
Ahmad H. Al-Gabbiesh
University of Jordan, Jordan
Dhia S. Hassawi
University of Jordan, Jordan
Mutasem O. Taha
University of Jordan, Jordan
Fatma U. Afifi
University of Jordan, Jordan
Biotechnology
Year: 2007 | Volume: 6 | Issue: 2 | Page No.: 218-224







ABSTRACT
Calli cultures from Iris atrofusca and I. petrana were propagated using standard Murashige and Skoog (MS)- and quinine treated MS-media. The propagated calli were analysed for their volatile oil contents. An irone-type volatile substance was identified by GC-MS analysis for the first time in both threatened Iris species. When calli cultures were grown in quinine treated MS-medium, quinine induced a dose-dependent decrease/cessation of the volatile compound. Furthermore, quinine caused gradual reduction of the calli biomass propagated from both Iris species.
PDF Abstract XML References
How to cite this article
Ahmad H. Al-Gabbiesh, Dhia S. Hassawi, Mutasem O. Taha and Fatma U. Afifi, 2007. Effect of Increasing Concentrations of Quinine on the Volatile Compounds in Cultured Tissues of Iris atrofusca Baker and Iris petrana Dinsmore. Biotechnology, 6: 218-224.
DOI: 10.3923/biotech.2007.218.224
URL: https://scialert.net/abstract/?doi=biotech.2007.218.224
DOI: 10.3923/biotech.2007.218.224
URL: https://scialert.net/abstract/?doi=biotech.2007.218.224
INTRODUCTION
Plant tissue culture in its broad sense refers to the in vitro culturing of all plant parts (single cell, tissue or organ) under aseptic conditions (Slater et al., 2003). Plant cell and organ cultures have been employed in the investigation of the physiological, biochemical, genetic and structural problems related to plant growth and development (Rasol et al., 1999). Tissue culture provided a new experimental approach thus avoiding the complex interactions of biological processes in intact plant. Through this in vitro technique, important advances have been made to a better understanding of cell division, expansion and differentiation in plants (Evans, 2002).
Volatile oils produced from different pathways have been isolated from various plants using steam distillation and characterized using mainly GC and MS (De Hoffmann et al., 1996; Robbers et al., 1996; Magiatis et al., 2001; Prokopios et al., 2001; Evans, 2002). Several authors have studied the production of essential oils in cultured tissues propagated from different plants, such as in oregano and tansy (Svobada et al., 1995; Sokmen et al., 2004), Eucalyptus camaldulensis (Giamakis et al., 2001), Mentha spicata (Abou-Mandour and Binder, 1998), Citrus aurantifolia (Agrawal et al., 1991), Coleonema album (Berger et al., 1990), Rosmarinus officinalis (Shervington et al., 1997) and Teucrium polium (Aburjai et al., 2006) Intensive studies have been conducted to elucidate the effect of different growth regulators (kinetin, 1-naphthaleneacetic acid (NAA) and 2,4-dichlorphenoxyacetic acid (2,4-D)) and culture conditions on the production of the biomass formation and volatile oils as secondary metabolites from callus cultures of various plant species (Cotton et al., 1991; Chaturvedi et al., 2001; Magiatis et al., 2001; Prokopios et al., 2001; Biemelt et al., 2004).
The topic of animal based toxicity testing has recently received great interest. In October 2003, the European Commission proposed that instead of continuing to perform animal tests, relevant and reliable animal testing strategies should be implemented (Sauer, 2004). Unless an efficient substitution for animals became available, this drive for the avoidance of live animal use in toxicity studies would pose a major obstacle in scientific research. Hence cultivated cells could be a suitable alternative. Put together, these prompted us to evaluate the possibility of employing plant tissue cultures (calli) as experimental models for the study of intoxications in the future. Since to the best of our knowledge, no similar studies have been published to date, the present study can be regarded as one of the first attempts carried out in this aspect.
The volatile oils can be easily detected and quantified, so they may provide an excellent and quick tool for assessing cellular response to different pharmaceutical entities. Moreover, it is well known that the biosynthesis of volatile oils involves large number of enzyme-systems, naturally expected to be highly sensitive towards different classes of possible intoxicants, such as the anti-malarial agent quinine (Robbers et al., 1996; Fullerton, 1998). Subsequently, to test the effect of quinine, we selected two volatile oil containing threatened plant species of Jordan (Iris atrofusca Baker and I. petrana Dinsmore).
