
M.A. Faramarzi
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
M. Fazeli
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
M. Tabatabaei Yazdi
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
S. Adrangi
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
K. Jami Al Ahmadi
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
N. Tasharrofi
Biotechnology Research Center, Department of Pharmaceutical Biotechnology, Faculty of Pharmacy, Tehran University of Medical Sciences, P.O. Box 14155-6451, Tehran 14174, Iran
F. Aziz Mohseni
Iranian Research Organization for Science and Technology, Tehran, Iran







ABSTRACT
The aim of this study was to characterize chitinase-producing bacteria isolated from environmental samples and to investigate conditions affecting chitinase production by these bacteria. Ninety-eight isolates recovered from 20 soil samples were screened for chitinase production. Eighteen isolates showed chitinolytic activity, among which isolate U2 was selected for further study based on dinitrosalisylic acid assay results. The isolate U2 was identified as Massilia timonae through phenotypic characterization and 16S rDNA sequencing and the optimal conditions for chitinase production were determined to be 25-30°C, initial pH 6.0-6.5 and chitin concentration of 1% (w/v). The maximum chitinolytic activity was achieved after 36 h of incubation. The addition of different nitrogen sources to the production medium had no significant effect on chitinase production. Among various carbon sources tested, N-acetylglucosamine (GlcNAc), fructose, lactose, maltose and glucose showed modest inhibitory effect while arabinose did not affect enzyme production by M. timonae isolate U2. The addition of Triton X100 increased chitinase production by 12.4%. The enzyme was reasonably stable in the pH range 5-7 and at temperatures up to 50°C. These results indicate that M. timonae is capable of producing chitinase in relatively simple media containing colloidal chitin as the sole carbon and nitrogen source.
PDF Abstract XML References Citation
How to cite this article
M.A. Faramarzi, M. Fazeli, M. Tabatabaei Yazdi, S. Adrangi, K. Jami Al Ahmadi, N. Tasharrofi and F. Aziz Mohseni, 2009. Optimization of Cultural Conditions for Production of Chitinase by a Soil Isolate of Massilia timonae. Biotechnology, 8: 93-99.
DOI: 10.3923/biotech.2009.93.99
URL: https://scialert.net/abstract/?doi=biotech.2009.93.99
DOI: 10.3923/biotech.2009.93.99
URL: https://scialert.net/abstract/?doi=biotech.2009.93.99
INTRODUCTION
Chitin, the second most abundant polymer in nature after cellulose, is made of N-acetylglucosamine (GlcNAc) units linked together with β (1→4) glycosidic bonds (Duo-Chuan, 2006). It is found in the exoskeleton of crustaceans and insects and the cell wall of fungi as a structural component. Chitinolytic enzymes hydrolyze chitin randomly at internal sites (endochitinase, EC 3.2.1.14) or cleave GlcNAc mono- and disaccharides from the nonreducing end of chitin chains (β-N-acetylhexosaminidase, EC 3.2.1.52) (Dahiya et al., 2005).
Chitinases have diverse physiological functions. In fungi, crustaceans and insects they are mainly involved in morphogenesis. In plants and probably vertebrates, chitinases play a role in defense against pathogens. On the other hand, bacteria produce chitinolytic enzymes to meet nutritional needs (Wiwat et al., 1999; Jami Al Ahmadi et al., 2008a, b). As a result, chitinolytic enzymes have found a wide range of applications including biological control of plant pathogens and vectors of human diseases, production of single cell proteins, isolation of fungal protoplasts and potentially, treatment of human fungal diseases. In addition, some chitooligosaccharides including chitohexaose and chitoheptaose show antitumor properties. Such chitooligosaccharides can be obtained using chitinolytic enzymes. However, obtaining oligomers of the desired length requires a particular combination of chitinolytic enzymes with high endo- and low exo-chitinase activity (Patil et al., 2000). Therefore, the identification of new chitinolytic enzymes with different characteristics may be valuable for the pharmaceutical industry.
