
Abu Nasar Md. Aminoor Rahman
School of Agriculture and Rural Development, Bangladesh Open University, Gazipur-1705, Bangladesh
Ramli Bin Abdullah
Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
Wan Khadijah Wan Embong
Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences, University of Malaya, Lembah Pantai, 50603 Kuala Lumpur, Malaysia
Biotechnology
Year: 2007 | Volume: 6 | Issue: 3 | Page No.: 373-382







ABSTRACT
Experiments were conducted to develop an intracytoplasmic sperm injection (ICSI) protocol to produce goat embryos from in vitro matured heterogeneous oocytes. Two different protocols (‘Protocol 1’ or P-1 and ‘Protocol 2’ or P-2) were formulated and tested to develop a suitable one. Cumulus-oocyte complexes (COCs) were obtained either by LOPU (P-1) or by ovariectomy and slicing (P-2). Recovered COCs were cultured in a microdrop (50 μL) of HEPES-buffered in vitro maturation (IVM) medium with 10% oestrus goat serum (OGS) (P-1) or IVM medium without HEPES with 15% OGS (P-2) in presence of CO2 (5%) at 38.5°C either for 24 h (P-1) or 27 h (P-2). After IVM, COCs were denuded to select a mature oocyte (MII) and held in the incubator in presence of CO2 (5%) until ICSI. Motile sperm were selected from frozen-thawed buck semen by swim-up either in mDM (P-1) or mSOF (P-2) medium. A motile sperm was immobilized and injected, head-first, into the ooplasm of a MII oocyte. Sham injections were also performed as control. Injected oocytes were cultured in mSOF medium either with 10% (P-1) or 20% (P-2) goat serum in presence of CO2 (5%) at 38.5°C. A significantly higher (p<0.01) number of COCs was recovered with P-2 (17.5 COCs/donor) than P-1 (6.7 COCs/donor). However, the quality of the COCs was better with P-1 than P-2. A higher proportion of COCs were matured with P-2 (69.3%) than P-1 (55.5%), which was not statistically significant. A fertilization rate of 28.6% was obtained in P-1 without any cleavage. Fertilization rate (54.1%) was significantly higher (p<0.05) in P-2 with a cleavage rate of 18.2% and few morula-stage embryos were obtained (2.9%). In conclusion, better fertilization and embryo development rates were obtained with P-2 than P-1. Morula-stage embryos were obtained using ICSI technique from in vitro matured heterogeneous goat oocytes without any artificial activation and first reported in Malaysia.
PDF Abstract XML References Citation
How to cite this article
Abu Nasar Md. Aminoor Rahman, Ramli Bin Abdullah and Wan Khadijah Wan Embong, 2007. Goat Embryo Development from in vitro Matured Oocytes of Heterogeneous Quality Through Intracytoplasmic Sperm Injection Technique
. Biotechnology, 6: 373-382.
DOI: 10.3923/biotech.2007.373.382
URL: https://scialert.net/abstract/?doi=biotech.2007.373.382
DOI: 10.3923/biotech.2007.373.382
URL: https://scialert.net/abstract/?doi=biotech.2007.373.382
INTRODUCTION
Goat meat has gained high popularity among Malaysians due to their delicacy and nutritive value. However, country’s existing goat population can meet up less than 8% of local demand (Jamaluddin and Ng, 2005). Every year the country spent nearly 100 millions Malaysian Ringgit to import goat meat to cater the increasing demand. Therefore, the livestock industry was given a high priority in the Malaysia’s Third National Agricultural Policy (NAP3) (1998-2010). In order to make the government policy a reality, it is utmost important to adapt modern assisted reproductive technologies (ARTs) to boost livestock production. The conventional goat husbandry practice will lead to nowhere because of global competition. The application of ART enables to increase the rate of genetic progress (Nicholas, 1996). The ARTs allow animals of high genetic merit to produce more offspring than would be possible by natural breeding (Baldassarre and Karatzas, 2004). The ARTs those are widely used include artificial insemination; embryo transfer; multiple ovulation and embryo transfer; laparoscopic ovum pick-up (LOPU); in vitro maturation (IVM), in vitro fertilization (IVF) and in vitro culture (IVC) collectively known as in vitro production (IVP); intracytoplasmic sperm injection (ICSI); semen sexing; embryo sexing; cryopreservation of sperm, ova and embryos and embryo cloning. Among these, ICSI is the most recent innovation.