In the present study, the aim was to propagate calli from both threatened Iris species, to determine the main volatile constituent (s) from the steam distilled fraction of the intact plants and propagated calli using GC-MS and to observe the fate of the volatile substance (s) upon addition of different concentrations of quinine into the growth media. Furthermore, to the best of our knowledge we are the first to report the presence of irone type volatile substances in both studied Iris species.
MATERIALS AND METHODS
Plant material: Both Iris species were collected from Wadi-Shueib, Tafila and Al-Ardah, Jordan in Ferbuary 2003 and were identified using descriptive references and by Mr. Ibrahim Al-Khader, plant taxonomist (Al-Eisawi, 1986). A voucher specimen of each species has been kept in the Faculty of Agriculture/Department of Biotechnology at Al-Balq’a Applied University-Salt-Jordan.
Equipments: All tissue culture experiments were carried out under laminar air flow cabinet (Esco, Model 1851, Singaphore). Calli were incubated in a Precision incubator (Model 815, USA). Callus excision and photography were performed using dissecting light microscope (Nikon, Model 2080, Japan) attached to digital camera (Nikon, Model 3030, Japan). GC analyses were performed using Hewlett Packard GC apparatus model 5890 series II with attached Flame Ionization Detector (FID) and Hewlett Packard 3395 integrator. Mass analyses were carried out using GC/MS (Shimadzu-GCqp2010, Japan) with electron impact as ionization mode.
Calli propagation: The flower bases of sterilized I. atrofusca and I. petrana flowers were excised (Eunice, 1995). Every 7 excisions were transferred to the surface of M and S medium (12 mL) placed in 9 cm Petri dishes containing 2,4-D (1.0 mg, 4.5 μM), kinetin (0.11 mg, 0.50 μM), NAA (0.84 mg, 4.5 μM) and quinine sulfate in different concentrations. Five Petri dishes were allocated for each Iris species exposed to a particular quinine concentration. A total of six different quinine concentrations were prepared within the growth media: 0.01, 0.02, 0.03, 0.04, 0.05 and 0.09% (w/v) in addition to an untreated control. The experiments were run in quadruplets. Hence 140 explants were worked out for each Iris species. After incubation for five weeks under dark condition at 25°C, the produced calli were excised, weighted, photographed, crushed and submitted to micro-steam distillation.
Steam distillation and GC conditions for volatile oil analysis: Finely crushed calli (200 mg) or intact plant material were submitted to micro-steam distillation and the obtained volatile substances were subjected to TLC and GC analysis (Wagner and Bladt, 1996). One micoliter-samples were injected into the GC injection port preheated at 200°C. The samples were eluted in an HP-5 (5% diphenyl-, 95% dimethyl-polysiloxane) column employing dry nitrogen gas as elution carrier gas at a flow rate of 1 mL min-1. The temperature of the GC column was adjusted according to the following program: 45°C over 3 min then heated at 4°C per minute up to 250°C. Eluted samples were detected using FID apparatus.
GC-MS conditions for volatile oil analysis: Samples with detectable volatile oils, i.e., control sample and samples exposed to 0.01, 0.02, 0.03 and 0.04% (w/v) quinine sulfate, were analyzed by GC-MS. Each 1 μL sample was injected into the GC injection port preheated at 90°C. The samples were eluted in a Treb-Merck-αS column employing dry nitrogen gas as elution carrier gas at a flow rate of 1 mL min-1. The temperature of the GC column was adjusted according to the following program: 45°C over 3 min then heated at 4°C per minute up to 25°C. Eluted samples were analyzed using an attached MS apparatus configured to measure m/z up to 500.
RESULTS AND DISCUSSION
Ten species of Iris were confirmed to occur in Jordan; nine of which are growing in the wild and these are: I. atrofusca Baker, I. atropurpurea Baker, I. aucheri (Baker) Sealy, I. edomensis Sealy, I. nigricans Dinsmore, I. petrana Dinsmore, I. postii Mouterde, I. regis-uzziae Feinbrun and I. vartanii Foster. The tenth species, I. germanica L., is a cultivated one (Al-Khader, 1997). I. atrofusca and I. petrana are classified as threatened plant species of Jordan (Al-Eisawi et al., 2000). The genus Iris has been reported to contain irone-type volatile oils (Williams and Harborne, 1985).
In the current effort, several trials were carried out to propagate tissue cultures from different organs of I. atrofusca and I. petrana.