Massilia timonae, the first species of its genus to be described, was first isolated from a patient with common variable immunodeficiency in 1998 (La Scola et al., 1998). Other species of the genus including M. dura, M. albidiflava, M. plicata, M. lutea and M. aurea have been isolated from soil (Zhang et al., 2006) or water (Gallego et al., 2006) samples. The objective of this study was to screen the chitin-degrading ability in several bacterial strains isolated from environmental samples and to optimize the conditions for chitinase production by these microorganisms in liquid culture medium. Among the isolated bacteria, the M. timonae isolate U2 showed the highest ability to degrade chitin and was selected for further study. To our knowledge this is the first report on chitinase production by this species.
MATERIALS AND METHODS
This study was conducted in 2006-2008 by Tehran University of Medical Sciences, in Tehran, Iran.
Culture media: The basic medium used for fermentation contained the following constituents (g L-1): Na2HPO4 (0.65), KH2PO4 (1.5), NaCl (0.25), NH4Cl (0. 5), MgSO4 (0.12), CaCl2 (0.005) and colloidal chitin (10), pH 6.5 (Wen et al., 2002). Cultivation was done using 500 mL Erlenmeyer flasks containing 100 mL medium. Chitin agar medium containing the basic medium and 20 g L-1 of agar was used for screening the bacterial strains in plates.
Isolation of chitinase-producing bacteria: Twenty soil samples were collected between March and May 2006 from various locations in Tehran, Iran. Soil samples (1 g) were shaken in sterile distilled water (100 mL) containing Tween 80 (0.5%, v/v) for 30 min and 0.1 mL of the suspension was inoculated onto chitin agar plates, followed by incubation at 30°C for 10 days. Plates were examined for formation of clearing zones (CZ). Both the size of the CZ and the colony size (CS) were determined and the colonies were transferred to new plates for further investigation.
Selection of high chitinase producers: Isolates which showed a higher CZ/CS ratio in the chitin agar plate assays were inoculated into 100 mL of liquid medium and incubated at 30°C for at least 96 h with agitation at 150 rpm. Samples were taken every 12 h, centrifuged at 3000 rpm for 10 min and the chitinase activity was determined in the supernatant.
Identification: The isolate used in this study was identified as M. timonae based on phenotypic characterization and 16S rDNA sequence analysis. In order to obtain genomic DNA, bacterial cells were harvested from overnight cultures, washed three times with apyrogen sterile distilled water and boiled for 10 min to release the DNA. The suspension was centrifuged at 14000 rpm for 5 min and the supernatant was used as DNA template for PCR amplification of the 16s rDNA gene. The PCR amplification program included an initial denaturation at 94°C for 180 sec, 30 cycles of (denaturation 94°C for 60 sec, annealing 60°C for 45 sec and extension 72°C for 90 sec) and a final extension at 72°C for 90 sec. A large fragment of the 16s rDNA gene was amplified using primers 27F (5’-AGAGTTTGATCCTGGCTCAG-3’) and 1492R (5’-TACGGTTACCTTGTTACGACTT-3’) and sequenced with primers 27F, 515F (5’-GTGCCAGCAGCCGCGGT-3’), 1099F (5’-GCAACGAGCGCAACCC-3’), 517R (5’-GTATTACCGCGGCTGCTGGC-3’), 1115R (5’-AGGGTTGCGCTCGTTG-3’) and 1492R (Furushita et al., 2003). The 1499 bp sequence was submitted to the NCBI Nucleotide database (accession number EU274637).
Preparation of colloidal chitin: Colloidal chitin was prepared according to the method described by Robert and Selitrennikoff (1988) with minor modifications. Briefly, 10 g of chitin powder from crab shell (Fluka, USA) was added into 150 mL of concentrated HCl and stirred overnight at 4°C. The mixture was added slowly to 1000 mL of ice cold ethanol (96%) and kept overnight at room temperature with vigorous stirring. The precipitate was collected by centrifugation at 5000 rpm for 20 min, washed three times with 0.1 M sodium phosphate buffer pH 7.0 and stored at 4°C until used.