The ICSI technique has been the major breakthrough in the field of assisted reproduction among all micromanipulation techniques developed in the twentieth century. By far, it is the most widely used ART in human reproduction. ICSI is the mechanical insertion of a single sperm directly into the ooplasm of a mature, metaphase II (MII) oocyte using a microscopic needle. This technique bypasses many barriers of fertilization including the process of sperm penetration through cumulus and corona cells, zona pellucida and oolemma by directly depositing the sperm into the ooplasm. Therefore, ICSI offers an extremely interesting scope for studying some of the basic mechanisms underlying fertilization, e.g., oocyte activation, sperm decondensation and pronucleus formation. This technique could be useful in gender pre-selection (Hamano et al., 1999) and propagation of useful genes (Wang et al., 2003). It would be an effective technique to use genetically important but biologically inferior male gametes for procreating domestic and wild goat species through the application of ICSI (Keskintepe et al., 1997). Development of an ideal ART like ICSI would also improve the management of infertile or sub-fertile but valuable bucks.
Although ICSI is widely used, however till now, most of the ICSI investigations are concentrated in humans and laboratory animals. Among the domestic ruminant species, most of the ICSI research has been performed in cattle followed by sheep and has not been well studied in goat. The literature related to ICSI in goat is also scarce compared with those of other domestic animals and those of IVF studies in goat. A review of available literature revealed that studies related to ICSI in goat are very limited (Keskintepe et al., 1997; Wang et al., 2003; Zhou et al., 2004; Jiménez-Macedo et al., 2005; Jiménez-Macedo et al., 2006). Although many live births have been reported in cattle, sheep and other animals, only one report has been published on the birth of a male kid (Wang et al., 2003). Like in human and other animals, various aspects of ICSI technique have not been well studied in goat. It has been documented that advances in production of goat IVF embryos have been slow compared with those for other ruminant species like cow and sheep (Keskintepe et al., 1996). In Malaysia, the main obstacles to produce goat embryos by IVF include poor oocyte quality and limited oocyte sources for in vitro goat embryo production. The ICSI technique could be an option to overcome the obstacles and would be a useful tool for studying oocyte competence for embryo development more accurately. Therefore, ICSI could be a better option for the propagation of useful genes and rapid increment of goat population in Malaysia.
The objective of the present study was to develop a suitable protocol to produce ICSI-derived goat embryos from in vitro matured heterogeneous oocytes in Malaysia by comparing two different ICSI protocols, P-1 (developed based on existing IVF protocol of the lab) and P-2 (a modification of P-1).
MATERIALS AND METHODS
Study location and management of goats: The study was conducted in the Animal Biotechnology-Embryo Laboratory (ABEL), Institute of Biological Sciences (ISB), Faculty of Science, University of Malaya (UM) and Nuclear Transfer and Reprogramming Laboratory (NaTuRe), Institute of Research Management and Consultancy (IPPP), UM, Malaysia from August 2005 to July 2006. A total of 28 female goats were used, 17 for ‘Protocol 1’ (P-1) and 11 for ‘Protocol 2’ (P-2). The goats used in this study were housed in the ISB Mini Farm, ISB, UM throughout the experimental periods and were fed a diet consisting of good quality Napier grass and pellet feed (goat/sheep pellet, KMM Berhad, Malaysia) and given water ad libitum.
Chemicals, reagents and media: All the chemicals and media used in this study were from Sigma-Aldrich (St. Louis, MO, USA) unless otherwise stated. For all the media, the pH was adjusted to 7.2-7.4 and osmolarity to 280 mOsm kg-1.
Protocols: Two different protocols, P-1 and P-2, were developed starting from oocyte retrieval (OR), IVM, sperm preparation, ICSI until IVC. P-1 was developed based on the ABEL’s existing protocol for goat IVF (Phua, 2006; Amir, 2007) and P-2 was a modification of P-1. Different aspects of P-1 and P-2 have been summarized in Table 1.
Hormonal treatments and anesthesia: Experimental goats of P-1 were superovulated and their oestrus was synchronized using hormones, while no oestrus synchronization and superovulation was done for the goats of P-2. Oestrus synchronization and superovulation was done with a similar protocol used by (Baldassarre and Karatzas, 2004) with some modification. Briefly, the oestrus was synchronized with the insertion of a controlled internal drug release device (CIDR, progesterone, 0.3 g; Pharmacia and Upjohn Limited, Auckland, New Zealand) for 10 days combined with a luteolytic treatment of cloprostenol (Estrumate, 125 μg; Schering-Plough, NSW, Australia) 48 h prior to CIDR removal, which took place at the time of laparoscopic ovum pick-up (LOPU). Ovarian stimulation was obtained with gonadotrophin treatment consisting of Ovagen (FSH, 70 mg; Immuno Chemicals Products Ltd., Auckland, New Zealand) and Profasi (hCG, 1000 IU; Laboratories Serono, Switzerland) administered intramuscularly (i.m.) 36 h prior to LOPU. Goat for both P-1 and P-2 were off-fed for 24 h prior to LOPU or laparotomy (for ovariectomy).
