
Muhammad Azhar Bashir
Horticultural Research Station, Bahawalpur-63100, Pakistan
Muhammad Akbar Anjum
University College of Agriculture, Bahauddin Zakariya University, Multan, Pakistan
Hamid Rashid
Agricultural Biotechnology Institute, National Agriculture Research Centre, Islamabad, Pakistan
Biotechnology
Year: 2007 | Volume: 6 | Issue: 4 | Page No.: 465-472







ABSTRACT
The study was conducted to initiate roots from in vitro derived shoots of six promising strains of jojoba i.e., PKJ-1 to PKJ-6. MS medium was supplemented with three auxins i.e., IBA, IAA and NAA each at the rate of 1.25, 2.5 and 5.0 mg L-1 in the one experiment and at the rate of 0.5, 1.0 and 1.5 mg L-1 in the other experiment. Different growth parameters pertaining to root formation were recorded. In the 1st experiment, the lowest concentration (1.25 mg L-1) of each auxin gave satisfactory results, the other two higher concentrations of auxins (2.5 and 5.0 mg L-1) caused callus induction. Efficacy of auxins was improved in the 2nd experiment as each auxin and its concentrations affected significantly the root characteristics of each strain. IBA and 0.5 mg L-1 were the most effective auxin and concentration, respectively. Among the strains, PKJ-3 performed the best of all, as it took the minimum time to root, produced maximum number of roots and attained longer primary root and higher percentage of rooted shoots in both experiments.
PDF Abstract XML References Citation
How to cite this article
Muhammad Azhar Bashir, Muhammad Akbar Anjum and Hamid Rashid, 2007. In vitro Root Formation in Micropropagated Shoots of Jojoba (Simmondsia chinensis). Biotechnology, 6: 465-472.
DOI: 10.3923/biotech.2007.465.472
URL: https://scialert.net/abstract/?doi=biotech.2007.465.472
DOI: 10.3923/biotech.2007.465.472
URL: https://scialert.net/abstract/?doi=biotech.2007.465.472
INTRODUCTION
Jojoba (Simmondsia chinensis Link. Schneider), the only member of family Simmondsiaceae, is a dioecious, long-lived evergreen perennial desert shrub. Its seeds contain about 50% saturated seed oil wax that is an alternative to sperm whale oil. It is also well known for its utilization in cosmetics, lubricants and pharmaceuticals etc. and is recently attracting the attention of research workers throughout the world. The plants have an exceptionally deep tap root system which helps to survive in drought conditions. Hence, it could be a prime plant species for introduction in desert areas of Pakistan like Cholistan, Thal and Thar. Jojoba plantations are established by using seeds, seedlings, rooted cuttings, or plantlets produced from tissue culture. Superior clones of jojoba, when used in field production, will allow a uniform, predictable plant growth and yield. Vegetative plant propagation from the selected plants can be performed by rooting the regular semi-hardwood cuttings (Palzkill, 1988). The use of simple node, double node and three node cuttings from different individuals of jojoba by applying different plant hormones will increase the total number of propagules obtained from a stock plants (Cao and Gao, 2003). However, the maximum number of possible propagules will still be limited to one or two thousands per year. Shoot tips established in tissue culture, however, will generally give rise to multiple shoots that can be rooted. Thus a single explant source, shoot tip, could conceivably provide thousands of new plantlets a year. Among the biotechnological techniques, micropropagation is an area of practical aspects for large scale mass multiplication of elite planting material. With the success of raising in vitro plants, the micropropagation has reached a commercial level in many plant species in recent years (Chandra and Mishra, 2003).