Enzyme assay: Supernatants from M. timonae cultures were harvested by centrifugation at 3000 rpm for 10 min. The reaction mixture consisted of 1 mL of culture supernatant and 1 mL of 1% (w/v) colloidal chitin in citrate-phosphate buffer pH 5. The mixture was incubated at 50°C, 125 rpm for 30 min. The remaining chitin was removed by centrifugation at 3000 rpm for 10 min. Chitinolytic activity was assayed by measuring the release of reducing sugars from colloidal chitin by the dinitrosalycilic acid (DNS) method (Driss et al., 2005). An aliquot of 1.5 mL of the supernatant and 1.5 mL of DNS solution were mixed, heated at 100°C for 15 min and the absorbance of the reaction mixture at 540 nm was measured after cooling to room temperature. One enzyme unit was defined as the amount of enzyme that produces 1 μmol reducing sugar per hour under the reaction conditions.
Effects of cultural conditions and media composition on chitinase production: The effects of temperature, pH and substrate concentration on chitinolytic enzyme production by M. timonae were studied by growing cultures at temperatures between 15 and 40°C, initial pH between 5 and 8 and colloidal chitin concentrations of 0.25, 0.5, 1, 1.5 and 2% (w/v), respectively. All experiments were carried out in 500 mL Erlenmeyer flasks containing 100 mL culture medium incubated at 30°C (except for temperature experiments) and 150 rpm for 36 h.
In order to determine the effect of different nitrogen sources (casein, peptone, tryptone and yeast extract, 0.1%, w/v), carbon sources (arabinose, fructose, GlcNAc, glucose, lactose and maltose, 0.1%, w/v) and detergents (Tween 20, Tween 80 and Triton X100, 0.05 %, w/v) on chitinase production, the medium was separately supplemented with each compound and incubated at 30°C and 150 rpm for 36 h. All experiments were performed in triplicate.
Physico-chemical characteristics of chitinase: To evaluate the effect of temperature on the stability of chitinase from M. timonae isolate U2, enzyme solutions were incubated for 1 h at temperatures between 25 and 60°C and the residual activity was assayed after cooling. The pH stability was determined by incubating the enzyme solution at pH values between 4 and 10 for 5 h at ambient temperature.
Statistical analysis: Data were analyzed through analysis of variance (ANOVA) using SPSS 11.5 for Windows (SPSS Inc., Chicago, IL, USA). Statistically significant results were determined by a p-value of <0.05.
RESULTS AND DISCUSSION
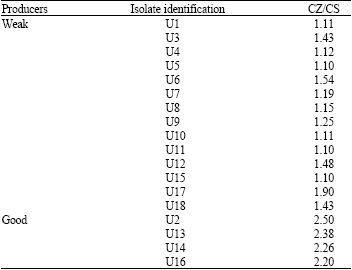
Isolation and identification: To identify efficient chitin-degrading bacteria, 20 soil samples collected from various locations were screened by culturing them on chitin agar plates and searching for clearing zones produced around the colonies. A total number of 98 colonies were observed when inoculated onto chitin agar plates. Eighteen of them produced clearing zones. These strains, designated U1 to U18, were classified on the basis of their CZ/CS ratio as good (CZ/CS>2) or weak (CZ/CS<2) producers (Table 1). Isolates U2, U13, U14 and U16 which showed a higher CZ/CS ratio were transferred into liquid culture medium and their chitinolytic activity was measured by the DNS method (Fig. 1). As can be seen in the Fig. 1, isolates U2 and U14 produced the highest amounts of enzyme (9.9 and 8.3 U mL-1, respectively) among the four isolates. Although these two isolates showed similar chitinolytic activity (p = 0.082), the time required for isolate U2 to reach its maximum activity (36 h) was much shorter than for isolate U14 (72 h).