| Table 1: | Differences between ‘Protocol 1’ (P-1) and ‘Protocol 2’ (P-2) |
 | |
Anesthesia was induced with i.m. administration of Xylazine hydrochloride (Ilium Xylazine-20, Troy Laboratories Pty Ltd., NSW, Australia) at the rate of 0.22 mg kg-1 body weight (bwt). Ketamine hydrochloride (Ketamil, Troy Laboratories Pty Ltd) (22 mg kg-1 bwt) was administered i.m. as maintenance doses every 20-30 min or as required.
OR and IVM: The COCs were obtained either by LOPU (P-1) or by ovary slicing after ovariectomy (P-2). A similar LOPU procedure as described by Baldassarre et al. (1994) and Baldassarre et al. (2003) was followed. The COCs were classified according to their cumulus-corona cell investments and morphology of the ooplasm (Table 2). Healthy COCs or naked oocytes with finely granulated and homogeneous ooplasm were selected for IVM. It should be noted here that all the COCs retrieved were not used in the present study as other researchers of the laboratory shared these for their research. Therefore, actual number of COCs cultured in the present study was lower than the total number of viable COCs. Groups of COCs (10) were matured in a microdrop (50 μL) of IVM medium consisting M-199 with Earle’s salt and sodium bicarbonate (Sigma, M-7528) supplemented with sodium pyruvate (20 mM, Sigma, P-3662), hCG (3 μg mL-1, Chorulon; Intervet), ovine FSH (3 μg mL-1, Ovagen, Immuno Chemicals Products Ltd), estradiol 17β (1 μg mL-1, Sigma, E-8875), gentamycin sulfate (25 μg mL-1, Sigma, G-1272) and oestrus goat serum (OGS).
| Table 2: | Classification of the recovered COCs according to the cumulus-corona cell investments and morphology of the ooplasm |
 | |
The differences between P-1 and P-2 were presented in the Table 1. The COCs were incubated at 38.5°C in a humidified atmosphere of CO2 (5%) in air under mineral oil (Sigma, M-8410).
Preparation of sperm and oocytes: Motile sperm fractions were selected by swim-up after motility of sperm was evaluated under an inverted microscope. Briefly, frozen buck semen (500 μL) from Jermasia breed (a breed developed in ABEL) of proven quality was thawed and incubated at 38.5°C in an humidified atmosphere of CO2 (5%) for 60 min in 2 mL of sperm medium supplemented with GS (10%). After incubation, supernatant (600 μL) was removed and centrifuged (200 x g) for 10 min. After 2 times washing the sperm pellet was resuspended in 1:1 sperm suspension: fresh medium containing heparin (50 μg mL-1, Sigma, H-3149) and incubated for another 15 min in a humidified atmosphere of CO2 (5%) at 38.5°C in the incubator.
After IVM duration, COCs were checked for maturation (Fig. 1A) and treated with 100 μL of hyaluronidase (80 IU mL-1) in HEPES-buffered medium. The cumulus-corona cells were carefully stripped off the zona pellucida by sequential pipetting through glass pipettes of 250 and 200 μm ID, respectively, under a dissecting microscope. Denuded oocytes were assessed for maturation at 20x magnification (eye-piece) in an inverted microscope and oocytes with a clear first polar body (PB) (Fig. 1B) were considered as mature oocytes (MII) and selected for ICSI. Oocytes without a visible PB were considered as immature and discarded. The selected MII oocytes were then cultured in the incubator in presence of CO2 (5%) at 38.5°C until ICSI.