Kacker et al. (1993) incubated the micropropagated shoots of jojoba in the dark in a liquid half-strength MS medium containing 10 mg L-1 NAA for 72 h (for early root initiation on subsequent medium) and then transferred to half-strength MS rooting medium containing 2500 mg L-1 activated charcoal. They observed root initiation within a week. More than 80% shoots developed roots within a month. Rugini et al. (1993) placed explants of jojoba for rooting on modified Bourgin and Nitsch medium with or without 0.93 mg L-1 NAA. None of the explants rooted in the absence of NAA. Apostolo et al. (1996) obtained the 31.08% rooting after 70 days from micropropagated shoots cultured on half-strength MS medium containing IBA at the rate of 3.0 mg L-1. Benavides and Radice (1998) found that root induction on in vitro shoots of jojoba was improved after 60 days co-cultivation with Agrobacterium rhizogenes, with or without IBA added. Combined bacteria-IBA-cefotaxime significantly augmented root number and root length. Elhag et al. (1998) attempted rooting of micropropagated shoots of jojoba on various media (e.g., IAA, NAA, IBA, ½ MS or MS). The highest rate of rooting occurred on the medium containing IBA. Llorente and Apostolo (1998) reported that after 15 days of culture on modified MS medium supplemented with 3 mg L-1 IBA, 25% of the shoots had developed roots. The response of clones was highly variable; some exhibiting 75% root formation at 60 days, while others displayed little response. Sardana and Batra (1998) obtained complete plantlets with 1 or 2 thick roots per shoot after an incubation period of 35-40 days from shoot tips of jojoba cultured on MS medium supplemented with NAA and BA, both at 1.0 mg L-1. Agrawal et al. (1999) reported that nearly 80% of in vitro raised shoots produced roots with a pulse-treatment of 10 mg L-1 IBA (for 20 and 40 min, respectively) in female and male regenerants of jojoba. Khanam et al. (1999) obtained successful rooting from shoot bunches on rooting media supplemented with 0.2-2.0 mg L-1 BA, 2 mg L-1 IBA and 5000 mg L-1 activated charcoal. Roussos et al. (1999) treated the shoots produced during the proliferation stage with NAA, IBA and IAA to induce rhizogenesis. Rooting reached 64% in some treatments (10 mg L-1 IBA and 10 mg L-1 NAA). Gao and Cao (2001) reported that the best culture medium for in vitro rooting of branch segments of asceptic jojoba seedlings was half-strength MS + 1 mg -1 IBA + 1 mg L-1 IAA. Agrawal et al. (2002) reported that nearly 85% of the shoots produced roots with IBA treatment at 10 mg L-1 prior to transfer to MS medium containing 2 mg L-1 IBA + 5000 mg L-1 activated charcoal + 0.23 mg L-1 BA. Hassan (2003) achieved maximum frequency o f conversion of encapsulated buds of jojoba into plantlets on MS + 1 mg L-1 BAP + 40 mg L-1 adenine sulfate + 3 mg L-1 IAA. He observed after 35 days, a well developed shoots and roots on this medium. The average length of the shoots was 3.4 cm and that of the roots was 1.8 cm. Tyagi and Prakash (2004) reported that the pulse treatment of 10 mg L-1 IBA for 20 min caused in vitro rhizogenesis in 44-67% cultures of various jojoba genotypes tested.
The literature indicates that various auxins at different concentrations have been used for rooting of in vitro regenerated shoots of jojoba and different genotypes showed differential response. The present study is an endeavor to standardize auxins and their concentrations for root formation of in vitro derived shoots of jojoba and to evaluate the response of six jojoba strains to the various levels of these auxins.
MATERIALS AND METHODS
The experiments were conducted in Tissue Culture Laboratory of Agricultural Biotechnology Institute, National Agriculture Research Centre, Islamabad during 2005-2006.
 | |
| Fig. 1: | Shoot formation of PKJ-3 on MS + 1.25 mg L-1 BA |
Source of explants and its preparation: The nodal segments, 1.5-3.0 cm long with 1 or 2 nodes, excised aseptically from in vitro raised shoots of six jojoba strains, were used for shoot proliferation by culturing on MS medium (Murashige and Skoog, 1962) supplemented with 1.25 mg L-1 BA (Fig. 1). All cultural manipulation was carried out under Laminar Air Flow Hood.
