoassay Test (EMIT). ELISA is a highly sensitive assay that can be used to detect either antigen or antibody. Applications of ELISA include diagnostics for noninfectious diseases involving hormones, drugs, serum components, oncofetal proteins, or autoimmune diseases, as well as diagnostics for infectious diseases caused by bacterial, viral, mycotic or parasitic organisms. The enzymes frequently used in ELISA are Horseradish peroxidase, alkaline phosphatase and βgalactosidase. In EMIT, the activity of malate dehydrogenase is assayed by standard enzyme methodology for the detection of thyroxine by Enzyme-labelled immunoassay.
ENZYMES ACTED AS THERAPEUTIC AGENTS
In a few cases enzymes have been used as drugs in the therapy of specific medical problems (Devlin, 1986). Streptokinase is an enzyme mixture prepared from streptococcus. It is usefull in clearing blood clots that occur in the lower extremities. Streptokinase activates the fibrinolytic proenzyme plasminogen that is normally present in plasma. The activated enzyme is plasmin is a serine protease like trypsin that attacks fibrin, cleaving it into several soluble components. Another enzyme of therapeutic importance is asparaginase. Asparaginase therapy is used for some types of adult leukemia. Tumor cells have a nutritional requirement for asparagine and must scavenge it from the host’s plasma. By administering asparaginase i.e., the host’s plasma level of asparagine is markedly depressed, which results in depressing the viability of the tumor. Enzyme replacement in individuals that are genetically deficient in a particular enzyme are also applications of enzymes as therapeutic agents. Also, enzymes such as u-plasminogen activator, formerly known as urokinase, extracted from human urine, can be infused into the blood stream of patients at risk from a pulmonary embolism (a fragment of a blood-clot lodging in the pulmonary artery): these enzymes stimulate a cascade system responsible for the production of active plasmin, a proteolytic enzyme which digests fibrin, the main structural component of blood-clots. Some enzymes may also be used to restrict the growth of cancer cells by depriving them of essential nutrients: for example, Lasaparaginase may be used in the treatment of several types of leukaemia, since the tumour cells, in contrast to normal cells, have a requirement for exogenous Lasaparagine. Another example of therapeutic application of enzymes is the use of immobilized enzymes as components of artificial kidney machines, which are used to remove urea and other waste products from the body, where kidney disease prevents this being done by natural processes (Palmer, 2001). Urea enters the machine from the blood, by dialysis (termed haemodialysis) and is converted to CO2 and NH4+ by immobilized urease; toxic NH4+ is then either trapped on ion exchange resins or incorporated into glutamate by the action of immobilized glutamate dehydrogenase linked to alcohol dehydrogenase to ensure coenzyme recycling, before the fluid is returned to the blood stream.
ACKNOWLEDGMENT
We thank the Department of Microbiology, Jamal Mohamed College (Autonomous), Tiruchirappalli-620 020 for supporting and fulfilling all the needs to carry out this work.
REFERENCES
- Aksenova, M., D.A. Butterfield and W.R. Markesbery, 2000. Oxidative modification of creatine kinase BB in Alzheimer`s disease brain. J. Neurochem., 74: 2520-2527.
CrossRef - Bittinger, M.A., L.P Nguyen and C.A. Bradfield, 2003. Aspartate aminotransferase generates proagonists of the aryl hydrocarbon receptor. Mol. Pharmacol., 64: 550-556.
CrossRef - Drolet, R., M. Simard, J. Plante, P. Laberge and Y. Tremblay, 2007. Human type 2 17 betahydroxysteroid dehydrogenase mRNA and protein distribution in placental villi at mid and term pregnancy. Reprod. Biol. Endocrinol., 5: 30-30.
CrossRef - Ellis, J.M., 2005. Cholinesterase inhibitors in the treatment of dementia. J. Am. Osteopath. Assoc., 105: 145-158.
PubMedDirect Link - Abir, F., S. Alva, L. Kaminski, L. Donald and E. Walter, 2005. The role of arachidonic acid regulatory enzymes in colorectal disease. Dis. Colon Rectum, 48: 257-303.