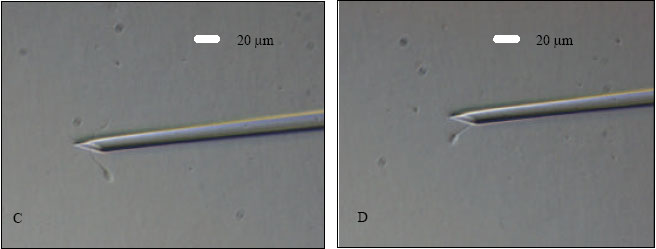
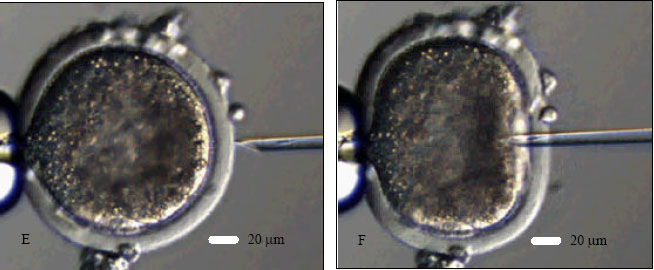
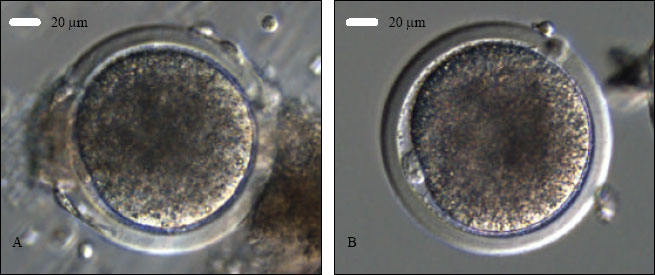 | |
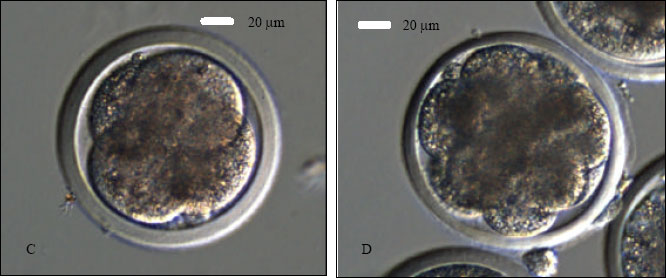
| Fig. 1: | Goat ooctye maturation, sperm catching and ICSI. A- a COC after 27 h of IVM to be denuded; B- a MII oocyte after denudation; C- a sperm is being immobilized in the PVP droplet; D- catching the immobilized sperm in tail-first manner; E- holding the oocyte with first PB at 6 o’clock and positioning the sperm at 3 o’clock for ICSI and F- injection of an immobilized sperm, head-first, into the ooplasm of a MII oocyte |
Micromanipulation technique for ICSI: The ICSI was performed on the lid of a small (35 mm) Falcon dish (Becton Dickinson Labware, Franklin Lakes, NJ, USA) under an Olympus inverted microscope (Model IX71) with Normarski optics fitted with Narishige hydraulic micromanipulators. The holding pipettes used in the experiments were made from borosilicate glass tubing of 1.0 mm Outer Diameter (OD) using a Sutter Micropipette Puller (Sutter Instrument Co., Novato, CA) in combination with a Narishige microforge and Narishige microgrinder (Narishige Scientific Instrument, Tokyo, Japan). The holding pipettes were pulled to an OD of 150 μm and then fire polished to create an ID of 30-40 μm. The pipette tips were bent to an angel of 35°. Injection needles used in P-1 were prepared in our laboratory using the same glass tubing and same instruments as mentioned for holding pipettes. For P-2 commercial injection needles were used (Table 1).
An aliquot (1-2 μL) of washed sperm suspension was added to an elongated microdrop (5 μL) of ICSI medium prepared on the left side of the ICSI dish. A motile sperm was aspirated into the injection needle head-first and it was brought into the PVP (10%) microdrop located at the center of the dish and immobilized by breaking its tail against the bottom of the dish by the tip of the injection needle (Fig. 1C). The immobilized sperm was aspirated again, tail-first, into the injection pipette (Fig. 1D). Each MII oocyte was positioned with its first PB at 6 or 12 o’clock on the holding pipette (Fig. 1E) in one of the four injection microdrop (5 μL sized) located on the right side of the ICSI dish. The injection pipette was at 3 o’clock position (Fig. 1E). The sperm was injected, head-first, into the ooplasm of the oocyte (Fig. 1F). During the injection, ooplasm was aspirated into the injection needle to ensure that the needle is in the ooplasm. The sperm was injected into the ooplasm with a minimum volume (<5 pl) of the medium. However, unlike human, monkey, mice or other laboratory animals it was too difficult to see the sperm inside the ooplasm as goat ooplasm is very dark due to a huge quantity of lipid materials. Sham injections (with vehicle medium only) were also performed as control in a similar manner. The oolemma was ruptured and the ooplasm was aspirated into the injection pipette and re-injected into the oocyte with a minimum volume of vehicle medium.