Culture Medium and Culture Conditions: In vitro regenerated shoots were cultured on solidified MS medium containing 3% (w/v) sucrose and 0.7% (w/v) agar and supplemented with different auxins (NAA, IAA and IBA). The concentrations of these auxins varied according to treatments in each experiment as given below:
Experiment No. 1
| • | NAA at the rate of 1.25, 2.5 or 5.0 mg -1 |
| • | IAA at the rate of 1.25, 2.5 or 5.0 mg L-1 |
| • | IBA at the rate of 1.25, 2.5 or 5.0 mg L-1 |
Experiment No. 2
| • | NAA at the rate of 0.5, 1.0 or 1.5 mg L-1 |
| • | IAA at the rate of 0.5, 1.0 or 1.5 mg L-1 |
| • | IBA at the rate of 0.5, 1.0 or 1.5 mg L-1 |
The pH of the media was adjusted to 5.7 ± 0.1 using either 0.1N NaOH or 0.1N HCl prior to adding agar. Media were dispensed in 10 mL aliquots into culture tubes (2.5x15 cm), which were plugged with non-absorbent cotton wrapped in one layer of cheese cloth. Media were autoclaved at 121°C and 1.05 kg cm-2 for 20 min. The cultures were incubated under a 16 h photoperiod in cool, white fluorescent light of Philips tubes with a light intensity of 55 μmol m-2 sec-1 at 25 ± 2°C. Subculturing was carried out at monthly interval on fresh rooting medium with the same composition.
Data recording and statistical analysis: Each experiment was laid out in factorial Completely Randomized Design (CRD) with 3 replications and 3 factors i.e., auxins, concentrations and strains. Initially 5 shoots were cultured under each treatment per replication of each experiment, keeping one shoot in a test tube. The data were recorded on the following root parameters.
Number of days to root: The cultured shoots were observed vigilantly during the experimental period that continued for about three months. The count of days started from the date of culturing to the date of appearance of root primordia at the base of shoots. The days were averaged over number of shoots per treatment per replication.
Number of roots per shoot: The number of roots which arose from the shoot within 3 months was recorded and averaged over number of shoots per treatment per replication.
Length of primary root (cm): The length (cm) attained by the primary root that arose from the shoot within 3 months was recorded and averaged over number of shoots per treatment per replication.
Percentage of rooted shoots: At the end of 3rd month from first culturing, the percentage of rooted shoots was recorded by the given formula and averaged over treatment per replication.
Data collected were subjected to Fisher’s Analysis of Variance technique and treatment means were compared by Duncan’s Multiple Range test at 5% probability (Steele and Torrie, 1984).
RESULTS AND DISCUSSION
In the 1st experiment, all the three auxins remained effective only at the rate of 1.25 mg L-1, the other two concentrations of auxins i.e., 2.5 and 5.0 mg L-1 caused callus formation at the base of the shoots and no root formation was observed at these concentrations. Hence, the cultures containing these concentrations of auxins were excluded from data recording and statistical analysis.
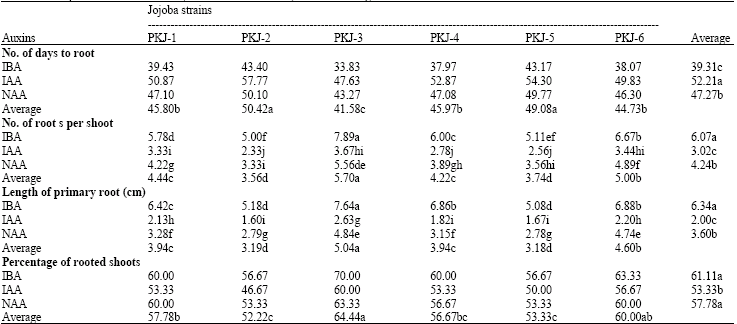
Number of days to root: The results of the 1st experiment indicated that strains and auxins had significant effect on the time required to root. However, the interaction between auxins and strains was statistically non-significant (Table 1). The cultured shoots of PKJ-3 took the minimum time to root (41.58 days) and differed significantly from cultured shoots of all other strains, while the shoots of PKJ-2 took the maximum time to root (50.42 days) that was not statistically different from that of PKJ-5 (49.08 days). The minimum time to root (39.31 days) was recorded by IBA added in the culture medium, while the maximum time to root (52.21 days) was recorded in case of IAA.