Direct Link - Griffith, K.L., I.M. Shah and R.E. Wolf, 2004. Proteolytic degradation of Escherichia coli transcription activators SoxS and MarA as the mechanism for reversing the induction of the superoxide (SoxRS) and multiple antibiotic resistance (Mar) regulons. Mol. Microbiol., 51: 1801-1816.
Direct Link - Gupta, K.B., V. Ghalaut, R. Gupta, S. Tandon and P. Prakash, 2001. Estimation of serum and pleural fluid amylase and iso-enzyme in cases of malignant pleural effusion. Ind. J. Tub, 48: 87-87.
Direct Link - Hoskeri, H.J., V. Krishna and C. Amruthavalli, 2010. Effects of extracts from lichen Ramalina pacifica against clinically infectious bacteria. Researcher, 2: 81-85.
Direct Link - Jiang, X., L. Jing and M. Lan, 2007. Metabolic enzymes link morphine withdrawal with metabolic disorder. Cell Res., 17: 741-743.
CrossRef - El-Kabbani, O., C. Darmanian and R.P.T. Chung, 2004. Sorbitol dehydrogenase: Structure, function and ligand design. Curr. Med. Chem., 11: 465-476.
Direct Link - Kikuchi, H., S. Hirose, S. Toki, A. Kazuhito and F. Takaiwa, 1999. Molecular characterization of a gene for alanine aminotransferase from rice (Oryza sativa). Plant Mol. Biol., 39: 149-159.
PubMed - Lone, M.A., A. Wahid, S.M. Saleem, P. Koul, G.H. Nabi Dhobi and A. Shahnawaz, 2003. Alkaline phosphatase in pleural effusions. Indian J. Chest Dis. Allied Sci., 45: 161-163.
Direct Link - Machetti, M., M. Feasi and N. Mordini, M.T. Van Lint and A. Bacigalupo et al., 1998. Comparison of an enzyme immunoassay and a latex agglutination system for the diagnosis of invasive aspergillosis in bone marrow transplant recipients. Bone Marrow Transplant., 21: 917-921.
Direct Link - Vellard, M., 2003. The enzyme as drug: Application of enzymes as pharmaceuticals. Curr. Opin. Biotechnol., 14: 444-450.
PubMed - Nielsen, K.H., L. Kelly, D. Gall, S. Balsevicius, J. Bosse, P. Nicoletti and W. Kelly, 1996. Comparison of enzyme immunoassays for the diagnosis of bovine brucellosis. Prev. Vet. Med., 26: 17-32.
CrossRef - Porcelli, A.M., A. Ghelli, C. Ceccarelli, M. Lang and G. Cenacchi et al., 2010. The genetic and metabolic signature of oncocytic transformation implicates HIF1{alpha} destabilization. Human Mol. Genet., 19: 1019-1032.
CrossRef - Qijun, W., Y. Zhang, C. Yang, H. Xiong and Y. Lin et al., 2010. Acetylation of metabolic enzymes coordinates carbon source utilization and metabolicflux. Science, 327: 1004-1007.
CrossRef - Raja, A., P. Prabakaran and P. Gajalakshmi, 2010. Isolation and screening of antibiotic producing psychrophilic actinomycetes and its nature from rothang hill soil against viridans Streptococcus sp. Res. J. Microbiol., 5: 44-49.
CrossRefDirect Link - Reetz, M.T., R. Wenkel and D. Avnir, 2000. Entrapment of lipases in hydrophobicsol-gel materials: Efficient heterogeneous biocatalysts in aqueous medium. Synthesis, 6: 781-783.
CrossRef - Soares, C.M.F., H.F. de Castro, M.H.A. Santana and G.M. Zanin, 2001. Selection of stabilizing additive for lipase immobilization on controlled pore silica by factorial design. Applied Biochem. Biotechnol., 91: 703-718.
PubMed - Shinoda, T. and K. Itoyama, 2003. Juvenile hormone acid methyltransferase: A key regulatory enzyme for insect metamorphosis. PNAS, 100: 11986-11991.
CrossRef - Van Roon, J., R. Beeftink, K. Schroen and H. Tramper, 2002. Assessment of intraparticle biocatalytic distribution as a tool inrational formulation. Curr. Opin. Biotechnol., 13: 398-405.
CrossRef