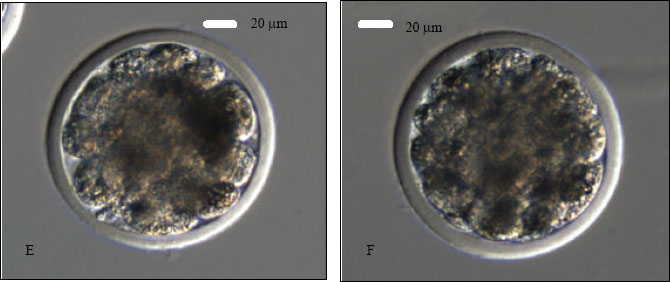
IVC: Injected or sham-injected oocytes were cultured in mSOF medium for the first 48 h in a humidified chamber of a CO2 incubator in presence of CO2 (5%) at 38.5°C. After 48 h of IVC, the old medium was replaced by freshly prepared mSOF medium and embryos were cultured up to 192 h in CO2 (5%) at 38.5°C without further medium change. The cultured embryos were assessed for cleavage under an inverted microscope. A zygote (fertilized oocyte) was confirmed by the presence of a second PB next to the first PB (Fig. 2A).
Statistical analysis: The statistical analyses were carried out by ANOVA using Statistical Packages for Social Sciences, version 11.5 for windows (SPSS, Inc., Chicago, IL, USA). Differences between treatments were evaluated with the Duncun multiple range test (DMRT). Data were expressed as mean±standard error of means (Mean±SEM) and p<0.05 was considered significant.
RESULTS
The results of goat OR using two different protocols, P-1 and P-2, are summarized in Table 3. Recovery rate of COCs per goat was significantly higher (p<0.01) with P-2 than P-1. However, good quality COCs were obtained with P-1 than P-2 as evidenced by the recovery of a significantly higher proportion (p<0.01) of viable COCs from P-1. When considering grades, a higher proportion (p<0.05) of COCs were recovered from grade C followed by grades A and B, while grade D yielded lowest proportion of COCs in P-1. On the other hand, a significantly higher proportion (p<0.01) of COCs were recovered from grade B followed by grades C and A, while grade D yielded lowest proportion of COCs in P-2.
The results of IVM of goat COCs have been depicted in Table 4. Although, a significantly higher (p<0.01) number of COCs were cultured in P-2 than P-1, however, it is evident that there was no significant difference in survival rate of COC between the two protocols. It is also evident that a higher proportion of oocytes were matured in P-2 compared with P-1, although the result was not statistically significant. On the other hand, when comparing MII versus immature COCs, no difference was found in P-1, whereas a very significantly higher (p<0.001) proportion of COCs became MII over immature COCs in P-2.
As shown in Table 5, there was no significant difference in the survival rate of the injected oocytes between P-1 and P-2. Compared with P-1 (28.6%), a higher (p<0.05) fertilization rate was obtained with oocytes in P-2 (54.1%). There was no cleavage obtained with P-1, however, 18.2% of P-2-derived intact injected oocytes were cleaved and finally some of them developed to morula stage (Fig. 2B-F), which was statistically significant (p<0.05). Injection without a sperm (sham injection) did not produce any zygote with P-1-derived MII oocytes.
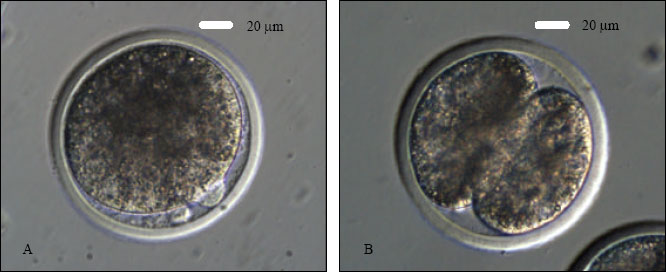 | |
| Fig. 2: | ICSI-derived goat embryos at various stages of development. A- a zygote represented by the presence of a second PB next to the first PB at 16-18 h of IVC; B- a 2-cell embryo after 36 h of IVC; C- a 4-cell embryo at 48 h of IVC; D- a 8-cell embryo at 72 h of IVC; E- a 16-cell embryo at 96 h of IVC and F- a compacting morula at 120 h of IVC. Note: all the embryos have been produced with Protocol 2 |
| Table 3: | Recovered COCs using two different protocols with special reference to grading of viable COCs |
 | |
| P-1 (LOPU) = Oestrus synchronized and superovulated goats, P-2 (ovariectomy) = Non-stimulated goats; number of COCs was based on per goat; n = No. of oocytes used, a,b Means within the same column with different superscripts differed significantly (p<0.01), x,y,z Means within the same row with different superscripts differed significantly (p<0.05 for P-1 and p<0.01 for P-2) | |
| Table 4: | In vitro maturation of goat oocytes using two different protocols |
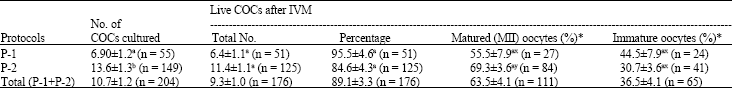 | |
| n = No. of oocytes used, *Percentage of MII and immature oocytes were calculated from total live oocytes after IVM, a,b Means within the same column with different superscripts differed significantly (p<0.01), x,y Means within the same row with different superscripts differed significantly (p<0.001) | |
| Table 5: | Fertilization and embryo development rate of ICSI and sham-injected oocytes using two different protocols |
 | |
| Mean number of injected oocytes was based on per replication; n = No. of oocytes used, *Percentage of zygotes and embryos were calculated from intact oocytes, a.bMeans within the same column with different superscripts differed significantly (p<0.05) | |
Although only one zygote was formed with P-2-derived oocytes, however, no embryo development at first cleavage or beyond was observed in sham-injected groups with any of the experiments.