| Table 1: | Root parameters as affected by the strains and auxins, each at 1.25 mg L-1 |
 | |
| Means sharing similar letter(s) in a group are statistically non-significant at α = 0.05 (DMR test) | |
| Table 2: | Root parameters as affected by auxins, concentrations and strains |
 | |
| Means sharing similar letter(s) in a group are non-significant at α = 5% (DMR test) | |
| Table 3: | Number of days to root initiation as affected by auxinsxconcentrations interaction |
 | |
| Means sharing similar letter(s) are non-significant at α = 5% (DMR test) | |
| Table 4: | Number of days to root initiation as affected by concentrationsxstrains interaction |
 | |
| Means sharing similar letter(s) are non-significant at α = 5% (DMR test) | |
In the 2nd experiment, the effect of auxins, their concentrations and the strains was statistically significant (Table 2). IBA was more effective than the other two auxins as shoots took minimum days (43.30) to root in response to this auxin. The lowest concentration (0.5 mg L-1) of auxins remained better with 34.57 days for causing root initiation than that of the other two concentrations. The highest concentration (1.5 mg L-1) of auxins caused delay in root initiation up to 61.40 days. Among strains, the shoots of PKJ-3 initiated roots significantly earlier taking 43.47 days than that of the other strains, while the shoots of PKJ-2 took maximum time (52.81 days) for root initiation. The interaction between auxins and their concentration was significant (Table 3). The shoots in response to IBA at the rate of 0.5 mg L-1 took significantly fewer days (29.83), while the shoots in response to IAA at the rate of 1.5 mg L-1 took maximum days (66.89). The interaction between concentrations and strains (Table 4) was also significant. The shoots from PKJ-3 took 29.13 days to root initiation at 0.5 mg L-1 concentration, while the shoots of PKJ-2 took maximum days (66.36) to initiate roots at 1.5 mg L-1 concentration. Number of days to root initiation depends upon auxins used, their concentrations, type of media, strength of media, supplements of media, type of explant, genotypes, cultural conditions and various in vitro techniques as documented by previous researchers. The results obtained partially supported the findings of Llorente and Apostolo (1998) who observed rooting after 15 days of culture on MS + 3 mg L-1 IBA, but this high concentration of IBA caused only 25% rooting. The response of clones was highly variable as some clones exhibited root formation after 60 days. The results were also in conformity with the findings of Sardana and Batra (1998) who obtained complete plantlets with 1 or 2 thick roots per shoot after an incubation period of 35-40 days from shoot tips cultured on MS medium supplemented with 1 mg L-1 NAA and 1 mg L-1 BA. Previously by applying a different technique, Kacker et al. (1993) observed rooting within 30 days after culturing the micropropagated shoots in dark in a liquid half-strength MS medium containing 10 mg L-1 NAA for 72 hours and then transfer to ½ MS + 2500 mg L-1 activated charcoal medium. Similarly, Benavides and Radice (1998) who noted root induction after 60 days co-cultivation with Agrobacterium rhizogenes, with or without IBA added. However, Apostolo et al. (1996) observed rooting after 70 days from micropropagated shoots cultured on half-strength MS medium containing 3 mg L-1 IBA i.e. higher concentration possibly caused delay in rooting. Hassan (2003) observed well developed roots from encapsulated buds on MS + 1 mg L-1 BAP + 40 mg L-1 adenine sulfate +3 mg L-1 IAA after 35 days of culture.