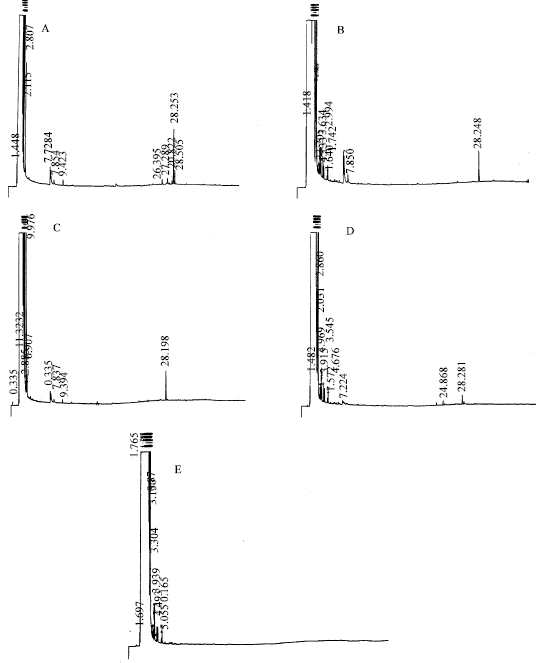 | |
| Fig. 1a: | GC diagrams of representative volatile fractions isolated from I. atrofusca: (A) Untreated control calli; (B) Calli treated with 0.01% w/v quinine sulfate; (C) Calli treated with 0.02% w/v quinine sulfate; (D) Calli treated with 0.03% w/v quinine sulfate and (E) Calli treated with 0.04% w/v quinine sulfate |
However, all attempts were futile except those performed using flower base explants. Incidentally, successful calli propagation was reported from the leaf base of another Iris species, namely, I. nigricans (Shibli and Ajlouni, 2000).
Previous reports indicated the presence of irone-type volatile substances in different Iris species, such as I. florentina L. (Harborne and Williams, 2001). Accordingly, it was anticipated that I. atrofusca and I. petrana will contain the same volatile substance (s). This was established by TLC analysis of intact plant distillates obtained from the flowers. Similarly, the propagated calli obtained from flower base explants from both species have been found to contain the same volatile substance as the parent plant. The current effort represents the first attempt to propagate calli from I. atrofusca and I. petrana and to isolate irone type substance from the calli of these species.
GC analysis of the eluates from both Iris species showed the presence of one major peak (retention time ca. 28 min), few moderate peaks (retention times ca. 7 min) and several minor spikes (Fig. 1a and 1b). The major component of the volatile mixture was identified as an irone-type volatile compound based on MS analysis (Fig. 2) and comparison with published literature (Maurer et al., 1989).
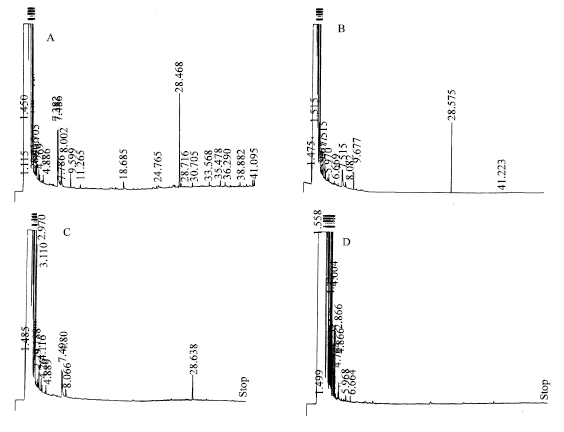 | |
| Fig. 1b: | GC diagrams of representative volatile fractions isolated from I. petrana: (A) Untreated control calli; (B) Calli treated with 0.01% w/v quinine sulfate; (C) Calli treated with 0.02% w/v quinine sulfate and (D) Calli treated with 0.03% w/v quinine sulfate |
The presence of quinine in the culture media affected the height/area of all peaks. However, emphasis was given only to the major peak corresponding to the irone type volatile compound in both species. Figure 1a, b and 3 show a clear dose dependent effect of quinine sulfate on the irone peak. Evidently, increasing the concentration of quinine sulfate in the culture media was associated with a significant reduction in the volatile oil concentration in both species. From these figures, one can clearly see that at 0.03 and 0.04% (w/v) quinine concentrations, the major peak disappeared completely from the distillates isolated from I. petrana and I. atrofusca calli, respectively. Probably, the reason for this conduct is related to toxic effects induced by quinine on certain enzymatic steps in the biosynthesis of this irone-type compound.