DISCUSSION
To establish a suitable ICSI protocol for the production of goat embryos from in vitro derived oocytes, two different sets of protocol starting from OR until IVC were formulated and tested. The present piece of study was presumably the first successful report of ICSI-derived goat embryo production in Malaysia as well as in the tropics. In the present study, a simple and conventional technique was used rather than a highly sophisticated and complicated one to succeed in producing ICSI-derived goat embryos from COCs of heterogeneous qualities including low quality COCs.
In the present study, the result of LOPU (P-1) was in agreement with the findings of Baldassarre and Karatzas (2004) who mentioned LOPU almost always results in >5 oocytes/donor. However, OR result of the present study was lower than the average recovery of 13.4 oocytes/donor of the same group (Baldassarre et al., 2002; Baldassarre and Karatzas, 2004). Although, the reason could not be known, however, they used standard breed of goat (Nigerian dwarf) and a slightly different superovulation and oestrus synchronization protocol, whereas in the present study, goats of heterogeneous breeds and ages were used and breed and age of some goats were unknown. Anyway, a significantly higher (p<0.01) number of COCs were recovered with P-2 than P-1 in the current study. However, P-1 (by LOPU) yielded significantly higher (p<0.01) proportion of viable COCs than P-2. Again, a significantly higher proportion of C (p<0.05) and B (p<0.01) grade COCs were recovered, respectively from P-1 and P-2.
From the IVM data, it is clear that P-2 provided a higher proportion of MII oocytes compared with P-1 (69.3 vs 55.5%) although result was not statistically significant. However, when comparing MII versus immature COCs, no significant difference was found with P-1, but a very significantly higher (p<0.001) proportion of MII COCs was obtained than immature with P-2. Although, the reason for higher proportion of MII compared with immature COCs with P-2 could not be separated from effects of various factors, however, this could be partly due to the exclusion of HEPES from the medium and increment of IVM duration from 24 h (P-1) to 27 h (P-2). Generally, HEPES is used to maintain the pH of the medium (7.2-7.4) outside the CO2 incubator. The use of HEPES in IVM medium inside the CO2 incubator could lower the pH. However, there have been reports of obtaining good maturation rate of goat oocytes using HEPES-buffered IVM medium (Keskintepe et al., 1997; Wang et al., 2003). Therefore, HEPES (20 mM) was added in the IVM medium for culturing COCs in P-1. Culturing goat COCs for 27 h at 38.5°C better maturation rates were obtained by other groups (Wang et al., 2003; Jiménez-Macedo et al., 2005; Jiménez-Macedo et al., 2006). However, compared with other groups, lower maturation rate was obtained in the present study especially with P-1. The reason for lower maturity in the present study probably, in part, due to the quality of the cultured COCs. While other groups cultured quality COCs with at least 4 (Jiménez-Macedo et al., 2005; Jiménez-Macedo et al., 2006), 2 (Keskintepe et al., 1997) and 1-2 (Wang et al., 2003) layers of CC, in the present study, COCs of heterogeneous quality was cultured including COCs with <1 complete layer of CC or naked oocyte due to a shortage of goat oocytes. It is to be mentioned here that in Malaysia female goats are highly priced and rarely slaughtered because the country has huge shortage of breeding stock. For this reason, in the present study there was a scarcity of goat oocytes, which forced the researchers to use lower quality oocytes.
