
Behrooz Darbani
Department of Biotechnology, University of Rabe Rashidi, Tabriz, Iran
Safar Farajnia
Biotechnology Research Center, Drug Applied Research Center, Tabriz University of Medical Science, Tabriz, Iran
Mahmoud Toorchi
Department of Agronomy and Plant Breeding, Faculty of Agriculture, Tabriz University, Tabriz, Iran
Saeed Zakerbostanabad
Department of Biotechnology, University of Rabe Rashidi, Tabriz, Iran
Shahin Noeparvar
Agricultural Biotechnology Research Institute for Northwest and West of Iran, Tabriz, Iran
C. Neal Stewart Jr.
Department of Plant Sciences, University of Tennessee, Knoxville, TN, USA
Biotechnology
Year: 2008 | Volume: 7 | Issue: 3 | Page No.: 385-402







ABSTRACT
Since the 1980s, diverse methods for plant transformation have been described including biological, chemical and physical based methods. Transformation is performed to introduce novel traits, study basic biological processes, or produce recombinant proteins of interest. We review Agrobacterium-mediated transformation as well as non-biological based approaches for the production of transgenic plants. This review presents the methods of gene transfer into plants, applications, advantages and disadvantages of each method.
PDF Abstract XML References Citation
How to cite this article
Behrooz Darbani, Safar Farajnia, Mahmoud Toorchi, Saeed Zakerbostanabad, Shahin Noeparvar and C. Neal Stewart Jr., 2008. DNA-Delivery Methods to Produce Transgenic Plants. Biotechnology, 7: 385-402.
DOI: 10.3923/biotech.2008.385.402
URL: https://scialert.net/abstract/?doi=biotech.2008.385.402
DOI: 10.3923/biotech.2008.385.402
URL: https://scialert.net/abstract/?doi=biotech.2008.385.402
INTRODUCTION
The second green revolution needed for the production of enough food for the world will likely include plant biotechnology. At first glance, molecular biology is now an old technology, but its power has recently been realized and it continues to evolve. The first production of recombinant DNA molecules with the use of biochemical scissors called restriction enzymes (Smith and Wilcox, 1970; Meselson and Yuan, 1968) occurred in the early 1970s and subsequently, the first stably transformed plants were reported in the early 1980s (Herrera-Estrella, 1983). After this engineered herbicide-tolerant varieties in 1987 were allowed to be field planted in the US by the US Department of Agriculture Animal and Plant Health Inspection Service (USDA APHIS; Comai et al., 1985). Presently, transgenic crops are increasingly becoming a dominant feature of the agricultural landscapes of the USA and other countries. The estimated global area for 2003 was 67.7 million ha and for 2005 was 90 million ha. Since the introduction of Genetic Modified (GM) crops in 1996, the average growth in area has been more than 10% per year. This growth represents one of the highest adoption rates for any crop technology (Dunwell, 2004; James, 2005).
While transgenic plants have transformed agriculture, there are even greater potential impacts in non-agricultural uses. Transgenic crops to produce medically useful products might represent the foundations of a new pharming (pharmaceutical farming) industry. Plants have many advantages compared with traditional systems for the molecular farming of pharmaceutical proteins (Teli and Timko, 2004). These include the low cost of production, rapid scalability, the absence of human pathogens and the ability to fold and assemble complex proteins accurately (Ma et al., 2003). At present, transgenic plants and plant cell culture systems are becoming widely accepted as a general platform for the large-scale production of recombinant proteins like plantbodies and plantigens (Fischer et al., 2004; Artsaenko et al., 1998). Potato is one of the candidate plants as a bioreactor for recombinant protein production and is also one of the potential crops for production of the edible vaccine for oral virus immunization (Richter et al., 2000). Furthermore, transgenic plants are widely used in basic plant biology. All the above factors drive the continued evolution of transgenic plant technologies.
There are two requisite biotechnologies for the production of transgenic plants. First is the stable introduction of new genetic material into plant cells (transformation) which is the main focus of this review. Second is utilization of tissue culture methods to regenerate transgenic plants from single transformed cells. Gene delivery systems involve the use of several techniques for transfer of isolated genetic materials into a viable host cell. At present, there are two classes of delivery systems: biological and non-biological (which the latter includes physical and chemical) systems. The process of transgenic plant production is a complex multi-stage procedure involving three discrete phases as follows: 1. Exogenous DNA is delivered the host cell; 2. Exogenous DNA is stably integrated into the host cell`s genome in a manner that will result in proper expression of the gene; and 3. A viable transgenic plant is recovered. In this review, we will discuss various transformation methods and their applications to develop transgenic plants. This research represents an update of other reviews on subtopics of this study (Stanton, 2003; De la Riva et al., 1998; Rommens et al., 2004; Li et al., 1993; Racoczy-Trijanowska, 2002; Lazzeri and Shewry, 1994).
BIOLOGICAL GENE TRANSFER
Biologically-based transformation systems are biomimetic, relying on natural processes. However, plant biotechnologists have turned often negative biological processes (at least for the host cell); e.g., infection and mutagenesis, into positive forces that are intended on increasing the fitness of the host.
Agrobacterium-mediated transformation: In the 1970s the prospect of using Agrobacterium tumefaciens for the rational gene transfer of exogenous DNA into crops was revolutionary. Genetic transformation of plants was viewed as a prospect. In retrospect, Agrobacterium was the logical and natural transformation candidate to consider since it naturally transfers DNA (T-DNA) located on the tumor inducing (Ti) plasmid into the nucleus of plant cells and stably incorporated the DNA into the plant genome (Chilton et al., 1977). Now, some thirty years later, this method has been the most widely-used and powerful technique for the production of transgenic plants. There still remain, however, many challenges for genotype-independent transformation of many economically important crop species, as well as forest species (Stanton, 2003; De la Riva et al., 1998).
The direct cloning (De Frammond et al., 1983; Hoekema et al., 1983) of cassettes into T-DNA, which are subsequently transferred into the plant genome with the vir genes residing are on a separate helper plasmid was proved to be more amenable and stable. This binary system uses smaller binary vectors (about l0-15 kb) and does not require a co-integration step. Also, this system could be used to transfer many genes at once-up to 150 kb (Hamilton, 1997; Hamilton et al., 1996, 1999; Rossi et al., 1996; Liu et al., 1999).
Despite the development other non-biological methods of plant transformation (Shillito et al., 1985; Potrykus, 1991; Uchimiya et al., 1986; De la Pena et al., 1987; Arencibia et al., 1992, 1995; Fromm et al., 1985, 1986; Sanford, 1988), Agrobacterium-mediated transformation remains popular and among the most effective. This is especially true among most dicotyledonous plants, where Agrobacterium is naturally infectious. Agrobacterium-mediated gene transfer into monocotyledonous plants was thought to be not possible. However, reproducible and efficient methodologies were established for rice (Hiei et al., 1994), banana (May et al., 1995), corn (Ishida et al., 1996), wheat (Cheng et al., 1997), sugarcane (Enriquez-Obregon et al., 1997, 1998; Arencibia et al., 1998), forage grasses such as Italian ryegrass (Lolium multiflorum) and tall fescue (Festuca arundinacea) (Bettany et al., 2003). Among the commercially important conifers, hybrid larch was the first to be stably transformed via co-cultivation of embryogenic tissue with A. tumefaciens (Levee et al., 1997). Subsequently, this method was successfully applied to several species of spruce (Klimaszewska et al., 2001; Arce-Johnson et al., 2002; Charity et al., 2005; Grant et al., 2004).
Methods relative to transformation targets can be classified into two categories: those requiring tissue culture and in-planta methods. Tissue culture based methods can lead to unwanted genetic changes such as alterations in cytosine methylation, induction of point mutations and various chromosomal aberrations (Phillips et al., 1994; Singh, 2003; Clough, 2004). In-planta methods obviate the need for tissue culture. Feldmann and Marks (1987) reported the successful genetically transformed Arabidopsis thaliana by inoculating seed with A. tumefaciens. Transformation rates greatly improved when Bechtold et al. (1993) inoculated plants that were at the flowering stage. At present, there are very few species that can be routinely transformed in the absence of a tissue culture-based regeneration system. A. thaliana can be transformed by several in-planta methods including vacuum infiltration (Clough and Bent, 1998), transformation of germinating seeds (Feldmann and Marks, 1987) and floral dipping (Clough and Bent, 1998). Other plants that were successfully subjected by vacuum infiltration include rapeseed, Brassica campestris and radish, Raphanus sativus (Ian and Hong, 2001; Desfeux et al., 2000). The labor-intensive vacuum infiltration process was eliminated in favor of simple dipping of developing floral tissues (Clough and Bent, 1998). Also, the results indicate that the floral spray method of Agrobacterium can achieve rates of in-planta transformation comparable to the vacuum-infiltration and floral dip methods (Chung et al., 2000). In-planta procedures have some advantages to tissue culture methods. They are extremely simple, tissue culture and the resulting somaclonal variations are avoided and only a short time is required in order to obtain entire transformed individuals. However, the mean frequency of transformants in the progeny of such inoculated plants is relatively low and very variable. In many species false positive results are apparently the case leading to the conclusion that this method might be effective when in reality it is not.
However, recently, Agrobacterium inoculation of germinating seeds of rice has resulted in transformation efficiencies higher than 40% (Supartana et al., 2005) but ranged from 4.7 to 76% for the flower infiltration method and from 2.9 to 27.6% for the seedling infiltration method (Trieu et al., 2000). In contrast with A. tumefaciens, most plant cells transformed with A. rhizogenes develop into transgenic hairy roots that are easily recognized and cultivated in vitro. A. rhizogenes mediated transformation is useful for recalcitrant species (Tepfer, 1984; Mugnier, 1988; Petit et al., 1987) as well as for many areas include for artificial seed production (Uozumi and Kobayashi, 1997), for production of plant secondary metabolites (Hamill and Lidgett, 1997), as an experimental system to study biochemical pathways (Braun et al., 2002), to study of responses to chemicals and to study interactions with other organisms (Downs et al., 1994; Mugnier, 1997; Kifle et al., 1999). In addition, hairy roots have been used to express antibodies and other recombinant proteins (Wongsamuth and Doran, 1997; Medina-Bolívar and Cramer, 2004).
