
Sauvaphap Ai-Noi
Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Suraini Abd-Aziz
Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Norjahan Alitheen
Faculty of Biotechnology and Biomolecular Sciences, Universiti Putra Malaysia, 43400 UPM Serdang, Selangor, Malaysia
Osman Hassan
Faculty of Science and Technology, School of Chemical Sciences and Food Technology, Universiti Kebangsaan Malaysia, 43600 Bangi, Selangor, Malaysia
Mohamed Ismail Abdul Karim
Department of Biotechnology Engineering, Faculty of Engineering, International Islamic University, Jalan Gombak, 53100 Kuala Lumpur, Malaysia







ABSTRACT
The aim of the study was to optimized cyclodextrin glycosyltransferase (CGTase) production using local isolated strain MK6 which was identified as Bacillus sp. Optimum activity obtained at temperature of 70 °C and the enzyme shows a wide range of pH stability ranging from 4-10 when stored at 4 °C for 24 h and temperature stability ranging from 30-80 °C at 1 h incubation period. The CGTase activity was even maintained at 0.4 U mL-1 at 90 °C for 40 min incubation. Prior to optimization of CGTase production, selection for the best carbon source through detection using modified phenolphthalein method containing different types of starch were performed. Sago starch gave significant result and was used for further optimization using statistical analysis namely Response Surface Methodology (RSM) approach. The optimal calculated values were 3.34% sago starch, initial pH of 10.15 and agitation speed of 187 rpm; with predicted activity of 2.07 U mL-1 of CGTase. These predicted optimal parameters were confirmed in the laboratory and the final CGTase activity obtained was very close to the predicted value of 2.56 U mL-1.
PDF Abstract XML References Citation
How to cite this article
Sauvaphap Ai-Noi, Suraini Abd-Aziz, Norjahan Alitheen, Osman Hassan and Mohamed Ismail Abdul Karim, 2008. Optimization of Cyclodextrin Glycosyltransferase Production by Response Surface Methodology Approach
. Biotechnology, 7: 10-18.
DOI: 10.3923/biotech.2008.10.18
URL: https://scialert.net/abstract/?doi=biotech.2008.10.18
DOI: 10.3923/biotech.2008.10.18
URL: https://scialert.net/abstract/?doi=biotech.2008.10.18
INTRODUCTION
Cyclodextrin glycosyltransferase (CGTase) or [1,4-α-D-glucopyranosyl]-transferase is an extracellular enzyme, which degrades starches into cyclodextrin (CDS) molecules via cyclization reaction. Cyclization happens when a linear oligosaccharide (starch) chain is cleaved and the new reducing end sugar is transferred to the non-reducing end sugar of the same chain. Therefore cyclodextrins are cyclic oligosaccharides consisting of 6-12 units of glucose joined by the α-1, 4-linkages. CGTases also catalyses two intermolecular transglycosylation reactions: coupling, in which a cyclodextrin ring is cleaved and transferred to an acceptor maltooligosaccharide substrate and disproportionation, in which a linear maltooligosaccharide is cleaved and the new reducing end sugar is transferred to an acceptor maltooligosaccharide substrate. Besides these reactions, the enzyme has a weak hydrolyzing activity (Penninga et al., 1995; Bart et al., 2000). Cyclodextrins with 6, 7 and 8 glucose units are most common and also known as α-, β-and γ-cyclodextrin, respectively.
CGTases with varying properties are produced by bacteria mainly belonging to the Bacillus species, by submerged culture in a complex medium (Adriana et al., 2002). Some of the known sources of CGTase producers are Bacillus macerans, Bacillus subtilis, Bacillus stereothermophillus, Bacillus megaterium, Klebsiella pneumonia and micrococcus species. Alkalophilic microorganism is also known to produce unusual enzyme that can be used in industrial and other processes. All known CGTases (Bart et al., 2000) produce a mixture of cyclodextrins (and linear malto-oligosaccharides) when incubated with starch. The CGTase crude enzyme isolated from local Bacillus sp. produces alpha (α) and beta (β) cyclodextrin only.
However a CGTase, which only produces a single type of cyclodextrin, is industrially favorable. The aims of this research are to optimise CGTase production using local isolated strain by Response Surface Methodology approach and finally production of cyclodextrin using the optimized condition obtained.