| Table 1: | Ratio of CZ/CS of various isolates. Isolates were classified according to CZ/CS ratio as good (CZ/CS>2) or weak (CZ/CS<2) chitinase producers |
 | |
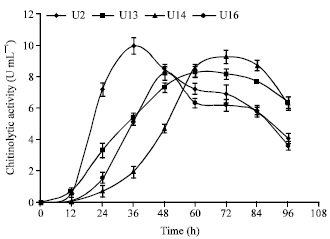 | |
| Fig. 1: | Time course of chitinolytic enzyme production by isolates U2, U13, U14 and U16. Each point represents the mean of three independent experiments and error bars indicate SD |
Based on these results, isolate U2 was selected for further study and identified through 16S rDNA sequence analysis and phenotypic characterization. A BLAST search of the 16S rDNA sequence against NCBI Nucleotide database revealed 97% identity with several members of the genus Massilia with the highest scores belonging to two isolates of M. timonae. To verify these results, phenotypic tests were also performed (Table 2). Phenotypic characteristics were in complete agreement with those reported for M. timonae (La Scola et al., 1998) and hence confirmed the 16S rDNA analysis results.
| Table 2: | Characteristics of M. timonae isolate U2 |
 | |
+: Present, -: Absent | |
 | |
| Fig. 2: | Effect of temperature on chitinolytic enzyme production by M. timonae isolate U2. Each point represents the mean of three independent experiments and error bars indicate SD |
Effects of cultural conditions on chitinolytic enzyme production: The optimum growth temperature for chitinase production by M. timonae was found to be in the range of 25-30°C (Fig. 2). The activity achieved at 20°C was 90% lower than the optimum temperature. The enzyme activity decreased as the temperature increased above 30°C and even at 35°C a 60% decrease in chitinolytic activity was observed. The result of influence of initial pH on chitinase production is shown in Fig. 3. The optimum pH was in the range of 6.0 (9.2 U mL-1) to 6.5 (10.1 U mL-1). At pH 5.5 (5.1 U mL-1) and 7.0 (4.7 U mL-1) the production of chitinase decreased by more than 45 % compared to the optimum level. Chitinase production by some soil bacteria e.g., Bacillus laterosporous (Shanmugaiah et al., 2008) appears to be less sensitive to temperature variations. The same can be said for some bacteria isolated from marine habitats including Aeromonas (Jami Al Ahmadi et al., 2008a). Temperature and pH affect biological processes through several mechanisms including enzyme induction or suppression, protein denaturation and altering cell viability (Nampoothiri et al., 2004). Hence, bacteria isolated from similar habitats may show different patterns of enzyme production.
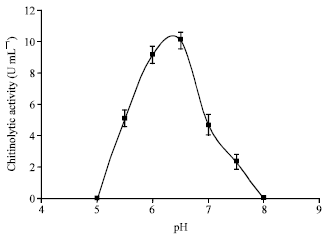 | |
| Fig. 3: | Effect of initial pH on chitinolytic enzyme production by M. timonae isolate U2. Each point represents the mean of three independent experiments and error bars indicate SD |
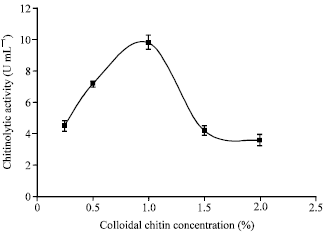 | |
| Fig. 4: | Effect of colloidal chitin concentration on chitinolytic enzyme production by M. timonae isolate U2. Each point represents the mean of three independent experiments and error bars indicate SD |
Effects of medium composition on chitinolytic enzyme production: Five different colloidal chitin concentrations ranging from 0.25 to 2% (w/v) were examined for chitinase induction. As shown in Fig. 4, M. timonae cells were very sensitive to colloidal chitin concentrations. Maximum enzyme yield (9.9 U mL-1) was achieved at a colloidal chitin concentration of 1%. At higher or lower chitin concentrations chitinolytic activity decreased significantly (p<0.001 for all concentrations when compared to chitin concentration of 1%). At a colloidal chitin concentration of 2%, chitinolytic activity (3.6 U mL-1) decreased to less than 34% of its optimal value. Microbial chitinases are inducible enzymes which are stimulated by chitin, chitooligosaccharides, chitobiose and/or GlcNAc (Cohen-Kupeic and Chet, 1998; Patil et al., 2000; Bachattacharya et al., 2007).
| Table 3: | Effect of nitrogen and carbon sources on chitinolytic enzyme production by M. timonae isolate U2 |
 | |
Data are expressed as Mean±SD | |
The molecular mechanism of chitinase induction in bacteria has been studied by cloning the chitinase gene from Streptomyces lividans in Escherichia coli (Dahiya et al., 2005). It has been suggested that for most microorganisms the optimum chitin concentration for chitinase induction is in the range of 1-2% (w/v) (Sandhya et al., 2004). This is in agreement with our results for M. timonae.