Number of roots per shoot: It is apparent from Table 1 that the effect of IBA was significantly better with 6.07 roots per shoot than that of the other two auxins and the performance of PKJ-3 was the best of all the strains with 5.70 roots per shoot, while PKJ-2 trailed to the minimum with 3.56 roots and it was not statistically different from PKJ-5 with 3.74 roots per shoot. The interaction between auxins and strains significantly affected the number of roots per shoot due to differential response of strains to different auxins. Maximum number of roots (7.89) was produced by the shoots of PKJ-3 in the medium containing IBA at the rate of 1.25 mg L-1, followed by those of PKJ-6 (6.67) in the same medium. The minimum number of roots (2.33) was produced by the shoots of PKJ-2 in the medium containing IAA at the rate of 1.25 mg L-1 that remained statistically at par with those of PKJ-4 (2.78) and PKJ-5 (2.56).
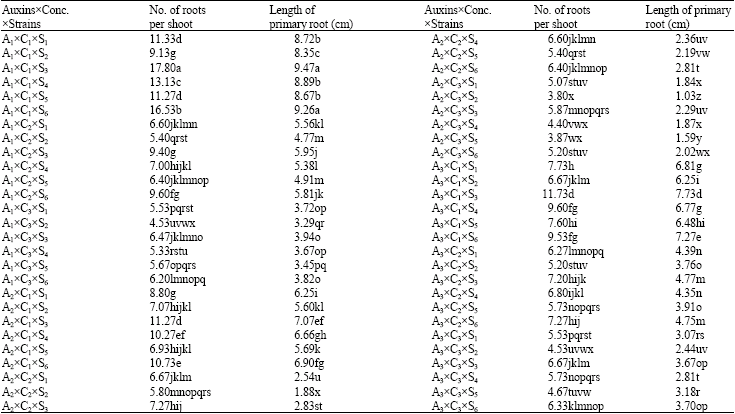
In the second experiment, auxins, their concentrations and strains significantly affected the number of roots per shoot (Table 2). Addition of IBA in the culture medium resulted in more number of roots (8.74) than that of the other two auxins (Fig. 2-4). Maximum number of roots (10.40) was produced at 0.5 mg L-1, while minimum number of roots (5.30) was produced at 1.5 mg L-1 concentration of auxins. PKJ-3 led the other strains with 9.30 roots, while PKJ-2 trailed with 5.79 roots per shoot. Interaction among three factors was statistically significant (Table 5). The shoots of PKJ-3 initiated the maximum number of roots (17.80) in response to IBA at the rate of 0.50 mg L-1 (Fig. 2) possibly due to early initiation of roots and sparing maximum time to enhance number of roots, while the shoots of PKJ-2 produced the minimum number of roots (3.80) in response to IAA at the rate of 1.50 mg L-1 possibly due to delay in root initiation; statistically this was not different from those of PKJ-4 and PKJ-5. The results are in conformity with the findings of Benavides and Radice (1998), Llorente and Apostolo (1998), Khanam et al. (1999), Agwawal et al. (2002) and Tyagi and Prakash (2004) who reported the prime role of IBA among auxins to affect number of roots, followed by that of NAA (Kacker et al., 1993; Sardana and Batra, 1998; Roussos et al., 1999). Differential response of genotypes to various concentrations of auxins was also reported by Llorente and Apostolo (1998), Agrawal et al. (1999) and Tyagi and Prakash (2004).
Length of primary root (cm): In the first experiment, the length of primary root was significantly affected by auxins and the strains (Table 1). As the roots were initiated earlier in the medium containing IBA, so it had more time to attain more root length (6.34 cm) than that of the other two auxins. Among the strains, the shoots of PKJ-3 led the other strains with maximum root length (5.04 cm) expressing significant clonal differences for this parameter, followed by PKJ-6 (4.60 cm), while the shoots of PKJ-5 trailed to the minimum root length (3.18 cm) that was statistically similar to that of PKJ-2 (3.19 cm).