MATERIALS AND METHODS
Screening and isolation of microorganism: Soil samples were collected from various places such as sago plantation, herbs plantation and soil near marketplace in Malaysia. Criteria for soil selection were mainly based on its soil`s pH (pH 5-6). Collected soil samples were suspended in sterile saline and the solid particles were allowed to settle. Plating was done at high pH (pH 7-9) to discourage the growth of fungal species and screening was done according to Park et al. (1989). Bacterial colonies with yellowish clearance zone were selected and streaked onto Horikoshi-Phenolphthalein (PHP) plate for several times until uniform colonies were formed. Suspensions of vegetative cells were grown in Horikoshi Broth (HB) for 24 h at 37 °C, mixed with sterilized glycerol (20% v/v) and kept in 1 mL aliquots at -80 °C until further used.
Morphological characterizations: Microscopic morphological characteristics include the shapes of cells and the characteristic arrangements of cells in groups. On the other hand, the macroscopic properties of pure cultures including the colour of colonies, shape and odour can be considered as among the morphological characteristics of microbes.
Biochemical identification of microorganism: Biochemical test was used to characterize the isolated microorganisms. API 50 CHB/E Medium kit was used in assistance of confirmation of the genus of bacteria based on biochemical reaction.
Preparation of bacterial inoculums: Bacillus sp. (Strain MK6) were grown in 20 mL Horikoshi medium (Park et al., 1989) contained the following solutions after autoclaving. Solution 1: Starch 1.0% (w/v), peptone 0.5% (w/v), yeast extracts 0.5%(w/v), K2HPO4 0.1% (w/v), MgSO4 0.02% (w/v). Solution 2: Na2CO3 1.0% (w/v). The culture was incubated at 37 °C, agitated at 200 rpm for 18 h. Cells were harvested by centrifugation at 5000 rpm for 5 min and washed once with normal saline solution (0.85% w/v NaCl) and were then suspended in normal saline solution to give an optical density reading of 0.5 at 660 nm, using a UV-spectrophotometer.
Production of crude CGTase: Ten percent (v/v) of bacterial inoculums was inoculated into 100 mL of Horikoshi Medium in a 500 mL conical shake flask. The bacteria culture was incubated at 37 °C in an incubator shaker for 24 h. At the end of incubation period, 1.0 mL of the culture was removed and separated by centrifugation at 8000 rpm for 10 min at 4 °C. The supernatant was assayed for CGTase activity and used as crude enzyme solution.
Optimization using Response Surface Methodology (RSM) approach: Response Surface Methodology or RSM is a collection of statistical techniques for designing experiments, building models, evaluating the effects of factors and searching optimum conditions of factors for desirable responses (Montgomery, 1976). Factorial designs of a limited set of variables is advantages in relation to the conventional method of the manipulation of a single parameter trial, because such an approach frequently fails to locate optimal conditions for the process due to its failure to consider the effect of possible interaction between factors. In direct fermentation of starch to production of CGTase, the preliminary test had indicated that the CGTase production is significantly affected by the concentration of starch, pH values and agitation speed. However, the true relationship between the response, Y and the independent variables is unknown. Therefore the Y can be written as a function of the levels of the variables χ1 with a significant influence on CGTase production (Eq. 1).
(1) |
The nature of this function is unknown but usually these kind of responses can be approximated by a second-order polynomial, as shown in Eq. 2:
(2) |
Where, Y is the predicted response, βo, βi, βii and βij are constant and regression coefficients of the model and χi is the independent variable in coded values. In this study, k = 3 because there were three independent variables involved. Thus the mathematical relationship connecting to the three variables and the response from Eq. 2 becomes:
(3) |
Where:
| Y | = The predicted response, |
| Bo | = Offset term, |
| B1, B2, B3 | = Linear coefficients, |
| B11, B22 and B33 | = The quadratic coefficients, |
| B12, B13 and B23 | = The cross product coefficients, |
| X1, X2 and X3 | = The independent variables. |
| Table 1: | Actual factor levels corresponding to the coded factor levels |
 | |
A randomization step was conducted using completely randomized design. Each treatment consist of 3 replicates were treated and given a number started from 00 until 26 with 27 total treatment. Using a random digit table, the treatment was assigned according to the number appeared in order. The range and the levels of the variables investigated in this study are given in Table 1. Response Surface Design using MINITAB software (version 13.1) was used to conduct the RSM analysis. A second-order polynomial expression of three variables as Eq. 2 was fitted. In order to visualize the relationship between the response (Y) and the experimental levels of each factor, surface plots were generated from the fitted second order polynomial equation. After collecting the experimental data and determining the optimum pH, sago starch concentration and agitation speed, optimum response was verified by real experiments under optimum condition. Response was monitored and results were compared with model prediction.