The present study was undertaken to develop a suitable ICSI protocol for goat embryo production under the local setting of ABEL, UM, Malaysia to boost country’s goat population. Compared with P-1, a better fertilization rate was obtained with P-2 (P-1 vs. P-2 = 28.6 vs 54.1). Although the reason for lower fertilization with P-1 was not identified, however, it could be speculated that factors such as larger needle diameter, improper immobilization of the sperm, injecting the oocyte without a spike could be responsible for this. As it was reported earlier that egg yolk cryoprotectant used to cryopreserve sperm change the surface or membrane structure of the sperm (Simplicio et al., 1997), therefore, no additional capcitation treatment except heparin was provided and the sperm was not manipulated vigorously in P-1 as frozen-thawed sperm was used. Besides, needles of bigger diameter lacking a sharp spike was used, as there has been report of obtaining better fertilization rate using needle without a spike (Hamberger et al., 1995). Therefore, in P-2, the technique was improved by manipulating sperm vigorously and using needle with a smaller diameter having sharp spike. A significantly higher (p<0.05) fertilization rate was obtained with P-2. Although, no blastocyst was obtained in the present study, however, a small proportion of embryos developed to morula-stage (2.9%). This is definitely a preliminary success of producing goat embryo with ICSI using oocytes of heterogeneous and lower quality.
The oolemma of goat oocyte is very elastic, it is very difficult to inject a sperm into the ooplasm without damaging the oocyte membrane and cytoplasmic structure. Reducing the mechanical damage to oocyte during injection and effectively activating oocyte are two important factors. To overcome the difficulty of sperm penetration through the oolemma and improve the efficiency of ICSI, a Piezo-driven procedure was used in goat with a 97% oocyte survival rate (Wang et al., 2003). This process also eliminated the need for use of chemicals such Ca-ionophore (Keskintepe et al., 1997), Ca-ionophore and DMAP (Zhou et al., 2004; Jiménez-Macedo et al., 2005) to activate goat oocytes after ICSI, which may be detrimental to the oocytes and embryo if not used in proper concentration. However, sophisticated equipment and extensive manipulating experience are needed to use Piezo-driven micromanipulator. It was noticed in the current study that the main cause of oocyte degeneration was the failure of the injected oocyte to heal the damage caused by the mechanical insertion of the needle, especially when using a needle of larger diameter without a spike (P-1). This problem was minimized by using a needle with a smaller diameter having a sharp spike that is slightly larger than the diameter of the goat sperm (P-1).
One of the important things to initiate the sperm nucleus decondensation and oocyte activation is the mutual interaction of the ooplasmic factors and sperm sub-membrane components (Perry et al., 1999). Permeabilization of the sperm membrane is a prerequisite of this process during natural fertilization (Kasai et al., 1999) or IVF. The ICSI technique bypasses sperm membrane breakdown that occurs during the fertilization as sperm penetrate the oolemma. Therefore, a number of process or treatments have been employed to break the sperm membrane before ICSI in goat, viz. breaking the sperm tail with a injection micropipette (Keskintepe et al., 1997), cutting the tail with the holding pipette (Wang et al., 2003), treating sperm with Triton X-100 (Zhou et al., 2004) or heparin plus Ca-ionophore (Jiménez-Macedo et al., 2006). Using these techniques fertilization rates in goat have been significantly improved. However, cutting the tail of sperm is not easy as it requires extensive manipulative experience and chances of oocyte damage with an unpolished holding pipette can not be ignored.
Culture conditions have a greater effect on fertilization, cleavage and subsequent embryo development after ICSI. Compared with other groups engaged in goat ICSI research, a comparatively lower cleavage (18.3%) and embryo development (2.9-18.2%) rate was achieved in the present study. Using a complex and sequential media consisting of mTALP medium with OGS (20%) for first 24 h, mKOSM with BSA (0.3%) from 24-96 h and mKOSM with fatty acid free BSA (1%) from 96-192 h, Wang et al. (2003) obtained a higher cleavage rate (71-90%) and morula (41%) or blastocyst (35%) production. However, using a slightly modified SOF medium, Jiménez-Macedo et al. (2005), Jiménez-Macedo et al. (2006) and Keskintepe et al. (1997) obtained 73.4, 60.3-75.0 and 67.0% cleavage rate, respectively. It is to be noted here that although a lower cleavage rate was obtained in the present study, however, like other groups sperm was not activated with heparin+ionomycin (Jiménez-Macedo et al., 2006) before ICSI and also oocytes were not activated with ionomycin+DMAP (Jiménez-Macedo et al., 2005) or ionomycin alone (Keskintepe et al., 1997) after ICSI or a Piezo micromanipulator was not used to facilitate sperm injection (Wang et al., 2003). On the other hand, using COCs of heterogeneous and lower quality, conventional ICSI technique, simple media and untreated sperm and oocyte, the results of the present study are still satisfactory and comparable with Jiménez-Macedo et al. (2005) who obtained only 1.5% cleavage from good quality COCs without activation of sperm or oocytes, although with prepeubartal goat oocytes. In the present study, no fertilization (1 oocyte fertilized with P-2) or cleavage was seen with sham-injection, which is in agreement with Wang et al. (2003).