For inoculation of manually impossible infection of viruses a method was developed that called agroinoculation (Grimsley, 1990). Furthermore, agroinoculation almost always increases infection efficiency even when manual inoculation is possible (Hayes et al., 1988; Briddon et al., 1989; Lesser et al., 1992). One of the most efficient applications for agroinoculation is induction of recombination which it is able for marker gene deletion (Jia et al., 2006).
The optimization of Agrobacterium-plant interaction is crucial for efficient transformation. Many factors (e.g., the type of explant) are also important and they must be suitable to allow the recovery of whole transgenic plants (De la Riva et al., 1998; Jefferson, 1987; Opabode, 2006; Hiei et al., 1994; Frame et al., 2002; Zhao et al., 2001; Cardoza and Stewart, 2003; Zhang et al., 2003; Arencibia et al., 1998; Enriquez-Obregon et al., 1999; Olhoft and Somers, 2001; Cheng et al., 1997, 2003; Desfeux et al., 2000; Jones et al., 2005). In addition, residual Agrobacterium can confound transgenic plant analysis and also be of environmental concern (Cubero and Lopez, 2004). One can test for contaminating Agrobacterium by culturing (Cubero, 1998; Cubero et al., 2006), or Polymerase Chain Reaction (PCR). Being of the viable but nonculturable (VBNC) state for the Agrobacterium (Manahan and Steck, 1997) and false positive results of PCR advise a combination of isolation and molecular techniques (Moore et al., 2001).
It is notable that the fundamental limitation of Agrobacterium based systems led to the reduced ability of them to transfer DNA into monocot cells. Also, this system relies completely on the functioning of the Vir proteins, which target only to the nuclear genome and not to organelle genomes. Public concerns about the permanent introduction of foreign DNA into food crops in order to produce the safe transformants has recently been caused to use plant-derived putative transfer DNAs (P-DNA) fragment which it can be used to replace the universally employed Agrobacterium T-DNA (Rommens et al., 2004; Depicker et al., 2001; Rommens, 2004, Darbani et al., 2007, 2008).
Viral based transformation: A number of biological alternative strategies have been developed over the last few years for gene delivery in higher plants, but none have led to stable plant transformation. Viral based transformation systems provide very high expression temporarily in plant cells. Potato virus X (Ruiz et al., 1998), barley stripe mosaic virus (Holzberg et al., 2002) and geminivirus vectors (Kjemtrup et al., 1998; Turnage et al., 2002) are some of plant viral vectors that are commonly used. Substrates of viral mediated transformation could be viral RNA or more conveniently, viral cDNA which can be used via direct inoculation, which has proven to be the most efficient way of introducing cDNA-derived viral RNA into plants (Ahlquist et al., 1984; Lu et al., 2003; Marillonnet et al., 2005; Jia et al., 2003). Modified viruses in which an antibiotic resistance gene replaces the viral coat protein genes, can be used to enhance integration (Elmer and Rogers, 1990). Rapidity and high level expression of transgene are the benefits of virus-based transient RNA and DNA replicons versus non-viral systems. Also, the potential of vectors based on viral genomes to spread throughout a host plant to produce systemic infection has inspired the potential combination of using viral vectors with in-planta based transformation. But they have low infectivity using average or large-length transgenes (Porta and Lomonossoff, 2002; Marillonnet et al., 2005). Other disadvantageous include large number integrated transgene copies, chromosomal rearrangement, appearance of disease symptoms and low frequency for stable transformation. Viral vectors can also serve as efficient initiators of Posttranscriptional Gene Silencing (PTGS) or Virus Induced Gene Silencing (VIGS) in natural virus infections (Tenllado and Diaz-Ruiz, 2001). Also, inducible expression is applicable by viral based transformation methods, which could be useful tool for marker gene elimination (Hull et al., 2005; Kopertekh and Schiemann, 2004; Jia et al., 2006).
NON-BIOLOGICAL BASED TRANSFORMATION
Non-biological based or direct gene transfer methods use chemical or physical DNA up-take induction into protoplasts, intact cells or whole plants. These methods are potentially suitable for all species and all plant materials and do not require to specialized vector systems. Often, until Agrobacterium-mediated transformation systems are optimized for plant species, direct DNA techniques are used for transformation. Direct transformation systems have the following disadvantages: requirement of specialized instrumentation and consumables in many cases, fragmentation and rearrangement of introduced sequences, high copy integration and co-integration of target gene cassette with plasmid vector sequence. As regards nucleus targeting of Agrobacterium-mediated transformation, however, gene targeting into other intracellular compartment, e.g., chloroplast genetic transformation is feasible via direct gene transfer methods. Targeting into the chloroplast have some advantageous, ex., high level expression, gene silencing and positional effects elimination and especially from the safety point of view (Darbani et al., 2007). Also, chloroplasts are better able to express bacterial genes than are nuclei. Therefore, worthy is clear for these methods in which will be discussed in this section.
Particle bombardment/biolistics: As an alternative to protoplast transformation, particle bombardment was first described as a method for the production of transgenic plants in 1987 (Sanford et al., 1987) especially for transformation of more recalcitrant cereals. John Sanford, Ted Klein and colleagues at Cornell University advanced the original bombardment concept (Sanford et al., 1987; Sanford, 2000) and coined ‘biolistics`. A variation in the microprojectile transformation is an electric Discharge Particle Acceleration Device (DPAD). Unique advantages of this methodology compared to alternative propulsion technologies are discussed in terms of the range of species and genotypes that have been engineered and the high transformation frequencies for major agronomic crops (McCabe and Christou, 1993). In plant research, the major applications of biolistics include transient gene expression studies, production of transgenic plants and inoculation of plants with viral pathogens (Sanford, 2000; Southgate et al., 1995; Taylor and Fauquet, 2002). Because of the low cost, more heterogeneous tungsten particles are also widely consumed as a physical factor but transformation efficiency is highest with gold particles in the range of 0.7-1.0 μm mean diameter (Southgate et al., 1995; Taylor and Fauquet, 2002; Sanford et al., 1993; Kikkert, 1993; Sanford et al., 1991).
Gene constructs for biolistics can be in the form of circular or linear plasmids or a linear expression cassette. Embryogenic cell cultures are likely the best explant to use for biolistic transformation because they can be spread out as uniform targets of cells and because they have a high recovery capacity (Kikkert et al., 2004). Use of mannitol or sorbitol as osmotica in bombardment medium is effective and caused higher rates of stable transformants for all suspension cultured cells (Kikkert et al., 2004), as does brief air-drying. Rice transformation has also been successfully achieved via the microbombardment of immature embryos or embryogenic calli (Li et al., 1993; Sivamani et al., 1996; Cao et al., 1992; Zhang et al., 1996), in which transformation efficiency has been raised to 50% (Li et al., 1993). Particle bombardment has emerged as a reproducible method for wheat transformation (De Block et al., 1997; Bliffeld et al., 1999) and the first stable transformation in a commercially important conifer species (Picea glauca) was achieved via embryogenic callus tissue as explant (Ellis et al., 1993) which it adapted for transformation of other species of spruce (Charest et al., 1996; Walter et al., 1999), larch (Klimaszewska et al., 1997) as well as radiata (Walter et al., 1998) and loblolly pines (Connett et al., 2002). Hypersensitive responses to Agrobacterium that leads plant cell death are eliminated (Perl et al., 1996) via biolistics as well as the need to kill Agrobacterium after transformation. Operation of the biolistic device is easy. Furthermore, plasmid construction is simplified and co-transformation with multiple transgenes (Francois et al., 2002; Leech et al., 1998) is routine. The use of linear expression cassettes called clean gene technology eliminates the possible of plasmid backbone integration into the target (Fu et al., 2000). Biolistics is the method of choice for the study of transient gene expression 24-48 h after bombardment as well as for plastid transformation (Southgate et al., 1995; Bruce et al., 1989; Norris et al., 1993). Co-integrated of this system with viral vectors can enhance expression through increased copy number by self-replication and integration (Hayes et al., 1988). In situ fertilization using in vitro biolistic based targeted microspores could be useful in order to transgenic plants production without any tissue culture phase, in-planta (Aziz and Machray, 2003).
Biolistics has some disadvantages. The transformation efficiency might be lower than with Agrobacterium and it is more costly as well. Intracellular targets are random and DNA is not protected from damage. Many researchers have not used biolistics because of the high frequency for complex integration patterns and multiple copy insertions that could cause gene silencing and variation of transgene expression (Dai et al., 2001; Darbani et al., 2008). But some laboratories have overcome this problem by reducing the quantity of DNA loaded onto the microcarriers and/or by use of linear cassettes (Fu et al., 2000). Also, biolistic mediated site-specific integration of rice is not less efficient than Agrobacterium-mediated transformation because single-stranded T-DNA is not a substrate for Cre/lox recombination and poly ethylene glycol-mediated (PEG-mediated) method since it requires the use of protoplasts (Srivastava and Ow, 2001). The ability of successful integration of Yeast Artificial Chromosomes (YACs) with insert up to 150 kb (Mullen et al., 1998) and of 106 kb BAC plasmid (Ercolano et al., 2004) into the plant genome by particle bombardment is an attractive proposition, especially for functional complementation analysis.
Efforts to reduce the incidence of recombination and reduction in the amount of DNA bombarded into each cell have been led to develop ‘agrolistics`, which was developed to increase efficiency of biolistics and yield simpler integration patterns (Taylor and Fauquet, 2002). Agrolistics is a refinement to microparticle bombardment technology which vir genes are co-bombarded. Use of agrolistics has been shown to increase the number of transgenic plants that have the clean integrate or precisely transgene as well as to reduce the frequency of degraded transgene integrations (Hansen and Chilton, 1996). This technology requires further development but has the potential as a refinement to alternative of biolistics.