CGTase assay: CGTase activity was determined using phenolphthalein assay (Kaneko et al., 1987).
Determination of cyclodextrin and sugars: The cyclodextrin and sugars concentration was determined using HPLC. Samples were first centrifuge to remove suspended solids and biomass. Centrifugation was done at 12,000 rpm for 10 min. The supernatant was then filtered through a nylon filter paper with 0.45 μm pore size. The liquid chromatograph comprised of a Jasco-PU 980 pump (Jasco, Japan) and a differential refractive index detector (Perkin Elmer LC-25, USA) with a sensitivity of 5x10-5 RIU. Data integration was done using Borwin Software package V1.21 for liquid chromatography integration. The column used was Merck NH2 column (Purospher Star; 5 μm, 250x4.6 mm). Injections were carried out using a 20 μL sample loop at room temperature (25-28 °C) with 75% acetonitrile as the mobile phase with the flow rate of 1.0 mL min-1. Samples (optimized crude enzyme) and cyclodextrin standard was prepared using deionized water.
RESULTS AND DISCUSSION
Isolation of microorganism: A total of 250 isolates have been successfully isolated in this study. CGTase activity was detected using plate assay containing soluble starch as substrate and phenolphthalein as indicator. However strain MK6 gave the highest CGTase activity through the formation of large diameter (5 cm) of clearance zone as a qualitative measurement.
Morphological characteristics: Gram staining was done to determine the basic morphological characteristics of the isolated microorganism. Gram staining showed strain MK6 was gram positive, rod in shape with rounded ends
Biochemical identification of strain MK6 using API 50 CHB/E medium: Biochemical identification was done after selection of the best strain for production of CGTase enzyme. The choices of medium used in this experiment depend much on the knowledge of morphological characteristic of the strain MK6 done earlier. API 50 CHB/E medium was used based on these characteristics as shown in Table 2.
Characteristics above are indicative of a Bacillus sp. Therefore further confirmation using biochemical testing were done using the API 50 CHB/E medium, analyzed using APILAB PLUS (V 3.2.2). During incubation, carbohydrates were fermented to acids which resulted in a decrease in pH, detected by the colour change of the indicator. Result obtained showed good identification to the genus of the isolated bacteria, which is of Bacillus sp. with 85% similarity.
Preliminary study for optimization of CGTase production
The effect of pH on CGTase activity and stability: Enzyme activity was measured using the standard assay method by varying the pH values ranging from pH 3 to 10 at 70 °C. It was observed that optimum pH of the crude CGTase was at pH 6.0 (Fig. 1). Nonetheless, the enzyme showed only slight activity at pH 4.0 and 9.0. This
| Table 2: | Summary of morphological characteristics of strain MK6 |
 | |
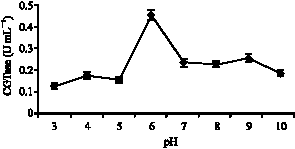 | |
| Fig. 1: | Effect of pH on the enzyme activity of Bacillus MK6 after 10 min incubation |
suggests that the CGTase from Bacillus MK6 requires a near neutral pH range to perform its cyclization reaction. Extreme pH values were unsuitable for the enzyme to carry out cyclization activity. Most of the reported CGTase exhibited optimum pH ranging from 5.0 to 8.0 (Ho et al., 2004). Some purified CGTase enzyme however may exhibit more than 1 optimum peak. This phenomenon suggested the existence of more than one peak were due to presence of different CGTases (acid-, neutral-and alkaline) in the culture filtrate (Turnes and Bahar, 1996). It was also reported that at 60 °C, the CGTase from alkalophilic Bacilllus sp. showed stability over a wide range of pH of 6-10. However, the enzyme activity decreased drastically beyond that range and almost lost its activity below pH 4.0 and above pH 11.0 (Cao et al., 2005).