The effect of various nitrogen and carbon sources on chitinase production is shown in Table 3. The addition of casein, peptone, tryptone and yeast extract (0.1%, w/v) had no significant effect on chitinase production by M. timonae (p-value for the model = 0.163). In contrast to this result, yeast extract has been reported to enhance chitinase production in some bacterial and fungal species including Beauveria bassiana, Serratia marcescens, Streptomyces cinereoruber, Alcaligenes xylosoxydans and Trichoderma harzianum (Monreal and Reese, 1969; Sherief et al., 1991; Vaidya et al., 2001; Nampoothiri et al., 2004). According to Nampoothiri et al. (2004) this may be due to the presence of chitin or growth factors in yeast extract. However, as stated above, such low concentrations of chitin fail to induce chitinase production in most microorganisms. In addition, some other nitrogen sources including peptone and urea are reported to enhance chitinase production by A. xylosoxydans and Myrothecium verrucaria (Vaidya et al., 2001), indicating that other mechanisms are involved. Fructose, GlcNAc, glucose, lactose and maltose slightly decreased chitinase production by M. timonae (p<0.05). On the other hand, the addition of arabinose to the culture medium had no significant effect on enzyme production (p = 0.984). Although at suboptimal chitin concentrations the addition of different carbon sources to the culture medium is reported to increase chitinase production by some microorganisms including T. harzianum, at optimal concentrations using chitin as the sole carbon source results in higher yields compared to other media containing additional carbon sources in most cases (Vaidya et al., 2001; Sandhya et al., 2004; Nawani and Kapadnis, 2005).
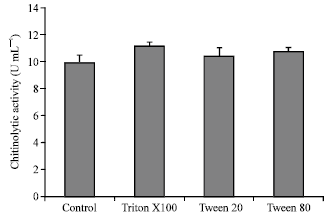 | |
| Fig. 5: | Effect of detergents on chitinolytic enzyme production by M. timonae isolate U2. Each bar represents the mean of three independent experiments and error bars indicate SD |
The only exception is arabinose which is able to enhance chitinase production by Streptomyces viridificans and A. xylosoxydans. It is of interest to note that among different carbon sources tested, only arabinose showed no inhibitory effect on enzyme production by M. timonae. However, more studies are required to establish whether this results from the combined effect of enzyme induction at the molecular level (Vaidya et al., 2001) and chitinase inhibition by the presence of a readily utilizable nutrient, or it simply indicates the absence of any direct relationship between arabinose and chitinase production.
The influence of the addition of various detergents to the basic culture medium is shown in Fig. 5. Triton X100 (0.05%, w/v) had a positive effect on chitinase production, resulting in a 12.4% increase (p = 0.016), while Tween 20 and Tween 80 showed no statistically significant effect (p = 0.261 and 0.078, respectively). It has been proposed that detergents enhance enzyme secretion by increasing cell membrane permeability (Chellapandi and Himanshu, 2008). This probably explains the increase in chitinolytic activity following the addition of detergents into the culture medium of M. timonae and other microorganisms including fungi such as A. xylosoxydans (Vaidya et al., 2001).
Physico-chemical characteristics of chitinase: At temperatures lower than 50°C, no significant activity loss was observed (p>0.05). However, at 60°C chitinolytic activity decreased to ca. 4% after 1 h (Fig. 6).