Electro-transfection: Electroporation is the application of strong electric field pulses to cells and tissue is known to cause some type of structural rearrangement of the cell membrane resulting in a temporary increase in porosity and providing a local driving force for ionic and molecular transport through the pores. In vitro introduction of DNA into cells is now the most common application of electroporation. Several physical factors such as created transmembrane potential by the imposing pulse electric field, extent of membrane permeation, duration of the permeated state, mode and duration of molecular flow, global and local (surface) concentrations of DNA, form of DNA, tolerance of cells to membrane permeation and the heterogeneity of the cell population (Hui, 1995; Weaver, 1995) may affect the electro-transfection efficiency.
Transient expression from electroporated plant cells has been used to research at different field (Higo and Higo, 1996; Ecker and Davis, 1986; Fisk and Dandekar, 1998). Electroporation coupled with a transient expression assay is rapid, allowing for the reproducible detection of gene products within hours of the introduction of DNA in contrast with a stable transformation (Fisk and Dandekar, 2004). Electroporation has several advantages over biolistics in that it does not require the expensive particle gun apparatus, associated consumable supplies and licensing and has worked well for stable-transformation experiments (Shewry et al., 1994). Also, the efficiency of electroporation was relatively high: 6.25% for embryos and 54.6% for callus clusters, which is fully comparable with the best results obtained for this species after microbombardment (Deshayes et al., 1985). Using protoplasts as explants, regeneration of transformants limits its use for stable transformation (Lazzeri and Shewry, 1994). Zea mays embryos are suitable explants which enables the recovery of fertile transgenic plants recovering (D`Halluin et al., 1992). However, electroporation-mediated gene transfer into intact meristems was reported by Chowrira et al. (1995). It is mentionable that use of pollen electroporation is also able to escape regeneration phase (Saunders and Matthews, 1995). Despite apparent advantages of electroporation, it is not used very often because of sometimes irreproducible results and non-Mendelian segregation of progeny.
In order to efficiently apply electric field, especially to expel the tissue culture step, electrophoresis-mediated gene transfer was developed to force negatively charged foreign DNA into the host cells (Ahokas, 1989). It was proposed as an alternative method to biolistics, but is not nearly as efficient (Ahokas, 1989; Griesbach and Hammond, 1994). A voltage of 25 mV and amperage of 0.5 mA for 15 min are the most often used parameters (Ahokas, 1989; Griesbach and Hammond, 1994; Songstad et al., 1995). On average, this technique gives about 55% survival rate which up to 57% of the survivors expressed a marker gene (Songstad et al., 1995). Comparison of this technique with Agrobacterium and particle gun delivery approaches introduced the in vivo electrophoresis transformation as an acceptable method by use of intact poinsettia meristems (Vik et al., 2001).
Silicon carbide fibers: One of the recently developed methods to delivery of DNA to plants is Silicon Carbide-Mediated Transformation (SCMT). Physical and chemical characteristics of silicon carbide fibers make them capable of puncturing the cells without killing them. The advantages of this system are: rapid, inexpensive, easy to set-up and is effective on a variety of cell types. Some of its disadvantages include, low transformation efficiency, cell damage and inhalation of the fibers can be a health hazard (Komatsu et al., 2006; Kaeppler et al., 1990, 1992; Wang et al., 1995; Songstad et al., 1995). The efficiency of SCMT depends on the fiber size, parameters of vortexing, shape of the vessels used, plant species and explant and characteristics of the plant cells and especially the thickness of the cell wall (Racoczy-Trijanowska, 2002). There are several known examples SCMT success including maize (Kaeppler et al., 1992; Wang et al., 1995; Petolino et al., 2000), rice (Komatsu et al., 2006), wheat (Brisibe et al., 2000), tobacco (Kaeppler et al., 1990), Lolium multiflorum, Lolium perenne, Festuca arundinacea and Agrostis stolonifera (Dalton et al., 1998). Recently, the efficiency of rice callus transformation based on whisker fibers was 30 to 50% (Komatsu et al., 2006). SCMT has been shown to be comparable in efficiency to biolistics in some instances and therefore, represents an alternative option for soybean embryogenic tissue transformation (Terakawa et al., 2005). Also, silicon carbide fibers can be used for wounding to improve frequency of Agrobacterium-based transformation (Singh and Chawla, 1999).
Polymer based transfection (polyfection): A barrier to more widespread use of genetic manipulation techniques is the difficulty of transforming some of the major crop plant species. Some cereal and grass species are recalcitrant to genetic manipulation. Nevertheless, because of the agronomic importance of monocots as staple food plants, there is great interest in transforming these species. Cereal transformation has been achieved by several different techniques of direct gene transfer. The first successful method was protoplast transformation (Rhodes et al., 1988) and subsequently particle bombardment (Gordon-Kamm et al., 1990), cell electroporation (D`Halluin et al., 1992) and vortexing tissues with silicon carbide whiskers (Wang et al., 1995) have all resulted in transgenic cereal plants.
The most common method of delivering foreign DNA into plant protoplasts involves treatment with PEG as a hydrophilic long-chain polymer (Paszkowski et al., 1984). Protoplast transformation has several advantages. Many cells can be handled with ease, many independent transformants can be produced, the selection of transformants is relatively simple and no specialized equipment is needed. As mentioned earlier, regenerating whole plants from transgenic protoplasts has proven difficult (Lazzeri and Shewry, 1994; Golovkin et al., 1993; Shimamoto et al., 1989). The system is only rarely employed due to the low frequency of transformants and the inability of many species to be regenerated into whole plants from protoplasts (Paszkowski et al., 1984). However, it has been reported that the PEG based method is efficient: 10-20% for Physcomitrella transformation (Schaefer, 2002) and 65 to 28% for rice protoplasts (Bart et al., 2006) according to plasmid size (4.5 to 12 kb). Recently, a fast PEG-mediated transient expression system was reported that is suitable for production of biopharmaceuticals (Buar et al., 2005). In addition, long-chain cations such as poly-L-ornithine, poly-L-lysine, dextran sulphate and high concentrations of divalent ions such as Zn2+ and Cu2+ have been applied to achieve high levels of DNA uptake (Potrykus, 1991). Among the polycations combined polybrene-spermidine based callus treatment was introduced as an efficient method for cotton transformation which it is less toxic than the other polycations and spermidine protects DNA from shearing because of its condensation effect (Sawahel, 2001).
Liposome-mediated transfection (lipofection): A derivation of PEG-mediated transformation is the liposome-mediated transformation technique. Cationic liposomes are positively charged lipids and are increasingly used for DNA uptake due to their favorable interactions with negatively charged DNA and cell membranes. In this approach foreign DNA must be encapsulated in a spherical lipid bilayer termed a liposome to prepare lipoplexes (Gad et al., 1990). After endocytosis (Fukunaga et al., 1983), the DNA is free to recombine and integrate into the host genome. As with other transformation systems, a variety of vectors including viral vectors can be applied with this system. Successful transformation based on this system was reported for tobacco (Dekeyser et al., 1990), wheat (Zhu et al., 1993) and potato (Sawahel, 2002). In the recent case frequency of stable transformation was 7% of calli regenerating from protoplasts. This method is relatively non-toxic (Antonelli and Stadler, 1989), is simple to perform with readily available chemical reagents, is highly reproducible and efficient and requires no sophisticated equipment (Antonelli and Stadler, 1990; Felgner et al., 1987). Transformation of intact YACs into plant cell was achieved via lipofection-like particle bombardment. A lipofection-PEG combination method was more efficient than each one of them separately (Wordragen et al., 1997; Freeman et al., 1984). It is determined lipoplex size is a major factor determining lipofection efficiency which large lipoplex particles showed, in general, higher lipofection efficiency than small particles (Almofti et al., 2003).
Injection based methods:
Microinjection is one of the most effective techniques of transforming animal cells and has been used for the transformation of plant cells as well (Banks and Evans, 1976). An important feature of the method is the use of Low Melting Point (LMP) agarose, both for holding protoplasts during microinjection and for their subsequent culture (Harwood and Davies, 1990). This direct and precise delivery of DNA into the plant cell or its nucleus is achievable by means of a glass microcapillary-injection pipette (Crossway et al., 1986; Morikawa and Yamada, 1985). The procedure is very slow and requires an expensive micromanipulator. However, it allows the introduction not only of plasmids but also whole chromosomes into plant cells (Griesbach, 1987; Jones-Villeneuve et al., 1995). Several plant species such as tobacco (Schnorf et al., 1991), petunia (Griesbach, 1987), oilseed rape (Neuhaus et al., 1987) and barley (Holm et al., 2000) have been transformed using microinjection. Also, the technique has been used to study intercellular communication and macromolecular trafficking in plants (Storm et al., 1998). This technique is very precise from the delivery point of view, is capable for transformation of any protoplast or single-cell system and has extremely high transformation efficiencies, but requires expensive equipment and tedious procedures.
The injection of inheritable materials using a hypodermic syringe is called macroinjection. Macroinjection is an approach to the delivery of exogenous DNA that could circumvent the expense, time and genome stress associated with the transformation of cells in culture and subsequent plant regeneration. Also, plant regeneration from transformed protoplasts, still remains a problem. Therefore, cultured tissues, which facilitate the development of immature structures, provides an alternate cellular target for transformation. These immature structures may include immature embryos, meristems, immature pollen, germinating pollen, etc. (Soyfer, 1980; Zhou et al., 1983). The main disadvantage of this technique is the likelihood for the production of chimeric plants with only a part of the plant transformed. However, from this chimeric plant, transformed plants of single cell origin can be subsequently obtained. To escape these problems Pollen-Tube Pathway (PTP) utilizing the normal fertilization cycle was developed to eliminate the regeneration phase. PTP based transformation is an injection/delivery of naked DNA into ovaries to produce transformed progeny (Touraev et al., 1997). In this approach the stigma is cut off and a drop of DNA solution is applied to the cut end of the style of recently pollinated plant florets. This procedure was used for the first time in rice (Luo and Wa, 1988). Afterward, other species such as wheat (Mu et al., 1999), soybean (Hu and Wang, 1999; Shou et al., 2002), Petunia hybrida (Tjokrokusumo et al., 2000) and watermelon (Chen et al., 1998) were subjected. In rice, the transformation efficiency was higher after excising the palea than after cutting top floret (Xie et al., 1990). A variation is the injection of a bacterial inoculum or plasmid DNA into inflorescences with pollen mother cells in the premeiotic stage without removing the stigma. Such an approach has been employed for rye (De la Pena et al., 1987). Although the results of the described experiments could be encouraging, the final transformation efficiency was about 10-fold lower than with biolistics. However, this approach has recently been introduced as a potential method for stable plant transformation (Peffley et al., 2003).