The effect of temperature on CGTase activity and stability: The effect of different temperatures on CGTase activity was measured at pH 6.0 following standard assay. The optimum temperature for CGTase from Bacillus MK6 was 70 °C using sago starch as the substrate (Fig. 2). It is interesting to note the presence of another peak at 90 °C, suspected to be an isozyme of crude CGTase used, having different physical characteristics such as temperatures or pH but catalyzes the same reaction. Most reported alkalophillic bacteria have optimum temperatures ranging from 45 to 60 °C, except for Bacillus stearothermophillis which have optimum temperature of 80 °C (Rita and Rajni, 2002). There are no other known reports of CGTase with optimum temperature exceeding 60 °C for alkalophillic bacteria, except the one reported by Tien (2001), where its optimum temperature was found to be at 70 °C. Presence of other protein in crude CGTase enzyme increases its stability hence giving another optimum peak at 90 °C.
The effect of temperature on stability of crude CGTase enzyme was also investigated and the enzyme was observed to be stable ranging from temperatures 30-80 °C (for 1 h incubation). The isolated crude CGTase enzyme was found active even at 90 °C in the first 40 min incubation period before being totally inactivated (Fig. 3). It may be due to influence of other proteins presence in crude enzyme that enhanced its stability over high temperature.
Cao et al. (2005) found that the CGTase isolated from alkalophilic Bacillus sp. in China shows wide thermal stability (40-70 °C) when kept in buffer at pH 8.5. However, above 70 °C, rapid loss in activity occurred and only 14% of activity remained at 80 °C. It was also reported that CGTase are more resistant to thermal denaturation in the
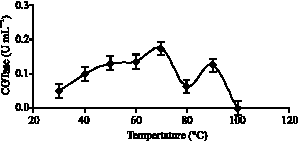 | |
| Fig. 2: | Effect of temperature on the activity of CGTase isolated from Bacillus MK6 after 10 min incubation |
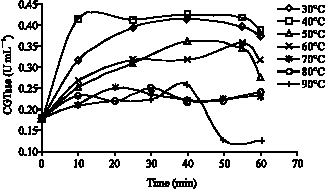 | |
| Fig. 3: | Temperature stability of crude enzyme at 60 min incubation time |
presence of its substrate, product and calcium ions, respectively. Although without the addition of any additives, the isolated enzyme has shown high stability over wide range of pH and temperature (Rita and Rajni, 2002).
It was reported that only CGTase from Bacillus steareothermophillus, or thermal tolerant organism, will show high stability over high temperature. These phenomenon may be due to different enzyme characteristic as compared to those enzymes isolated from mesophilic bacteria. In thermophilic bacteria, presence of abundant ionic bonding and hydrophobic bonding and also increase in hydrophobic interaction between protein subunit (enzyme) brings to the stability of thermal enzyme (Suelter, 1985).
Growth profile for CGTase enzyme production: Production of CGTase enzyme followed a similar enzyme production pattern (Fig. 4). The enzyme synthesis begins from the early exponential phase. However the maximum CGTase activity as measured by dextrinizing activity was obtained at 22nd-26th h of cultivation. From literature CGTase production by Bacillus cereus was at its peak during the 16-20 h of incubation period
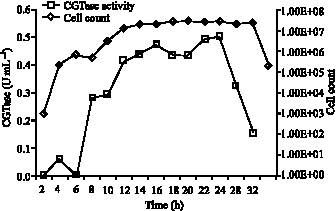 | |
| Fig. 4: | Production profile of CGTase by Bacillus MK6 |
(Jamuna et al., 1993). While for B. circulans alkalophillic sp. exhibits cyclizing activity after 40 h of growth with a long lag period (Mäkelä et al., 1988). Production of CGTase was observed only at the end of the stationary phase for alkalophillic Bacillus sp. G1 (Tien, 2001) and Bacillus agaradhaerens reaches stationary phase after 11 h of incubation with maximum enzyme production of 0.31 U mL-1 after 25 h cultivation (Rita and Rajni, 2002).