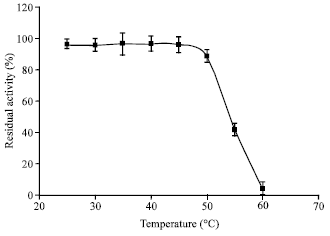 | |
| Fig. 6: | Stability of M. timonae chitinase against temperature. Each point represents the mean of three independent experiments and error bars indicate SD |
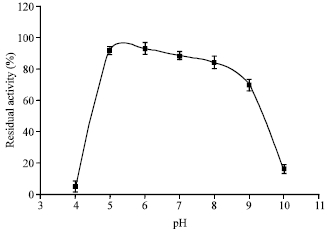 | |
| Fig. 7: | Stability of M. timonae chitinase against pH. Each point represents the mean of three independent experiments and error bars indicate SD |
Determination of pH stability (Fig. 7) indicated that the enzyme was stable between pH 5 to 7 (p>0.05).
CONCLUSION
In this study, it was demonstrated that M. timonae is able to produce chitinase when cultured in a relatively simple chitin-containing medium without requiring other external carbon and nitrogen sources. Although there are some reports describing the presence of enzymes such as proteases (Wery et al., 2003) and enzymes involved in the metabolism of N-acyl homoserine lactones (D’Angelo-Picard et al., 2005) in members of the genus Massilia, very little is known about the enzymatic capabilities of this genus due to the fact that it is a newly established genus. Further studies on this topic would be useful not only to better characterize this genus, but also, probably, to identify enzymes with potential industrial applications.
ACKNOWLEDGMENTS
This study was supported by grants No. 87-02-33-7162 (molecular identification of isolate U2 by 16S rDNA analysis) from Research Council of Tehran University and No. 425/386 (Screening and optimization) from Pharmaceutical Sciences Research Center, Tehran University of Medical Sciences, Tehran, Iran.
REFERENCES
- Bhattacharya, D., A. Nagpure and R.K. Gupta, 2007. Bacterial chitinases: Properties and potential. Crit. Rev. Biotechnol., 27: 21-28.
CrossRefDirect Link - Chellapandi, P. and M.J. Himanshu, 2008. Production of endoglucanase by the native strains of Streptomyces isolates in submerged fermentation. Braz. J. Microbiol., 39: 122-127.
CrossRefDirect Link - Dahiya, N., R. Tewari and G.S. Hoondal, 2005. Biotechnological aspects of chitinolytic enzymes: A review. Applied Microb. Biotechnol., 71: 773-782.
Direct Link - Driss, F., M. Kallassy-Awad, N. Zouari and S. Jaoua, 2005. Molecular characterization of a novel chitinase from Bacillus thuringiensis subsp. Kurstaki. J. Applied Microbiol., 99: 945-953.
CrossRef - Duo-Chuan, L., 2006. Review of fungal chitinases. Mycopathologia, 161: 345-360.
CrossRefPubMedDirect Link - Furushita, M., T. Shiba, T. Maeda, M. Yahata and A. Kaneoka et al., 2003. Similarity of tetracycline resistance genes isolated from fish farm bacteria to those from clinical isolates. Applied Environ. Microbiol., 69: 5336-5342.
Direct Link - Gallego, V., C. Sanchez-Porro, M.T. Garcia and A. Ventosa, 2006. Massilia aurea sp. nov., isolated from drinking water. Int. J. Syst. Evolut. Microbiol., 56: 2449-2453.
CrossRef - Al Ahmadi, K.J., M.T. Yazdi, M.F. Najafi, A.R. Shahverdi, M.A. Faramarzi, G. Zarrini and J. Behravan, 2008. Optimization of medium and cultivation conditions for chitinase production by the newly isolated: Aeromonas sp. Biotechnology, 7: 266-272.
CrossRefDirect Link - Jami Al Ahmadi, K., M. Tabatabaei Yazdi, M. Fathi Najafi, A.R. Shahverdi, M.A. Faramarzi, Gh. Zarrini and J. Behravan, 2008. Isolation and characterization of a chitinolytic enzyme-producing microorganism, Paenibacillus chitinolyticus JK2 from Iran. Res. J. Microbiol., 3: 395-404.