Wave and beam mediated transformation: Ultrasound treatment increases membrane permeability (Bommannan et al., 1992), thereby facilitating the entrance of macromolecules into cells for transformation (Wyber et al., 1997). Ultrasound for transfection has been widely applied in animal cells or tissues. However, there has been little research using ultrasound in plant cells or tissues. Ultrasound has been reported to mediate gene uptake in plant protoplast, suspension cells and intact pieces of tissues. In this technique explants are suspended in a few milliliters of sonication medium in a microcentifuge tube. Plasmid DNA (and possibly carrier DNA) is then added and after rapid mixing the samples are ready for sonication. The cells are finally transferred to fresh growth medium. In this term, sound frequency and exposure time determined the uptake efficiency (Liu et al., 2005). Early papers report transient expression of chloramphenicol acetyltransferase (cat) gene was reported in sugar beet (Beta vulgaris L.) and tobacco (Nicotiana tabacum L.) via a brief exposure of the protoplasts to 20 kHz ultrasound in the presence of plasmid DNA (Joersbo and Brunstedt, 1990a). Also, ultrasound treatment can be effective to transfection of virus particles (Joersbo and Brunstedt, 1990b). Stable transformation of tobacco by leaf pieces sonication was achieved via 1500 to 2000 fold longer time ultrasound treatment compared with protoplast sonication (Zhang et al., 1991). Another example for intact tissue sonication based transformation is potato tuber discs ultrasonication (Sawahel, 1996). Much of the ultrasound technique is aimed at Sonication-Assisted Agrobacterium-mediated Transformation (SAAT) in plant cells or tissues (Trick and Finer, 1997; Horsch et al., 1985; Weber et al., 2003) as introduced earlier. SAAT is a new technology and method involves subjecting the plant tissue to brief periods of ultrasound in the presence of Agrobacterium. Experiments have been demonstrated that SAAT tremendously improved the efficiency of Agrobacterium infection by introducing large numbers of micro-wounds into the target plant cells or tissues (Santarem et al., 1998; Tang, 2003). Trick and Finer (1997) reported that, in all tissues tested, the SAAT treatment greatly enhanced the levels of transient expression. In addition, 2.2-fold, 2.5-fold and 4.1-fold increase, consequently resulted by sonication, sonication with CaCl2 treatment and sonication with acetosyringone treatment in transformation frequency in contrast to manual wounding based method (Kumar et al., 2006). Also, SAAT could be efficiently useful for transformation of woody trees, particularly Eucalyptus sp. (Monica et al., 2004). In order to in-planta based transformation via SAAT, germinating seeds and seedlings were subjected that enhanced the efficiency of transformation (González et al., 2002). Recently, laser beam was applied to introduce genetic materials. Laser-mediated transformation works by a focused laser microbeam to puncture momentarily-made self-healing holes (~ 0.5 μm) in the cell wall and membrane. Therefore, exogenous DNA could simply be taken up by cells (Badr et al., 2005). The method was newly developed and requires further assessing for both of the different experimental conditions and plant species.
Desiccation based transformation: Since dried embryos have the unique ability to take up DNA during rehydration (Topfer et al., 1989), it was hypothesized that they could be mixed with a nutrient solution containing the foreign DNA for direct transformation. DNA would be taken up as the embryo rehydrates and seedlings would germinate in the presence of a selection medium to assess the incorporation of the foreign DNA (Topfer et al., 1989). The old and immature approach is potent at least theoretically to expel many problems associated with biological or chemical and/or physical methods but it needs further surveys.
CONCLUSION
Plant genetic transformation is of particular benefit to molecular genetic studies, crop improvement and production of pharmaceutical materials. These applications and the desire for higher transformation efficiency have stimulated work on inventing and improving various methods (Table 1). Aside from the elimination of tissue culture for Arabidopsis and a few other species, no great breakthroughs on basic transformation techniques have taken place since Agrobacterium and biolistics. Agrobacterium-based methods are usually superior for many species including dicots and monocots. For some angiosperms and non-angiosperms biolistics is usually used. The others are typically not done on a routine basis (Table 1). Biolistics is by far the most widely used direct transformation procedure both experimentally in research and commercially. In addition, it has proven to be the most efficient. So why have all these other methods emerged in the past 20 years if we already have efficient transformation techniques in Agrobacterium and biolistics? Then are there all these other methods? There are two reasons. First of all, there is hope that a more efficient and less expensive method will be developed. The commercial biolistics device costs thousands of US dollars and while less expensive home-made devices can be built fairly easily, they usually do not work as well. The second and most important reason is that biolistics and Agrobacterium are patented and obtaining a commercial license is not trivial, thus, commercialization pathways might be blocked without new technology.
Chloroplast targeting is one of the advantageous for direct transformation methods in sharp contrast with Agrobacterium based methods. High level recombinant protein expression is one of advantageous for chloroplast targeting but it have some drawbacks, e.g., low transformation frequency, prolonged selection procedures under high selection pressure and green part dependency for transgene products. However, maternal inheritance transgenes is one of the other advantageous for transplastomes, which has benefits relative to biosafety. In-planta transformation methods are immensely attractive; however, this has proven to be effective and efficient in Arabidopsis. Among these methods
| Table 1: | Juxtaposition of gene delivery methods |
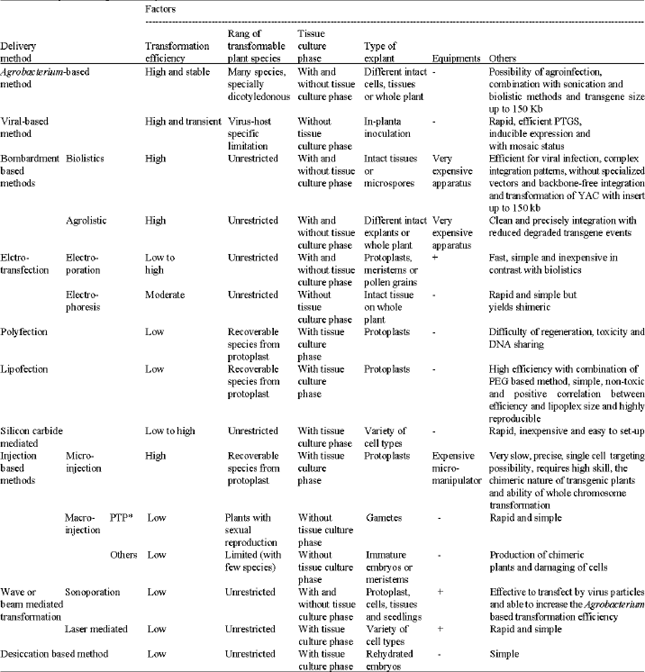 | |
| *Pollen-tube pathway, +: Necessary, -: Unnecessary | |
Agrobacterium based in-planta transformation is efficient than others especially using sonoporation, ex., seedling sonication, in order to eliminate the chimeric results. The reader should be cautious about investing many resources in techniques that are touted as effective but unproven. Even though plant transformation can no longer be described as a new technology, it continues to evolve. Moreover, tools go on to expand, leading to more facile and efficient transformation.
REFERENCES
- Ahlquist, P., R. French, M. Janda and L.S. Loesch-Fries, 1984. Multicomponent RNA plant virus infection derived from cloned viral cDNA. Proc. Natl. Acad. Sci. USA., 81: 7066-7070.
Direct Link - Ahokas, H., 1989. Transfection of germinating barley seed electrophoretically with exogenous DNA. Theor. Applied Genet., 77: 469-472.
CrossRefDirect Link - Almofti, M.R., H. Hideyoshi, S. Yasuo, A. Almofti, W. Li and K. Hiroshi, 2003. Lipoplex size determines lipofection efficiency with or without serum. Mol. Membrane Biol., 20: 35-43.
CrossRef - Antonelli, N. and J. Stadler, 1990. Genomic DNA can be used with cationic methods for highly efficient transformation of maize protoplasts. Theor. Applied Genet., 80: 395-401.
CrossRef - Arce-Johnson, P., F. Aquea, F. Cerda, M. Gebauer and C. Medina, 2002. Stable transformation of Pinus radiatai embryogenic tissue by Agrobacterium tumefaciens. Plant Cell Tissue Organ Culture, 70: 251-257.
CrossRef - Arencibia, A., P. Molina, G. de la Riva and G. Selman-Housein, 1995. Production of transgenic sugarcane (Saccharum officinarum L.) plants by intact cell electroporation. Plant Cell Rep., 14: 305-309.
CrossRefDirect Link - Arencibia, A.D., E.R. Carmona, P. Tellez, M.T. Chan and S.M. Yu et al., 1998. An efficient protocol for sugarcane (Saccharum spp. L.) transformation mediated by Agrobacterium tumefaciens. Transgenic Res., 7: 213-222.
CrossRefDirect Link - Artsaenko, O., B. Kettig, U. Fiedler, U. Conrad and K. Duering, 1998. Potato tubers as a biofactory for recombinant antibodies. Mol. Breed., 4: 313-319.
Direct Link - Aziz, N. and G.C. Machray, 2003. Efficient male germ line transformation for transgenic tobacco production without selection. Plant Mol. Biol., 51: 203-211.
CrossRef - Banks, M.S. and P.K. Evans, 1976. A comparison of the isolation and culture of mesophyll protoplasts from several Nicotiana species and their hybrids. Plant Sci. Lett., 7: 409-416.
CrossRef - Bart, R., M. Chern, C.J. Park, L. Bartley and P.C. Ronald, 2006. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods, 2: 13-13.
CrossRef - Bechtold, N., J. Ellis and G. Pelletier, 1993. In planta agrobacterium mediated gene transfer by infiltration of adult Arabidopsis thaliana plants. C. R. Acad. Sci. (Paris) Life Sci., 316: 1194-1199.