From the above comparisons, CGTase production from Bacillus MK6 was suggested as non-growth associated as the enzyme production was maximum at the late stationary phase. During the stationary phase, sporulation begins which appear to trigger enzyme synthesis for higher activity (Kabaivanova et al., 1999). CGTase production produced in shake flasks was studied by Thatai et al. (1999) and found to be at 7.5 U mL-1 after 24 h growth for alkalophillic Bacillus sp.
Nogrady et al. (1995) and Pocsi et al. (1998) suggested that extracellular CGTase are not involved in the degradation of starch in the exponential phase growth of the bacteria, because they are usually released into the culture medium when all the starches has been consumed. The enzyme was probably attached to the cell membrane during the exponential phase; it was retained between the cell membrane and the cell wall during the early stationary phase and was only released into the culture medium in the late stationary phase and during the cell lysis.
Optimization of CGTase production by RSM approach: Using completely randomized design, the experiments with different combination of sago starch concentrations, pH and agitation speed was assayed and calculated after Bacillus MK6 inoculum that was cultivated at 37 °C for 22 h. The results obtained were analyzed using analysis of variance (ANOVA) as appropriate to the experimental
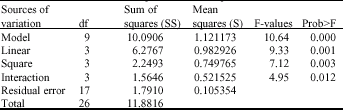
| Table 3: | Analysis of variance for the regression model of response, Y obtained from the response surface experiment |
 | |
design used. The regression (Eq. 4) obtained after analysis of variance gives the production of CGTase from Bacillus MK6 as a function of the different initial pH (X1), variables of sago starch concentration (X2, g L-1) and agitation (X3, RPM). All terms regardless of their significance was included in the following second-order polynomial equation:
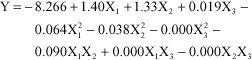 | (4) |
The regression model consisted of 1 offset, 3 linear, 3 quadratic and 3 interaction terms were generated when using the MINITAB version 13.0 software. The model was tested for adequacy and the quality of fit by the analysis of variance (Table 3). The variance ratio of the regression mean square gives a value of 10.64 [Tabulated F-value, F0.05 (9, 17) = 3.01]. As a practical rule, a model has a statistical significance when the calculated F-value is at least 3-5 times greater than the tabular value (Silva et al., 1999). Thus the regression is adequate in explaining the functional relationship between the response and the independent variables. The values of determination coefficient (R2 = 0.849, Adjusted R2 = 0.769) indicates that 15.1 - 23.1% of the total variations are not explained by the model.
The larger the magnitude of the t-value and the smaller the p-value, the more significant is the corresponding coefficient. For the first order effects, judging from the regression coefficient and t-values, it could be concluded that the sago starch percentage had the most significant effect on CGTase production, followed by agitation speed and pH value (Table 4).
For the first order effects, judging from the regression coefficient and t-values, it could be concluded that the sago starch (β2) concentration had the most significant effect on CGTase production, followed by agitation (β3) and pH (β1). The quadratic main effect of agitation (p<0.001) is the only significant factor. The pH and sago starch concentration were not significant at quadratic level. Therefore, agitation can act as a limiting factor and a little variation in their agitation speed may alter either growth or product formation rate or both to a considerable extent.