The synchronization of ooplasmic and nuclear maturation is essential for obtaining better fertilization and cleavage rate. In the present study, it was not known whether all mature oocytes obtained ooplasmic maturation with the nuclear maturation. Therefore, lack of synchronization of ooplasmic and nuclear maturation for obtaining better fertilization and cleavage rate in the current study could not be ignored.
Finally, although the objective of producing ICSI-derived goat embryos from in vitro matured heterogeneous oocytes was achieved up to morula-stage with Protocol 2 in the present study, however, the protocol still needs to be improved to get better results. Future studies of the laboratory will focus on some important aspects, such as optimization of IVM medium, sperm and oocyte activation, ICSI according to oocyte grades, PB position etc to increase fertilization, cleavage and embryo development rate.
ACKNOWLEDGMENTS
The authors wish to thank Dr. S.L. Liow, Embryonics International, Singapore for critically reviewing the manuscript. The ‘OR Team’ members are highly acknowledged for their constant help during LOPU. We are also grateful to Islamic Development Bank (IDB) for providing an IDB Merit Scholarship to the first author. This work was supported by grants from MOSTI Special Project (grant no 01-02-03-0696) and IPPP (grant No. Vote F-0179/2004D and 0145/2005D).
REFERENCES
- Baldassarre, H., B. Wang, N. Kafidi, C. Keefer, A. Lazaris and C.N. Karatzas 2002. Advances in the production and propagation of transgenic goats using laparoscopic ovum pick-up and in vitro embryo production technologies. Theriogenology, 57: 275-284.
CrossRef - Baldassarre, H., C. Keefer, B. Wang, A. Lazaris and C.N. Karatzas, 2003. Nuclear transfer in goats using in vitro matured oocytes recovered by laparoscopic ovum pick-up. Clon. Stem Cells, 5: 279-285.
Direct Link - Brackett, B.G. and G. Oliphant, 1975. Capacitation of rabbit spermatozoa in vitro. Biol. Reprod., 12: 260-274.
CrossRefDirect Link - Hamano, K., X. Li, X.Q. Qian, K. Funauchi, M. Furudate and Y. Minat, 1999. Gender preselection in cattle with intracytoplasmically injected, flow cytometrically sorted sperm heads. Biol. Reprod., 60: 1194-1197.
CrossRef - Hamberger, L., A. Sjogren, K. Lundin, B. Soderlund and L. Nilsson et al., 1995. Microfertilization techniques- The Swedish experience. Reprod. Fertil. Dev., 7: 263-268.
PubMedDirect Link - Kasai, T., K. Hoshi and R. Yanagimachi, 1999. Effect of sperm immobilisation and demembranation on the oocyte activation rate in the mouse. Zygote, 7: 187-193.
CrossRefPubMedDirect Link - Keskintepe, L., G.C. Luvoni, S.J. Rzucidlo and B.G. Brackett, 1996. Procedural improvements for in vitro production of viable uterine stage caprine embryos. Small Rum. Res., 20: 247-254.
Direct Link - Keskintepe, L., P. Morton, S.E. Smith, M.J. Tucker, A.A. Simplicio and B.G. Brackett, 1997. Caprine blastocyst formation following intracytoplasmic sperm injection and defined culture. Zygote, 5: 261-265.
PubMed - Nicholas, F.W., 1996. Genetic improvement through reproductive technology. Anim. Reprod. Sci., 42: 205-214.
CrossRefDirect Link - Perry, A.C., T. Wakayama and R. Yanagimachi, 1999. A novel trans-complementation assay suggests full mammalian oocyte activation is coordinately initiated by multiple, submembrane sperm components. Biol. Reprod., 60: 747-755.
Direct Link - Simplicio, A.A., B.G. Brackett and L. Keskintepe, 1997. Use of cryopreserved spermatozoa for caprine in vitro fertilization (IVF). Theriogenology, 47: 299-299.
CrossRefDirect Link - Takahashi, Y. and N.L. First, 1992. In vitro development of bovine one-cell embryos: Influence of glucose, lactate, pyruvate, amino acids and vitamins. Theriogenology, 37: 963-978.
PubMed - Wang, B., H. Baldassarre, J. Pierson, F. Cote, K.M. Rao and C.N. Karatzas, 2003. The in vitro and in vivo development of goat embryos produced by intracytoplasmic sperm injection using tail-cut spermatozoa. Zygote, 11: 219-227.
PubMed - Zhou, J.B., Y.G. Wu, D. Han, L.Q. Liu and X.W. Tan et al., 2004. [Effects of sperm and oocyte quality control on Intracytoplasmic Sperm Injection (ICSI) in goats]. Shi Yan Sheng Wu Xue Bao, 37: 367-374, (In Chinese).
PubMed