Direct Link - Bliffeld, M., J. Mundy, I. Potrykus and J. Futterer, 1999. Genetic engineering of wheat for increased resistance to powdery mildew disease. Theor. Applied Genet., 98: 1079-1086.
CrossRefDirect Link - Braun, R.H., C. Eady, M.C. Christey, M. Shaw, M. Pither-Joyce and J. McCallum, 2002. The use of hairy root cultures for the study of sulfur metabolism in plants. Microbes and Molecules. A combined meeting of the NZSBMB, NZSPP and NZMS. November 26-29, University of Canterbury: Christchurch, New Zealand.
- Briddon, R.W., J. Watts, P.G. Markham and J. Stanley, 1989. The coat protein of beet curly top virus is essential for infectivity. Virology, 172: 628-633.
PubMedDirect Link - Brisibe, E.A., A. Gajdosova, A. Olesen and S.B. Andersen, 2000. Cytodifferentiation and transformation of embryogenic callus lines derived from anther culture of wheat. J. Exp. Bot., 51: 187-196.
CrossRef - Buar, A., F. Kauffmann, H. Rolli, A. Wise and R. Leuthje et al., 2005. A fast and flexible PEG-mediated transient expression system in plants for high level expression of secreted recombinant biopharmaceuticals. J. Biotechnol., 119: 332-342.
CrossRef - Cao, J., X. Duan, D. McElroy and R. Wu, 1992. Regeneration of herbicide resistant transgenic rice plants following microprojectile-mediated transformation of suspension culture cells. Plant Cell Rep., 11: 586-591.
CrossRefDirect Link - Cardoza, V. and C.N. Stewart, 2003. Increased Agrobacterium-mediated transformation and rooting efficiencies in canola (Brassica napus L.) from hypocotyl segment explants. Plant Cell Rep., 21: 599-604.
CrossRef - Charest, P.J., Y. Devantier and D. Lachance, 1996. Stable genetic transformation of Picea mariana (black Spruce) via particle bombardment. In Vitro Cell Dev. Biol. Plant, 32: 91-99.
Direct Link - Charity, J.A., L. Holland, L. Grace and C. Walter, 2005. Consistent and stable expression of the nptII, uidA and bar genes in transgenic Pinus radiata after Agrobacterium tumefaciens-mediated transformation using nurse cultures. Plant Cell Rep., 23: 606-619.
CrossRef - Chen, W.S., C.C. Chiu, H.Y. Liu, T.L. Lee and J.T. Cheng et al., 1998. Gene transfer via pollen-tube pathway for antifusarium wilt in watermelon. Biochem. Mol. Biol. Int., 46: 1201-1209.
PubMed - Cheng, M., J.E. Fry, S.Z. Pang, H.P. Zhou and C.M. Hironaka et al., 1997. Genetic transformation of wheat mediated by Agrobacterium tumefaciens. Plant Physiol., 115: 971-980.
Direct Link - Cheng, M., T. Hu, J. Layton, C.N. Liu and J.E. Fry, 2003. Desiccation of plant tissues post-agrobacterium infection enhances T-DNA delivery and increases stable transformation efficiency in wheat. In vitro Cell Dev. Biol. Plant, 39: 595-604.
CrossRef - Chilton, M.D., M.H. Drummond, D.J. Merio, D. Sciaky, A.L. Montoya, M.P. Gordon and E.W. Nester, 1977. Stable incorporation of plasmid DNA into higher plant cells: The molecular basis of crown gall tumorigenesis. Cell, 11: 263-271.
CrossRefPubMedDirect Link - Chung, M.H., M.K. Chen and S.M. Pan, 2000. Floral spray transformation can efficiently generate arabidopsis. Transgenic Res., 9: 471-486.
CrossRef - Comai, L., D. Facciotti, W.R. Hiatt, G. Thompson, R.E. Rose and D.M. Stalker, 1985. Expression in plants of a mutant aroA gene from Salmonella typhimurium confers tolerance to glyphosate. Nature (London), 317: 741-744.
CrossRef - Crossway, A., J.W. Oakes, J.M. Irvine, B. Ward, V.C. Knauf and C.K. Shewmaker, 1986. Integration of foreign DNA following microinjection of tobacco mesophyll protoplasts. Mol. Gen. Genet., 202: 179-185.
CrossRefDirect Link - Cubero, J., B. Lastra, C.I. Salcedo, J. Piquer and M.M. López, 2006. Systemic movement of Agrobacterium tumefaciens in several plant species. J. Applied Microbiol., 101: 412-422.
CrossRef - D'Halluin, K., E. Bonne, M. Bossut, M. de Beuckeleer and J. Leemans, 1992. Transgenic maize plants by tissue electroporation. Plant Cell, 4: 1495-1505.
CrossRefDirect Link - Dai, S., P. Zheng, P. Marmey, S. Zhang and W. Tian et al., 2001. Comparative analysis of transgenic rice plants obtained by Agrobacterium-mediated transformation and particle bombardment. Mol. Breed., 7: 25-33.
CrossRefDirect Link - Dalton, S.J., A.J.E. Bettany, E. Timms and P. Morris, 1998. Transgenic plants of Lolium multiflorum, Lolium perenne, Festuca arundinacea and Agrostis stolonifera by silico carbide fibre-mediated transformation of cell suspensions. Plant Sci., 132: 31-43.
CrossRef - Darbani, B., A. Eimanifar, C.N. Stewart and W.N. Camargo, 2007. Methods to produce marker-free transgenic plants. Biotechnol. J., 2: 83-90.
PubMedDirect Link - De Block, M., D. Debrouwer and T. Moens, 1997. The development of a nuclear male sterility system in wheat. Expression of the barnase gene under the control of tapetum specific promoters. Theor. Applied Genet., 95: 125-131.
CrossRef - De Framond, A.J., K.A. Barton and M.D. Chilton, 1983. Mini-Ti: A new vector strategy for plant genetic engineering. Biotechnology, 5: 262-269.
CrossRefDirect Link - De la Pena, A., H. Lorz and J. Schell, 1987. Transgenic rye plants obtained by injecting DNA into young floral tillers. Nature, 325: 274-276.
CrossRef - De la Riva, G.A., J. Gonzalez-Cabrera, R. Vazquez-Padron and C. Ayra-Pardo, 1998. Agrobacterium tumefaciens: A natural tool for plant transformation. Elect. J. Biotechnol., 1: 118-133.
CrossRefDirect Link - Dekeyser, R.A., B. Claes, R.M.U. De Rycke, M.E. Habets, M.C. Van Montagu and A.B. Caplan, 1990. Transient gene expression in intact and organized rice tissues. Plant Cell, 2: 591-602.
PubMedDirect Link - Desfeux, C., S.J. Clough and A.F. Bent, 2000. Female reproductive tissues are the primary target of Agrobacterium-mediated transformation by the arabidopsis floral-dip method. Plant Physiol., 123: 895-904.
CrossRefDirect Link - Deshayes, A., L. Herrera-Estrella and M. Caboche, 1985. Liposome-mediated transformation of tobacco mesophyll protoplasts by an Escherichia coli plasmid. EMBO. J., 4: 2731-2737.
Direct Link - Downs,C.G., M.C. Christey, K.M. Davies, G.A. King, J.F. Seelye, B.K. Sinclair and D.G. Stevenson, 1994. Hairy roots of Brassica napus: II. Glutamine synthetase overexpression alters ammonia assimilation and the response to phosphinothricin. Plant Cell Rep., 14: 41-46.
CrossRefDirect Link - Ecker, J.R. and R.W. Davis, 1986. Inhibition of gene expression in plant cells by expression of antisense RNA. Proc. Natl. Acad. Sci. USA., 83: 5372-5376.
Direct Link - Ellis, D.D., D.E. McCabe, S. McInnis, R. Ramachandran and D.R. Russell et al., 1993. Stable transformation of Picea glauca by particle acceleration. Biotechnol., 11: 84-89.
Direct Link - Enriquez-Obregon, G.A., R.I. Vazquez-Padron, D.L. Prieto-Samsonov, M. Perez and G. Selman-Housein, 1997. Genetic transformation of sugarcane by Agrobacterium tumefaciens using antioxidant compounds. Biotecnologia Aplicada, 14: 169-174.
Direct Link - Enriquez-Obregon, G.A., R.I. Vazquez-Padron, D.L. Prieto-Samsonov, G.A. De la Riva and G. Selman-Housein, 1998. Herbicide-resistant sugarcane (Saccharum officinarum L.) plants by Agrobacterium-mediated transformation. Planta, 206: 20-27.
CrossRefDirect Link - Enriquez-Obregon, G.A., D.L. Prieto-Samsonov, A. Gustavo, M. Perez, G. Selman-Housein and R.I. Vazquez-Padron, 1999. Agrobacterium-mediated Japonica rice transformation: A procedure assisted by an antinecrotic treatment. Plant Cell Tissue Organ Cult., 59: 159-168.
CrossRefDirect Link - Ercolano, M.R., A. Ballvora, J. Paal, H.H. Steinbiss, F. Salamini and C. Gebhardt, 2004. Functional complementation analysis in potato via biolistic transformation with BAC large DNA fragments. Mol. Breed., 13: 15-22.
CrossRef - Feldmann, K.A. and M.D. Marks, 1987. Agrobacterium mediated transformation of germinating seeds of Arabidopsis thaliana: A non-tissue culture approach. Mol. Gen. Genet., 208: 1-9.
Direct Link - Felgner, P.L., T.R. Gadek, M. Holm, R. Roman and H.W. Chan et al., 1987. Lipofection: A highly efficient, lipid-mediated DNA-transfection procedure. Proc. Natl. Acad. Sci., 84: 7413-7417.
Direct Link - Fisk, H.J. and A.M. Dandekar, 1998. Nuclear localization of a foreign gene product in tobacco results in increased accumulation due to enhanced stability. Plant Sci., 133: 177-189.