| Table 4: | Regression coefficients, t-value and p-value of second-order response surface equation for yield of CGTase enzyme, Y |
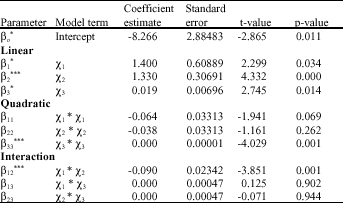 | |
| *: Significant at 5% level; **: Significant at 1% level; ***: Significant at 0.1% level, Linear: βo: Constant, β1: pH, β2: Sago Starch, β3: Agitation, Quadratic: β11: pH*pH, β22: Sago Starch*Sago Starch, β33: Agitation*Agitation, Interaction: β12: pH*Sago Starch, β13: pH*Agitation, β23: Sago Starch*Agitation | |
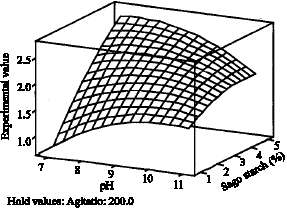 | |
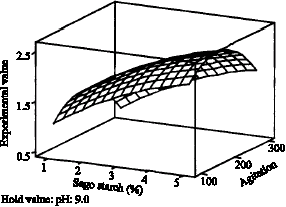
| Fig. 5: | Influence of pH and percent (%) sago starch on production of CGTase, Y when agitation was fixed at center point |
The non-additive effects of pH and percent (%) sago starch in Fig. 5 were due to the significant interaction between the two variables. The coefficient estimated for this interaction term has a negative sign (β12 = -0.090). Considering this interaction only, a negative sign of β12 may include that for an increase of the response, the coded levels of pH and % sago starch must not have the same sign, which means when there is increase in pH then there must be decrease in % sago starch. This may also indicate that the interaction term is not dominated by the other terms. Figure 6 and 7, shows no interaction between pH and agitation and % sago starch and agitation because effect of these pairs were addictive. At optimum point, the coded value of (pH) χ1 and (% sago starch) χ2 are 10.151 and 3.343, respectively.
The nature and concentration of the carbon-source are highly important in enzyme production from many organisms, especially when the carbon source also plays an important role in the enzyme induction (Gawande and
 | |
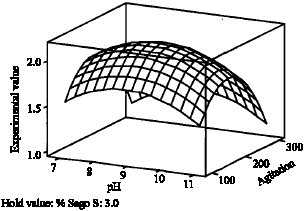
| Fig. 6: | Influence of percent (%) sago starch and agitation on production of CGTase, Y when agitation was fixed at center point |
 | |
| Fig. 7: | Influence of pH and agitation on production of CGTase, Y when agitation was fixed at center point |
Patkar, 2001). Comparisons with results shown by Khairizal et al. (2004), increase in sago starch concentration will bring to increase in CGTase production. In their research, it was found that the concentration of 1.48% of sago starch, was able to produce 84.32 U mL-1 of enzyme which is very high. From Fig. 6, although by increasing the concentration of sago starch will produce higher CGTase, this is however not economically feasible as pre heat treatment at a longer time needed to be done. Gawande et al. also commented that above a certain concentration of carbon substrate, when other nutrients are kept constant, catabolite repression may occur. This repression may occur due to limitation of other media components in the culture medium. Results reported for alkalophillic Bacillus sp. (ATCC 31007) reported that soluble starch concentration higher than 20-30 g L-1, resulted in low enzyme production.
For the effect of pH, optimum pH for CGTase production was at 10.151. This is due to the alkalophillic
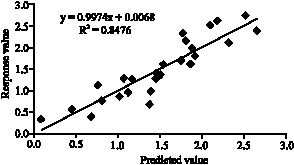 | |
| Fig. 8: | Correlation between observed response and predicted value |
nature of the isolated organism. However, when interact with higher concentration of sago starch, as shown in Fig. 7; surprisingly lower pH value is needed. This effect may be due to the high starch concentration resulting in high acid production, which decreases cell growth rate and enzyme production. This result is similar to those reported by Gawande and Patkar (2003).
Figure 8 shows the correlation between the predicted response and the observed response. It can be concluded that the experimental value is close to (R2 of 0.8476) the predicted experimental value given by the statistical software system. Therefore nature optimal values of the test variables are as follows: pH (χ1) = 10.151, % sago starch (χ2) = 3.343 and agitation (χ1) = 187 rpm. Using the optimal condition, the predicted response was 2.07 U mL-1. However the verification of results using the optimised medium was accomplished by carrying out shake flask experiments which gave final CGTase concentration of 2.56 U mL-1. Not only that these experimental finding are in close agreement with the model predictions, the experimental response value was 24% more than the predicted value. Furthermore, the experiments are controlled where numbers of experiments needed and the combination of parameters were clearly defined.