 | |
| Fig. 2: | Root formation of PKJ-3 on MS + 0.5 mg L-1 lBA |
 | |
| Fig. 3: | Root formation of PKJ-3 on MS + 1.0 mg L-1 lBA |
 | |
| Fig. 4: | Root formation of PKJ-3 on MS + 1.5 mg L-1 lBA |
| Table 5: | No. of roots per shoot and length of primary root as affected by auxinsxconcentrationxstrains interaction |
 | |
| Means sharing similar letter(s) are non-significant at α = 5% (DMR test) | |
Interaction between two factors was statistically significant due to differential response of strains to different auxins. The longest primary root (7.64 cm) was produced by the shoots of PKJ-3 in the medium containing IBA at the rate of 1.25 mg L-1 due to early initiation of roots and sparing more time to attain maximum length of root in this medium, followed by PKJ-6 (6.88 cm) that was statistically similar to that of PKJ-4 (6.86 cm) for the same auxin. While the shortest primary root (1.60 cm) was attained by PKJ-2 in the medium containing IAA at the rate of 1.25 mg L-1 and it remained statistically at par with PKJ-4 (1.82 cm) and PKJ-5 (1.67 cm).
In the second experiment, the auxins, their concentrations and strains significantly affected the length of primary root (Table 2). IBA significantly enhanced the root length (5.98 cm) as compared with the other two auxins. The lowest concentration of auxins was more effective in increasing root length to the maximum (7.38 cm), while the highest concentration of auxins reduced it to the minimum (2.86 cm). The strain PKJ-3 attained longer primary root (5.30 cm), while PKJ-2 attained the shorter one (4.15 cm) than the other strains. Interaction among three factors was statistically significant due to differential response of strains to different concentration of auxins (Table 5). The shoots of PKJ-3 attained the longest primary root (9.47 cm) in the medium containing IBA at the rate of 0.5 mg L-1, followed by that of PKJ-6 with 9.26 cm root length on this medium, both were statistically at par with each other. As roots were initiated earlier in this medium, so it had more time to attain longer root than that of the other auxins tested at three concentrations. The shoots of PKJ-2 attained the shortest primary root (1.03 cm) in the medium containing IAA at the rate of 1.5 mg L-1. The results are in lines with the findings of Benavides and Radice (1998) who reported that IBA significantly increased root length. Similar findings were also reported by Llorente and Apostolo (1998), Sardana and Batra (1998), Khanam et al. (1999), Roussos et al. (1999), Agrawal et al. (2002) and Tyagi and Prakash (2004). However, Hassan (2003) achieved plantlets with average 1.8 cm long roots from encapsulated buds cultured on MS + 1 mg L-1 BAP + 40 mg L-1 adenine sulfate + 3 mg L-1 IAA.
Percentage of rooted shoots: In the first experiment, the auxins and strains had significant effect on the parameter under study (Table 1). The maximum percent rooted shoots (61.11) were recorded on the medium containing IBA that was statistically at par with that of NAA (57.78). As concern to the strains, PKJ-3 showed the highest percentage of rooted shoots (64.44) that was statistically alike with that of PKJ-6 (60.00), while PKJ-2 showed the lowest percentage of rooted shoots (52.22) that was not statistically different from that of PKJ-5 (53.33) and PKJ-4 (56.67). Interaction between the two factors remained statistically non-significant.
In the second experiment, percentage of rooted shoots was affected significantly by the auxins, their concentrations and strains (Table 2). Maximum percentage of rooted shoots (63.15) was credited to IBA, followed by NAA (58.89). However, both the auxins behaved statistically unlike. The shoots cultured on the media with lowest concentrations of auxins showed maximum rooting percentage (62.96), while those on the highest one resulted in the minimum (54.44). The shoots from PKJ-3 gained the highest rooting percentage (66.67), followed by PKJ-6 (62.22) but both of these strains differed significantly. The shoots from PKJ-2 gained the lowest rooting percentage (52.96) that was statistically at par with that of PKJ-5 (54.44). The differences among strains could be attributed to their specific genetic make up as observed previously in other root parameters. The results obtained from both experiments supported the findings of Elhag et al. (1998) who obtained the highest rate of rooting of micropropagated shoots on the medium containing IBA. The results were in accordance with the findings of Llorente and Apostolo (1998) who reported very meager percentage (25%) of rooted explants on MS +3 mg L-1 IBA (i.e., high concentration of IBA as compared to that used in 2nd experiment of the present study) and highly variable response of clones. Agraval et al. (1999), Agwaval et al. (2002) and Tyagi and Prakash (2004) enhanced rooting percentage by pulse treatment with higher concentration of IBA, but it could be an alternative technique. However, the results obtained contradicted to the findings of Roussos et al. (1999) who observed about 64% rooting on higher concentration of IBA & NAA which could be probably due to the different genotypes used. Improvement in rooting was observed by the addition of two auxins into half-strength MS (Gao and Cao, 2001) or on ½ MS + 3 mg L-1 IBA (Apostolo et al., 1996) or by the addition of activated charcoal (Khanam et al., 1999; Agwaval et al., 2002). By changing the technique, Kacker et al. (1993) also attained more than 80% rooted shoots within 30 days after incubation the micropropagated shoots in the dark in a liquid half-strength MS medium containing 10 mg L-1 NAA for 72 h and then transfer to half-strength MS rooting medium containing 2500 mg L-1 activated charcoal.