CrossRefDirect Link - Frame, B.R., H. Shou, R.K. Chikwamba, Z. Zhang and C. Xiang et al., 2002. Agrobacterium tumefaciens-mediated transformation of maize embryos using a standard binary vector system. Plant Physiol., 129: 13-22.
CrossRefPubMedDirect Link - Francois, I.E.J.A., W.F. Broekaert and B.P.A. Cammue, 2002. Different approaches for multi-transgene-stacking in plants. Plant Sci., 163: 281-295.
CrossRefDirect Link - Freeman, J.P., J. Draper, M.R. Davey, E.C. Cocking, K.M.A. Gartland, K. Harding and D. Pental, 1984. A comparison of methods for plasmid delivery into plant protoplasts. Plant Cell Physiol., 25: 1353-1365.
CrossRefDirect Link - Fromm, M., L. Taylor and V. Walbot, 1985. Expression of genes transferred into monocot and dicot plant cells by electroporation. Proc. Natl. Acad. Sci. USA., 82: 5824-5828.
Direct Link - Fromm, M., L. Taylor and V. Walbot, 1986. Stable transformation of maize after gene transfer by electroporation. Nature, 319: 791-793.
CrossRefDirect Link - Fu, X., L.T. Duc, C.T. Fontana, B.B. Bong and P. Tinjuangjun et al., 2000. Linear transgene constructs lacking vector backbone sequences generate low-copy-number transgenic plants with simple integration patterns. Transgenic Res., 9: 11-19.
CrossRef - Fukunaga, Y., T. Nagata, I. Takeba, T. Kakehi and C. Matsui, 1983. An ultrastructural study of the interaction of liposomes with plant protoplasts. Exp. Cell Res., 144: 181-189.
CrossRefDirect Link - Gad, A.E., N. Rosenberg and A. Altman, 1990. Liposome-mediated gene delivery into plant cells. Physiol. Plant., 79: 177-183.
CrossRefDirect Link - Golovkin, M.V., M. Abraham, S. Morocz, S. Bottka, A. Feher and D. Dudits, 1993. Production of transgenic maize plants by direct DNA uptake into embryogenic protoplasts. Plant Sci., 90: 41-52.
CrossRefDirect Link - Gonzalez, E.R., A. de Andrade, A.L. Bertolo, G.C. Lacerda and R.T. Carneiro et al., 2002. Production of transgenic Eucalyptus grandis x E. urophylla using the sonication-assisted Agrobacterium transformation (SAAT) system. Funct. Plant Biol., 29: 97-102.
CrossRefDirect Link - Gordon-Kamm, W.J., T.M. Spencer, M.L. Mangano, T.R. Adams and R.J. Daines et al., 1990. Transformation of maize cells and regeneration of fertile transgenic plants. Plant Cell, 2: 603-618.
CrossRefPubMedDirect Link - Grant, J.E., P.A. Cooper and T.M. Dale, 2004. Transgenic Pinus radiata from Agrobacterium tumefaciens-mediated transformation of cotyledons. Plant Cell Rep., 22: 894-899.
CrossRefDirect Link - Griesbach, R.J., 1987. Chromosome-mediated transformation via microinjection. Plant Sci., 50: 69-77.
CrossRefDirect Link - Griesbach, R.J., 1994. An improved method for transforming plants through electrophoresis. Plant Sci., 102: 81-89.
CrossRefDirect Link - Hamilton, C.M., A. Frary, C. Lewis and S.D Tankskey, 1996. Stable transfer of intact high molecular weight DNA into plant chromosomes. Proc. Natl. Acad. Sci. USA., 93: 9975-9979.
PubMedDirect Link - Hamilton, C.M., 1997. A binary-BAC system for plant transformation with high-molecular-weight DNA. Gene, 200: 107-116.
CrossRefDirect Link - Hamilton, C.M., A. Frary, Y. Xu, S.D. Tanksley and H.B. Zhang, 1999. Construction of tomato genomic DNA libraries in a binary-BAC (BIBAC) vector. Plant J., 18: 223-229.
CrossRefDirect Link - Hansen, G. and M.D. Chilton, 1996. Agrolistic transformation of plant cells: Integration of T-strands generated in planta. Proc. Natl. Acad. Sci., 93: 14978-14983.
Direct Link - Hiei, Y., S. Ohta, T. Komari and T. Kumashiro, 1994. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J., 6: 271-282.
CrossRefDirect Link - Hiei, Y., S. Ohta, T. Komari and T. Kumashiro, 1994. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J., 6: 271-282.
CrossRefDirect Link - Higo, K. and H. Higo, 1996. Cloning and characterization of the rice CatA catalase gene, a homologue of the maize Cat3 gene. Plant Mol. Biol., 30: 505-521.
CrossRefDirect Link - Holm, P.B., O. Olsen, M. Schnorf, H. Brinch-Pederse and S. Knudsen, 2000. Transformation of barley by microinjection into isolated zygote protoplasts. Transgenic Res., 9: 21-32.
CrossRef - Holzberg, S., P. Brosio, C. Gross and G.P. Pogue, 2002. Barley stripe mosaic virus-induced gene silencing in a monocot plant. Plant J., 30: 315-327.
CrossRef - Horsch, R.B., J.E. Fry, N.L. Hoffmann, D. Eichholtz, S.G. Rogers and R.T. Fraley, 1985. A simple and general method for transferring genes into plants. Science, 227: 1229-1231.
CrossRefDirect Link - Hu, C.Y. and L. Wang, 1999. In-planta soybean transformation technologies developed in China: Procedure, confirmation and field performance. In vitro Cell Dev. Biol. Plant, 35: 417-420.
CrossRefDirect Link - Hull, A.K., V. Yusibov and V. Mett, 2005. Inducible expression in plants by virus-mediated transgene activation. Transgenic Res., 14: 407-416.
CrossRef - Curtis, I.S. and H.G. Nam, 2001. Transgenic radish (Raphanus sativus L. longipinnat us Bailey) by floral-dip method-plant development and surfactant are important in optimizing transformation efficiency. Transgenic Res., 10: 363-371.
CrossRef - Ishida, Y., H. Saito, S. Ohta, Y. Hiei, T. Komari and T. Kumashiro, 1996. High efficiency transformation of maize (Zea mays L.) mediated by Agrobacterium tumefaciens. Nat. Biotechnol., 15: 745-750.
CrossRefPubMedDirect Link - Jefferson, R.A., 1987. Assaying chimeric genes in plants: The GUS gene fusion system. Plant Mol. Biol. Rep., 5: 387-405.
CrossRefDirect Link - Jia, H., Y. Pang, X. Chen and R. Fang, 2006. Removal of the selectable marker gene from transgenic tobacco plants by expression of Cre recombinase from a Tobacco mosaic virus vector through agroinfection. Transgenic Res., 15: 375-384.
CrossRef - Jia, H.G., Y.Q. Pang and R.X. Fang, 2003. Agroinoculation as a simple way to deliver a Tobacco mosaic virus based expression vector. Acta Bot. Sin., 45: 770-773.
Direct Link - Joersbo, M. and J. Brunstedt, 1990. Direct gene transfer to plant protoplasts by mild sonification. Plant Cell Rep., 9: 207-210.
CrossRef - Jones, H.D., A. Doherty and H. Wu, 2005. Review of methodologies and a protocol for the Agrobacterium-mediated transformation of wheat. Plant Methods, 1: 5-5.
CrossRef - Jones-Villeneuve, E., B. Huang, I. Prudhomme, S. Bird, R. Kemble, J. Hattori and B. Miki, 1995. Assessment of microinjection for introducing DNA into uninuclear microspores of rapeseed. Plant Cell Tissue Org. Cult., 40: 97-100.
CrossRefDirect Link - Kaeppler, H., W. Gu, D.A. Somers, H.W. Rines and A.F. Cockburn, 1990. Silicon carbide fiber-mediated DNA delivery into plant cells. Plant Cell Rep., 9: 415-418.
CrossRef - Kaeppler, H., D.A. Somers, H.W. Rines and A.F. Cockburn, 1992. Silicon carbide fiber-mediated stable transformation of plant cells. Theor. Applied Genet., 84: 560-566.
Direct Link - Kifle, S., M. Shao, C. Jung and D. Cai, 1999. An improved transformation protocol for studying gene expression in hairy roots of sugar beet (Beta vulgaris L.). Plant Cell Rep., 18: 514-519.
CrossRef - Kikkert, J.R., 1993. The Biolistic PDS-1000/He device. Plant Cell Tissue Org. Cult., 33: 221-226.
CrossRef - Kikkert, J.R., J.R. Vidal and B.I. Reisch, 2004. Stable Transformation of Plant Cells by Particle Bombardment/Biolistics. In: Methods in Molecular Biology. Transgenic Plants: Methods and Protocols, Peña, L. (Ed.). Humana Press Inc., Totowa, NJ., ISBN: 978-1-58829-263-6, pp: 61-78.
CrossRef - Klimaszewska, K., Y. Devantier, D. Lachance, M.A. Lelu and P.J. Charest, 1997. Larix laricina (tamarack): Somatic embryogenesis and genetic transformation. Can. J. For. Res., 27: 538-550.
CrossRef - Klimaszewska, K., D. Lachance, G. Pelletier, M.A. Lelu and A. Seguin, 2001. Regeneration of transgenic Picea glauca, P. mariana and P. abies after cocultivation of embryogenic tissue with Agrobacterium tumefaciens. In vitro Cell Dev. Biol. Plant, 37: 748-755.
CrossRef - Komatsu, A., M. Ohtake, H. Hasegawa, T. Terakawa and K. Wakasa, 2006. Transgenic rice for animal feed with high tryptophan content generated by a selectable marker- and vector backbone-free technology. Plant Biotechnol., 23: 39-46.
CrossRefDirect Link - Kumar, V., A. Sharma, B.C.N. Prasad, H.B. Gururaj and G.A. Ravishankar, 2006. Agrobacterium rhizogenes mediated genetic transformation resulting in hairy root formation is enhanced by ultrasonication and acetosyringone treatment. Elect. J. Biotechnol., 9: 349-357.