CrossRefDirect Link - La Scola, B., R.J. Birtles, M. Mallet and D. Raoult, 1998. Massilia timonae gen. nov., sp. nov., isolated from blood of an immunocompromised patient with cerebellar lesions. J. Clin. Microbiol., 36: 2847-2852.
Direct Link - Monreal, J. and E.T. Reese, 1969. The chitinase of Serratia marcescens. Can. J. Microbiol., 15: 689-696.
CrossRefPubMedDirect Link - Nampoothiri, K.M., T.V. Baiju, C. Sandhya, A. Sabu, G. Szakacs and A. Pandey, 2004. Process optimization for antifungal chitinase production by Trichoderma harzianum. Process Biochem., 39: 1583-1590.
CrossRef - Nawani, N.N. and B.P. Kapadnis, 2005. Optimization of chitinase production using statistics based experimental designs. Process Biochem., 40: 651-660.
CrossRefDirect Link - Patil, R.S., V. Ghormade and M.V. Deshpande, 2000. Chitinolytic enzymes: An exploration. Enzyme Microbiol. Technol., 26: 473-483.
CrossRefDirect Link - Roberts, W.K. and C.P. Selitrennikoff, 1988. Plant and bacterial chitinases differ in antifungal activity. J. Gen. Microbiol., 134: 169-176.
CrossRefDirect Link - Sandhya, C., L.K. Adapa, K.M. Nampoothiri, P. Binod, G. Szakacs and A. Padney, 2004. Extracellular chitinase production by Trichoderma harzianum in submerged fermentation. J. Basic Microbiol., 44: 49-58.
Direct Link - Shanmugaiah, V., N. Mathivanan, N. Balasubramanian and P.T. Manoharan, 2008. Optimization of cultural conditions for production of chitinase by Bacillus laterosporous MML2270 isolated from rice rhizosphere soil. Afr. J. Biotechnol., 7: 2562-2568.
Direct Link - Sherief, A.A., M.M.A. El-Sawah and M.A. Abd El-Naby, 1991. Some properties of chitinase produced by a potent Aspergillus carneus strain. Applied Microbiol. Biotechnol., 35: 228-230.
Direct Link - Vaidya, R.J., I.M. Shah, P.R. Vyas and H.S. Chhatpar, 2001. Production of chitinase and its optimization from a novel isolate Alcaligenes xylosoxydans: Potential in antifungal biocontrol. World J. Microbiol. Biotech., 17: 691-696.
CrossRefDirect Link - Wen, C.M., C.S. Tseng, C.Y. Cheng and Y.K. Li, 2002. Purification, characterization and cloning of a chitinase from Bacillus sp. NCTU2. Biotechnol. Applied Biochem., 35: 213-219.
PubMed - Wiwat, C., P. Siwayaprahm and A. Bhumiratana, 1999. Purification and characterization of chitinase from Bacillus circulans No. 4.1. Curr. Microbiol., 39: 134-140.
CrossRef - Zhang, Y., W. Li, K. Zhang, X. Tian, Y. Jiang, L. Xu, C. Jiang and R. Lai, 2006. Massilia dura sp. nov, Massilia albidiflava sp. nov, Massilia plicata sp. nov. and Massilia lutea sp. nov, isolated from soils in china. Int. J. Syst. Evolut. Microbiol., 56: 459-463.
CrossRef - D'Angelo-Picard, C., D. Faure, I. Penot and Y. Dessaux, 2005. Diversity of N-acyl homoserine lactone-producing and -degrading bacteria in soil and tobacco rhizosphere. Environ. Microbiol., 7: 1796-1808.
CrossRefPubMedDirect Link - Wery, N., U. Gerike, A. Sharman, J.B. Chaudhuri, D.W. Hough and M.J. Danson, 2003. Use of a packed-column bioreactor for isolation of diverse protease-producing bacteria from antarctic soil. Applied Environ. Microbiol., 69: 1457-1464.
CrossRefPubMedDirect Link
Sargar Pavankumar Sopana Reply
Thanks for become avalable this amazing knowledge.