Production of cyclodextrin: From the optimal condition for cyclodextrin production obtained, the enzyme reactions were done in 2% slurried soluble starch, 0.1 M acetate buffer (pH 6.0) at 60 °C for 24 h. During incubation of CGTase enzyme in the presence of substrate, in the beginning of the reaction, the action of CGTase on starch will begin with the chaotic splitting of starch, followed by simultaneous cyclization. However, this chaotic splitting will also result in the formation of malto-oligosaccharides with various degrees of polymerization (Abelyan, 2001). CGTase from alkalophilic organisms have been known to be mainly β-CD producers while thermophilic and mesophillic CGTases produce a mixture of α-, β-and γ-CD, the α form being the dominant product (Abelyan et al., 1992).
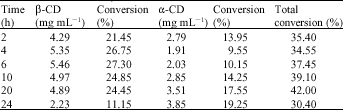
| Table 5: | Production of cyclodextrin at different incubation time |
 | |
| Soluble potato starch of 2% were used as substrate for cyclodextrin production and was detected using HPLC | |
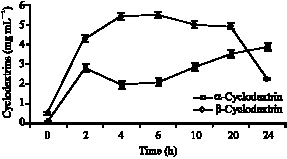 | |
| Fig. 9: | Time course of cyclodextrins production by Bacillus MK6`s CGTase |
Table 5 listed the production of CDS, in function of time using Bacillus MK6`s crude CGTase on 2% (w/v) slurried soluble potato starch. A maximum conversion of about 27.30% to 5.46 mg mL-1 of β-cyclodextrin was obtained after 6 h of incubation period and for α-cyclodextrin, the highest conversion of 19.25% corresponding to 3.85 mg mL-1 of was obtained after 24 h of incubation. However the maximum total conversion into both α-and β-cyclodextrin were obtained at 42% only after 20 h of incubation period.
The production profile (Fig. 9) shows that incubation of crude CGTase enzyme (2.5 U mL-1) per gram substrate yields mainly β-cyclodextrin (61.6% of the total cyclodextrins yield) with a α:β production ratio of 0.62:1. The concentration of β-cyclodextrin slowly decrease after 20 h incubation probably due to product degradation/decyclization in the reaction system (Kamarulzaman et al., 2004). The above results show similarity with those reported where the production of cyclodextrins particularly are β-cyclodextrin accelerated in the early hours of reaction (Ho et al., 2004; Rita and Rajni, 2002).
CONCLUSIONS
The effect of different carbon sources on CGTase production showed that enzyme production was highest when sago starch was used as carbon source. The use of hydrolyzed starch and simple sugars gave low yield of CGTase. Completely randomized design and response surface analysis were useful to determine the optimum levels of medium concentration and factors that significantly influence the production of CGTase from Bacillus MK6. The final composition of the defined medium to produce CGTase after the optimization step was as follows: 3.343% of sago starch; initial pH of 10.151 and agitation speed of 187 rpm. In theory, this optimized media produces 2.069 U mL-1 of CGTase while in practical, production of CGTase was at 2.56 U mL-1. Therefore this model was shown to adequately predict the optimization of CGTase production from Bacillus MK6. Results from the experiment done shown that CGTase from Bacillus MK6 were able to produce mainly β-cyclodextrin with 27.30% for 5.46 mg mL-1 β-cyclodextrin at 6th h, whilst α-cyclodextrin with only 19.25% for 3.85 mg mL-1 at 24th h; with a α:β production ratio of 0.62:1. No γ-cyclodextrin was detected.
REFERENCES
- Rosso, A.M., S.A. Ferrarotti, N. Krymkiewicz and B.C. Nudel, 2002. Optimisation of batch culture conditions for cyclodextrin glucanotransferase from Bacillus circulans DF 9R. Microl. Cell Fact., 1: 1-9.
Direct Link - Bart, A.V., C.M.U. Joost, W.D. Bauke and D. Lubbert, 2000. Engineering of cyclodextrin glycosyltransferase reaction and product specificity. Biochem. Biophys. Acta, 1543: 336-360.
Direct Link - Cao, X., Z. Jin, X. Wang and F. Chen, 2005. A novel cyclodextrin glycosyltransferase from an alkalophilic Bacillus species: Purification and characterization. Food Res. Int., 38: 309-314.
Direct Link - Gawande, B.N. and A.Y. Patkar, 2001. Purification and properties of a novel raw starch degrading-cyclodextrin glycosyltransferase from Klebsiella pneumoniae AS-22. Enzyme Microb. Technol., 28: 735-743.