CrossRef - Lesser, R.M., V. Ziegler-Graff, A. Reutenauer, E. Herrbach and O. Lemaire et al., 1992. Agroinfection as an alternative to insects for infecting plants with beet Western yellows luteovirus. Proc. Natl. Acad. Sci. USA., 89: 9136-9140.
CrossRefDirect Link - Levee, V., M.A. Lelu, L. Jouanin, D. Cornu and G. Pilate, 1997. Agrobacterium tumefaciens-mediated transformation of hybrid larch (Larix kaempferi × L. decidua) and transgenic plant regenerationn. Plant Cell Rep., 16: 680-685.
CrossRefDirect Link - Li, L., R. Qu, A. de Kochko, C.M. Fauquet and R.N. Beachy, 1993. An improved rice transformation system using the biolistic method. Plant Cell Rep., 12: 250-255.
CrossRefDirect Link - Liu, Y.G., Y. Shirano, H. Fukaki, Y. Yanai, M. Tasaka, S. Tabata and D. Shibata, 1999. Complementation of plant mutants with large genomic DNA fragments by a transformation-competent artificial chromosome vector accelerates positional cloning. Proc. Natl. Acad. Sci., 96: 6535-6540.
CrossRefDirect Link - Lu, R., A.M. Martin-Hernandez, J.R. Peart, I. Malcuit and D.C. Baulcombe, 2003. Virus-induced gene silencing in plants. Methods, 30: 296-303.
CrossRef - Luo, Z.X. and R. Wu, 1988. A simple method for the transformation of rice via the pollen-tube pathway. Plant Mol. Biol. Rep., 6: 165-174.
CrossRefDirect Link - Ma, J.K.C., P.M.W. Drake and P. Christou, 2003. The production of recombinant pharmaceutical proteins in plants. Nat. Rev. Genet., 4: 794-805.
CrossRef - Manahan, S.H. and T.R. Steck, 1997. The viable but nonculturable state in Agrobacterium tumefaciens and Rhizobium meliloti. FEMS Microbiol. Ecol., 22: 29-37.
CrossRefDirect Link - Marillonnet, S., C. Thoeringer, R. Kandzia, V. Klimyuk and Y. Gleba, 2005. Systemic Agrobacterium tumefaciens-mediated transfection of viral replicons for efficient transient expression in plants. Nature Biotechnol., 23: 718-723.
CrossRef - May, G.D., R. Afza, H.S. Mason, A. Wiecko, F.J. Novak and C.J. Arntzen, 1995. Generation of transgenic banana (Musa acuminata) plants via Agrobacterium-mediated transformation. Bio/Technol., 13: 486-492.
CrossRefDirect Link - McCabe, D. and P. Christou, 1993. Direct DNA transfer using electric discharge particle acceleration (ACCELLTM technology). Plant Cell Tissue Org. Cult., 33: 227-236.
CrossRef - Medina-Bolívar, F. and C. Cramer, 2004. Production of Recombinant Proteins by Hairy Roots Cultured in Plastic Sleeve Bioreactors. In: Methods in Molecular Biology. Recombinant Gene Expression: Reviews and Protocols, Balbas, P. and A. Lorence (Eds.). Humana Press Inc., Totowa, NJ.,ISBN: 978-1-58829-262-9, pp: 351-363.
CrossRef - Meselson, M. and R. Yuan, 1968. DNA restriction enzyme from E. coli. Nature, 217: 1110-1114.
CrossRef - Morikawa, H. and Y. Yamada, 1985. Capillary microinjection into protoplasts and intranuclear localization of injected materials. Plant Cell Physiol., 26: 229-236.
Direct Link - Mu, H.M., S.J. Liu, W.J. Zhou, Y.X. Wen, W.J. Zhang and R.X. Wei, 1999. Transformation of wheat with insecticide gene of arrowhead proteinase inhibitors by pollen tube pathway and analysis of transgenic plants. Yi Chuan Xue Bao, 26: 634-642.
PubMedDirect Link - Mugnier, J., 1988. Establishment of new axenic hairy root lines by inoculation with Agrobacterium rhizogenes. Plant Cell Rep., 7: 9-12.
CrossRefDirect Link - Mullen, J., G. Allen, A. Blowers and E. Earle, 1998. Biolistic transfer of large DNA fragments to tobacco cells using YACs retrofitted for plant transformation. Mol. Breed., 4: 449-457.
CrossRefDirect Link - Neuhaus, G., G. Spangeberg, O. Mittelsten-Scheid and H.G. Schweizer, 1987. Transgenic rape seed plants obtained by microinjection of DNA into microspore derived embryoids. Theor. Applied Genet., 75: 30-36.
CrossRefDirect Link - Norris, S.R., S.E. Meyer and J. Callis, 1993. The intron of Arabidopsis thaliana polyubiquitin genes is conserved in location and is a quantitative determinant of chimeric gene expression. Plant Mol. Biol., 21: 895-906.
PubMedDirect Link - Olhoft, P.M. and D.A. Somers, 2001. L-cysteine increases agrobacterium mediated T-DNA delivery into soybean cotyledonary-node cells. Plant Cell Rep., 20: 706-711.
CrossRef - Opabode, J.T., 2006. Agrobacterium-mediated transformation of plants: Emerging factors that influence efficiency. Mol. Biol. Rev., 1: 12-20.
Direct Link - Paszkowski, J., R.D. Shillito, M. Saul, V. Mandák and T. Hohn et al., 1984. Direct gene transfer to plants. EMBO J., 3: 2717-2722.
Direct Link - Perl, A., O. Lotan, M. Abu-Abied and D. Holland, 1996. Establishment of an Agrobacterium-mediated transformation system for grape (Vitis vinifera L.): The role of antioxidants during grape-Agrobacterium interactions. Natl. Biotechnol., 14: 624-628.
PubMedDirect Link - Petit, A., J. Stougarrd, A. Kuhle, K.A. Marcker and J. Tempe, 1987. Transformation and regeneration of the legume Lotus corniculatus: A system for molecular studies of symbiotic nitrogen fixation. Mol. Gen. Genet., 207: 245-250.
CrossRefDirect Link - Petolino, J.F., N.L. Hopkins, B.D. Kosegi and M. Skokut, 2000. Whisker mediated transformation of embryogenic callus of maize. Plant Cell Rep., 19: 781-786.
Direct Link - Phillips, R.L., S.M. Kaeppler and P. Olhoft, 1994. Genetic instability of plant tissue cultures: Breakdown of normal controls. Proc. Natl. Acad. Sci. USA., 91: 5222-5226.
Direct Link - Porta, C. and G.P. Lomonossoff, 2002. Viruses as vectors for the expression of foreign sequences in plants. Biotechnol. Genet. Eng. Rev., 19: 245-291.
Direct Link - Potrykus, I., 1991. Gene transfer to plants: Assessment of published approaches and results. Annu. Rev. Plant Physiol. Plant Mol. Biol., 42: 205-225.
Direct Link - Racoczy-Trijanowska, M., 2002. Alternative methods of plant transformation-a short review. Cell. Mol. Biol. Lett., 7: 849-858.
Direct Link - Rhodes, C.A., D.A. Pierce, I.J. Mettler, D. Mascarenhas and J.J. Detmer, 1988. Genetically transformed maize plants from protoplasts. Sci., 240: 204-207.
CrossRefDirect Link - Richter, L.J., Y. Thanavala, C.J. Arntzen and H.S. Mason, 2000. Production of hepatitis B surface antigen in transgenic plants for oral immunization. Nature Biotechnol., 18: 1167-1171.
Direct Link - Rommens, C.M., J.M. Humara, J. Ye, H. Yan and C. Richael et al., 2004. Crop improvement through modification of the plant's own genome. Plant Physiol., 135: 421-431.
CrossRef - Rommens, C.M., 2004. All-Native DNA transformation: a new approach to plant genetic engineering. Trends Plant Sci., 9: 457-464.
CrossRef - Rossi, L., B. Hohn and B. Tinland, 1996. Integration of complete transferred DNA units is dependent on the activity of virulence E2 protein of Agrobacterium tumefaciens. Proc. Natl. Acad. Sci. USA., 93: 126-130.
Direct Link - Ruiz, M.T., O. Voinnet and D.C. Baulcombe, 1998. Initiation and maintenance of virus-induced gene silencing. Plant Cell, 10: 937-946.
Direct Link - Sanford, J.C., T.M. Klein, E.D. Wolf and N. Allen, 1987. Delivery of substances into cells and tissues using a particle bombardment process. Particulate Sci. Technol., 5: 27-37.
CrossRefDirect Link - Sanford, J.C., F.D. Smith and J.A. Russell, 1993. Optimizing the biolistic process for different biological applications. Methods Enzymol., 217: 483-509.
CrossRef - Sanford, J.C., 2000. The development of the biolistic process. In vitro Cell Dev. Biol. Plant, 36: 303-308.
CrossRef - Santarem, E.R., H.N. Trick, J.S. Essig and J.J. Finer, 1998. Sonication-assisted Agrobacterium-mediated transformation of soybean immature cotyledons: Optimization of transient expression. Plant Cell Rep., 17: 752-759.
CrossRefDirect Link - Sawahel, W., 1996. Ultrasound-mediated stable transformation of potato tuber discs. Biotechnol. Technol., 10: 821-824.
CrossRefDirect Link - Sawahel, W.A., 2001. Stable genetic transformation of cotton plants using polybrene-spermidine treatment. Plant Mol. Biol. Rep., 19: 377a-377f.
Direct Link - Sawahel, W.A., 2002. The production of transgenic potato plants expressing human Ü-interferon using lipofection- mediated transformation. Cell. Mol. Biol. Lett., 7: 19-29.
Direct Link - Schaefer, D.J., 2002. A new moss genetic: Targeted mutagenesis in Physcomitrella patens. Annu. Rev. Plant Biol., 53: 477-501.
Direct Link - Schnorf, M., G. Neuhaus-Url, A. Galli, S. Lida, I. Potrykus and G. Neuhaus, 1991. An improvement approach for transformation of plant cells by microinjection; molecular and genetic analysis. Transgenic Res., 1: 23-30.