CONCLUSION
The efficacy of IBA over the other auxins used in the present study for in vitro root initiation and subsequent growth was proved as reported by earlier researchers. The auxins (IBA, IAA and NAA) at lower concentration (0.5 mg L-1) performed the best as compared to the other concentrations tested. The shoots of PKJ-3 showed significant supremacy over other strains with respect to in vitro root formation.
REFERENCES
- Agraval, V., S. Prakash, A. Altman, M. Ziv and S. Izhar, 1999. Differential hormonal requirements for clonal propagation of male and female jojoba plants. Proceedings of the 9th International Congress of the International Association of Plant Tissue Culture and Biotechnology, June 14-19, 1999, Jerusalem, Israel, pp: 25-28.
- Agraval, V., S. Prakash and S.C. Gupta, 2002. Effective protocol for in vitro shoot production through nodal explants of Simmondsia chinensis. Biol. Planta., 45: 449-453.
CrossRefDirect Link - Benavides, M.P. and S. Radice, 1998. Root induction in Simmondsia chinensis (Link.) Schneid. Using Agrobacterium rhizogenes. Biocell, 22: 109-114.
Direct Link - Cao, B. and H.D. Gao, 2003. Technology of cutting propagation of Simmondsia chinensis Link Schneider. J. Nanjing Forestry Univ., 27: 62-66.
Direct Link - Khanam, A., Y.B.N. Rao and S.A. Farooq, 1999. Standard in vitro protocol for high frequency mass micropropagation of jojoba [Simmondsia chinensis (Link) Schneider]. Adv. Plant Sci., 12: 361-366.
Direct Link - Llorente, B.E. and N.M. Apostolo, 1998. Effect of different growth regulators and genotype on in vitro propagation of jojoba. N. Z. J. Crop Hortic. Sci., 26: 55-62.
Direct Link - Murashige, T. and F. Skoog, 1962. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol. Plant., 15: 473-497.
CrossRefDirect Link - Roussos, P.A., A. Tolia-Marioli, C.A. Pontikis and D. Kotsias, 1999. Rapid multiplication of Jojoba seedlings by in vitro culture. Plant Cell Tissue Organ Cult., 57: 133-137.
CrossRefDirect Link - Rugini, E., A. Jacoboni and M. Luppino, 1993. Role of basal shoot darkening and exogenous putrescine treatments on in vitro rooting and on endogenous polyamine changes in difficult-to-root woody species. Scientia Horticulturae, 53: 63-72.
CrossRefDirect Link - Sardana, J. and A. Batra, 1998. In vitro regeneration of jojoba (Simmondsia chinensis): A plant of high potential. Adv. Plant Sci., 11: 143-146.
Direct Link - Steel, R.G.D. and J.H. Torrie, 1984. Principles and Procedures of Statistics: A Biometrical Approach. 2nd Edn., McGraw-Hill, New York, Pages: 633.
Direct Link - Tyagi, R.K. and S. Prakash, 2004. Genotypes and sex specific protocols for in vitro micropropagation and medium term conservation of jojoba. Biol. Plant., 48: 19-23.
Direct Link