CrossRefDirect Link - Ho, K.S., S. Mamot, H. Osman, K. Kamarulzaman, A.I. Fauzi, A.R. Roshanida, N.M. Nik Azmi and M.I. Rosli, 2004. Purification and characterization of cyclodextrin glucanotransferase from alkalophilic Bacillus sp. G1. Proc. Biochem., 40: 1101-1111.
Direct Link - Jamuna, R., N. Saswathi, R. Sheela and S.V. Ramakrishna, 1993. Synthesis of cyclodextrin glucosyl transferase by Bacillus cereus for the production of cyclodextrins. Applied Biochem. Biotechnol., 43: 163-176.
CrossRefDirect Link - Kamaruddin, K., R.M. Illias, S.A. Aziz, M. Said and O. Hassan, 2005. Effects of buffer properties on cyclodextrin glucanotransferase reactions and cyclodextrin production from raw sago (Cycas revoluta) starch. Biotechnol. Applied Biochem., 41: 117-125.
CrossRefDirect Link - Kaneko, T., T. Kato, N. Nakamura and K. Horikoshi, 1987. Spectrophotometric determination of cyclization activity of β-cyclodextrin-forming cyclomaltodextrin glucanotransferase. J. Jap. Soc. Starch Sci., 34: 45-48.
CrossRef - Mahat, M.K., R.M. Illias, R.A. Rahman, N.A.A. Rashid and N.A.N. Mahmood et al., 2004. Production of cyclodextrin glucanotransferase (CGTase) from alkalophilic Bacillus sp. TS1-1: Media optimization using experimental design. Enzyme Microb. Technol., 35: 467-473.
CrossRefDirect Link - Makela, M.J., T.K. Korpela, J. Puisto and S.V. Laakso, 1988. Nonchromatographic cyclodextrin assays: Evaluation of sensitivity, specificity and conversion mixture applications. J. Agric. Food Chem., 36: 83-88.
CrossRefDirect Link - Nogrady, N., I. Pocsi and A. Szentirmai, 1995. Cyclodextrin glycosyltransferase may be the only starch-degrading enzyme in Bacillus macerans. Biotechnol. Applied Biochem., 21: 233-243.
CrossRefDirect Link - Park, S.C., H.H. Park and S.H. Kim, 1989. A rapid screening method for alkaline β-cyclodextrin glucanotransferase using phenolphthalein-methyl orange-containing-solid media. Agric. Biol. Chem., 53: 1167-1169.
Direct Link - Penninga, D., B. Strokopytov, H.J. Rozeboom, C.L. Lawson, B.W. Dijkstra, J. Bergsma and L. Dijkhuizen, 1995. Site-directed mutations in tyrosine 195 of cyclodextrin glycosyltransferase from Bacillus circulans strain 251 affect activity and product specificity. Biochemistry, 34: 3368-3376.
CrossRefDirect Link - Pocsi, I., N. Nogrady, A. Liptak and A. Szentirmai, 1998. Cyclodextrins are likely to induce cyclodextrin glycosyltransferase production in Bacillus macerans. Folia Microbiol., 43: 71-74.
CrossRef - Martins, R.F. and R. Hatti-Kaul, 2002. A new cyclodextrin glycosyltransferase from an alkaliphilic Bacillus agaradhaerens isolate: Purification and characterisation Enzyme Microb. Technol., 30: 116-124.
CrossRefDirect Link - Stam, C.J., J.P.M. Pijn, P. Suffczynski and F.H.L. da Silva, 1999. Dynamics of the human alpha rhythm: Evidence for non-linearity. Clin. Neurophysiol., 110: 1801-1813.
CrossRefDirect Link - Thatai, A., M. Kumar and K.J. Mukherjee, 1999. A single step purification process for cyclodextrin glucanotransferase from a Bacillus sp. isolated from soil. Preparat. Biochem. Biotechnol., 29: 35-47.
Direct Link - Turnes, R.E. and S. Bahar, 1996. Production of cyclomaltodextrin glucanotransferase from an alkalophilic Bacillus sp. in rice and corn flakes. Acta Cientifica Venezolana, 47: 133-137.
Direct Link