CrossRefDirect Link - Shewry, P.R., A. Tatham, N.G. Halford, J.H.A. Barker and U. Hannapel et al., 1994. Opportunities for manipulating the seed protein composition of wheat and barley in order to improve quality. Transgenic Res., 3: 3-12.
CrossRefDirect Link - Shillito, R., M. Saul, J. Paszkowski, M. Muller and I. Potrykus, 1985. High efficiency direct transfer to plants. Nature Biotechnol., 3: 1099-1103.
CrossRef - Shimamoto, K., R. Terada, T. Izawa and H. Fugimoto, 1989. Fertile transgenic rice plants regenerated from transformed protoplasts. Nature, 338: 274-276.
CrossRefDirect Link - Shou, H., R.G. Palmer and K. Wang, 2002. Irreproducibility of the soybean pollen-tube pathway transformation procedure. Plant Mol. Biol. Rep., 20: 325-334.
CrossRef - Singh, N. and H.S. Chawla, 1999. Use of silicon carbide fibers for Agrobacterium-mediated transformation in wheat. Curr. Sci., 76: 1483-1485.
Direct Link - Singh, R.J., 2003. Chromosomal Aberrations in Cell and Tissue Culture. In: Plant Cytogenetics, Singh, R.J. (Ed.). CRC Press, Boca Raton, FL., pp: 307-326.
Direct Link - Sivamani, E., P. Shen, N. Opalka, R.N. Beachy and C.M. Fauquet, 1996. Selection of large quantities of embryogenic subcultured calli from indica rice seeds for production of fertile transgenic plants using the biolistic method. Plant Cell Rep., 15: 322-327.
CrossRefDirect Link - Smith, H.O. and K.W. Wilcox, 1970. A restriction enzyme from Hemophilus influenzae. I. Purification and general properties. J. Mol. Biol., 51: 379-391.
PubMedDirect Link - Songstad, D.D., D.A. Somers and R.J. Griesbach, 1995. Advances in alternative DNA delivery techniques. Plant Cell Tissue Organ Culture, 40: 1-15.
CrossRefDirect Link - Southgate, E.M., M.R. Davey, J.B. Power and R. Marchant, 1995. Factors affecting the genetic engineering of plants by microprojectile bombardment. Biotechnol. Adv., 13: 631-651.
CrossRef - Soyfer, V.N., 1980. Hereditary variability of plants under the action of exogenous DNA. Theor. Applied Genet., 58: 225-235.
CrossRefDirect Link - Srivastava, V. and D.W. Ow, 2001. Biolistic mediated site-specific integration in rice. Mol. Breed., 8: 345-350.
CrossRef - Gelvin, S.B., 2003. Agrobacterium mediated plant transformation: The biology behind the gene-jockeying tool. Microbiol. Mol. Biol. Rev., 67: 16-37.
CrossRefPubMedDirect Link - Storm, M.M.H., C. Van der Schoot, M. Prins, R. Kormeling and J.W.M. Van Lent et al., 1998. A comparison of two methods of microinjection for assessing altered plasmodesmal gating in tissue expressing viral movement proteins. Plant J., 13: 131-140.
Direct Link - Supartana, P., T. Shimizu, H. Shioiri, M. Nagava and M. Nozue et al., 2005. Development of simple and efficient in planta transformation method for rice (Oryza sativa) using Agrobacterium tumefacience. J. Biosci. Bioeng., 100: 391-397.
PubMedDirect Link - Tang, W., 2003. Additional virulence genes and sonication enhance Agrobacterium tumefaciens-mediated loblolly pine transformation. Plant Cell Rep., 21: 555-562.
CrossRef - Taylor, N.J. and C.M. Fauquet, 2002. Microparticle bombardment as a tool in plant science and agricultural biotechnology. DNA Cell Biol., 21: 963-977.
CrossRef - Teli, N.P. and M.P. Timko, 2004. Recent developments in the use of transgenic plants for the production of human therapeutics and biopharmaceuticals. Plant Cell Tissue Org. Cult., 79: 125-145.
CrossRef - F. Tenllado and J. R. Díaz-Ruíz, 2001. Double-Stranded RNA-Mediated Interference with Plant Virus Infection. J. Virol., 75: 12288-12297.
CrossRef - Tepfer, D., 1984. Transformation of several species of higher plants by Agrobacterium rhizogenes. sexual transmission of the transformed genotype and phenotype. Cell, 37: 959-967.
PubMedDirect Link - Terakawa, T., H. Hisakazu and Y. Masanori, 2005. Efficient whisker mediated gene transformation in a combination with supersonic treatment. Breed. Sci., 55: 456-458.
Direct Link - Tjokrokusumo, D., T. Heinrich, S. Wylie, R. Potter, J. McComb, 2000. Vacuum infiltration of Petunia hybrida pollen with Agrobacterium tumefaciens to achieve plant transformation. Plant Cell Rep., 19: 792-797.
CrossRef - Topfer, R., B. Gronenborn, J. Schell and H. Steinbiss, 1989. Uptake and transient expression of chimeric genes in seed-derived embryos. Plant Cell, 1: 133-139.
CrossRefDirect Link - Touraev, A., E. Stoger, V. Voronin and E. Heberle-Bors, 1997. Plant male germ line transformation. Plant J., 12: 949-956.
Direct Link - Trick, H.N. and J.J. Finer, 1997. SAAT: Sonication-assisted Agrobacterium-mediated transformation. Transgenic Res., 6: 329-336.
CrossRefDirect Link - Turnage, M.A., N. Muangsan, C.G. Peele and D. Robertson, 2002. Geminivirus-based vector for gene silencing in Arabidopsis. Plant J., 30: 107-114.
CrossRef - Uchimiya, H., T. Fushimi, H. Hashimoto, H. Harada, K. Syono and Y. Sugawara, 1986. Expression of a foreign gene in callus derived from DNA-treated protoplasts of rice (Oryza sativa L.). Mol. Gen. Genet., 204: 204-207.
CrossRefDirect Link - Vik, N.I., A.K. Hvoslef-Eide, H. Gjerde and K. Bakke, 2001. Stable transformation of poinsettia via electrophoresis. Acta Horticult., 560: 101-103.
Direct Link - Walter, C., L.J. Grace, A. Wagner, D.W.R. White and A.R. Walden et al., 1998. Stable transformation and regeneration of transgenic plants of Pinus radiata D. Plant Cell Rep., 17: 460-468.
Direct Link - Walter, C., L.J. Grace, S.S. Donaldson, J. Moody and E. Gemmell et al., 1999. An efficient biolistic transformation protocol for Picea abies embryogenic tissue and regeneration of transgenic plants. Can. J. For. Res., 29: 1539-1546.
CrossRef - Wang, K., P. Drayton, B. Frame, J. Dunwell and J.A. Thompson, 1995. Whisker mediated plant transformation: An alternative technology. In Vitro Cell. Dev. Biol. Plant, 31: 101-104.
CrossRefDirect Link - Weber, S., W. Friedt, N. Landes, J. Molinier and C. Himber et al., 2003. Improved Agrobacterium-mediated transformation of sunflower (Helianthus annuus L): Assessment of macerating enzymes and sonication. Plant Cell Rep., 21: 475-482.
CrossRef - Wongsamuth, R. and P.M. Doran, 1997. Production of monoclonal antibodies by tobacco hairy roots. Biotechnol. Bioeng., 54: 401-415.
PubMedDirect Link - Wordragen, M.V., R. Shakya, R. Verkerk, R. Peytavis, A.V. Kammen and P. Zabel, 1997. Liposome-mediated transfer of YAC DNA to tobacco cells. Plant Mol. Biol. Rep., 15: 170-178.
CrossRefDirect Link - Wyber, J.A., J. Andrew and A. D'Emanuele, 1997. The use of sonication for the efficient delivery of plasmid DNA into cells. Pharma. Res., 14: 750-756.
CrossRefDirect Link - Zhang, L.J., L.M. Chen, N. Xu, N.M. Zhao and C.G. Li et al., 1991. Efficient transformation of tobacco by ultrasonication. Biotechnology, 9: 996-997.
CrossRefDirect Link - Zhang, S., L. Chen, R. Qu, P. Marmey, R.N. Beachy and C.M. Fauquet, 1996. Regeneration of fertile transgenic indica (group 1) rice plants following microprojectile transformation of embryogenic suspension culture cells. Plant Cell Rep., 15: 465-469.
CrossRefDirect Link - Zhang, W., S. Subbarao, P. Addae, A. Shen and C. Armstrong et al., 2003. Cre/lox mediated gene excision in transgenic maize (Zea mays L.) plants. Theor. Applied Genet., 107: 1157-1168.
CrossRef - Zhao, Z.Y., W. Gu, T. Cai, L. Tagliani and D. Hondred et al., 2002. High throughput genetic transformation mediated by Agrobacterium tumefaciens in maize. Mol. Breed., 8: 323-333.
CrossRefDirect Link - Zhou, G.Y., J. Weng, Y. Zeng, J. Huang, S. Qian and G. Liu, 1983. Introduction of exogenous DNA into cotton embryos. Meth. Enzymol., 101: 433-481.
PubMedDirect Link - Zhu, Z., B. Sun, C. Liu, G. Xiao and X. Li, 1993. Transformation of wheat protoplasts mediated by cationic liposome and regeneration of transgenic plantlets. Chin. J. Biotechnol., 9: 257-261.
PubMedDirect Link - Hoekema, A., P. Hirsch, P.J.J. Hooykas and R.A. Schilperoort, 1983. A binary vector strategy based on separation of vir and T-region of the Agrobacterium tumefaciens Ti plasmid. Nature, 303: 179-180.
CrossRefDirect Link - Trieu, A.T., S.H. Burleigh, I.V. Kardailsky, I.E. Maldonado-Mendoza and W.K. Versaw et al., 2000. Transformation of Medicago truncatula via infiltration of seedlings or flowering plants with Agrobacterium. Plant J., 22: 531-541.
CrossRefDirect Link - Fischer, R., E. Stoger, S. Schillberg, P. Christou and R.M. Twyman, 2004. Plant-based production of biopharmaceuticals. Curr. Opin. Plant Biol., 7: 152-158.
CrossRefDirect Link