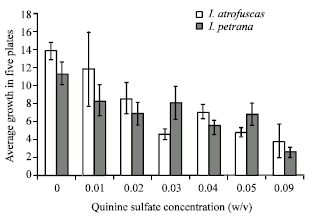
The Fig. 4 indicates that increasing the concentration of quinine sulfate causes steady decrease in callus biomass. However, contrary to the effect of quinine on volatile oil production, biomass growth has not been brought to complete cessation even at 0.09% (w/v) quinine sulfate concentration. Probably, this decrease in calli biomass growth is related to certain toxic effects induced by quinine on vital enzymatic steps. Obviously, the lethal levels of quinine on calli growth are higher than those required to shutdown the production of volatile compounds within both Iris species. Interestingly, this steady decrease of biomass growth was interrupted at two points corresponding to quinine concentrations responsible for the complete cessation of volatile oil production, i.e., 0.03 and 0.04% (w/v) for I. atrofusca and I. petrana, respectively (Fig. 3 and 4).
The association of the transient boost in biomass growth with the cessation of volatile oil production demonstrates the existence of a strong link between different metabolic activities; i.e., biomass and secondary metabolites production. For example, acetyl CoA is involved in the synthesis of volatile oils and vital steroidal substances of the cell membrane (Erman, 1985; Croteau and Gershenzon, 1994). Probably, blocking the volatile oil pathway preserves acetyl CoA for the synthesis of vital steroids, which might explain the excess biomass formation concomitant with the cessation of volatile oil production.
Apparently, other secondary metabolites were also affected by higher quinine sulfate concentrations. At 0.05% w/v the biomass of I. petrana calli showed another transient boost suggesting a similar association with another non-volatile secondary metabolite as seen in Fig. 4.
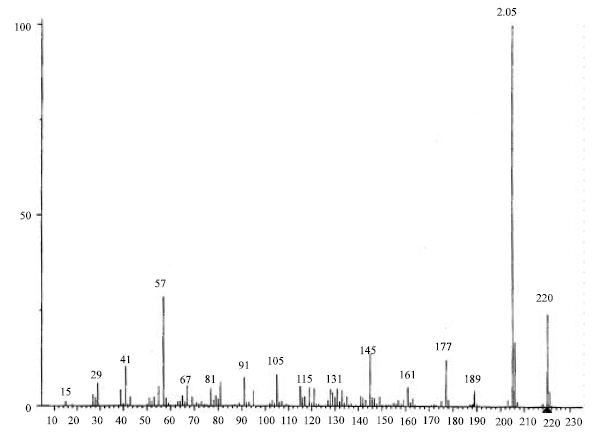 | |
| Fig. 2: | MS fragmentation patterns and isomeric structures of the Iron-type compound identified from calli propagated from I. atrofusca and I. petrana |
 | |
| Fig. 3: | The effect of increasing quinine sulfate concentrations on the production of the irone-type volatile substance in calli from both Iris species |
The observation of this phenomenon only in I. petrana indicates the occurrence of certain non-volatile quinine-sensitive secondary metabolite in this species.
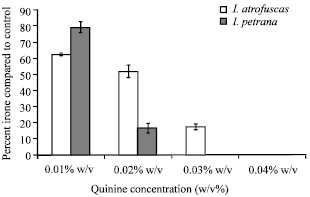 | |
| Fig. 4: | The effect of increasing quinine sulfate concentrations on calli propagation in both Iris species |
Further studies are needed to identify other volatile constituents found in the calli of these Iris species and to investigate the mechanism by which quinine sulfate interferes with volatile oil production and cellular intoxication. It would be very interesting to correlate the LD50’s obtained for pharmaceutical compounds in mice/rats with their observed toxic effects on the volatile oil production in cultured tissues.
ACKNOWLEDGMENTS
The authors wish to thank Mr. Ismail Abatha, Miss Faten Al-Hajj and Mr. Osama Al-Yamani for their technical assistance.
REFERENCES
- Abou-Mandour, A. and G. Binder, 1998. Effect of exogenous growth regulators on plant regenerates from tissue cultures of Mentha spicata and production of volatiles. J. Applied Bot., 72: 233-243.
Direct Link - Aburjai, T., M. Hudaib and V. Cavrini, 2006. Composition of the essential oil from Jordanian germander (Teucrium polium). J. Ess. Oil Res., 18: 97-99.
Direct Link - Al-Eisawi, D.M., 1986. Studies on the flora of Jordan 12. Monocotyledons new to Jordan, with notes on some interesting species. Kew Bull., 41: 349-357.
Direct Link - Berger, R.G., Z. Akkan and F. Drawert, 1990. The essential oil of Coleonema album (Rutaceae) and of a photomixotrophic cell culture derived thereof. Zeitschrift fuer Naturforsch. C, 45: 187-195.
Direct Link - Biemelt, S., H. Tschiersch and U. Sonnewald, 2004. Impact of altered Gibberelin metabolism on biomass accumulation, lignin biosynthesis and photosynthesis in transgenic Tobacco plants. Plant Physiol., 135: 254-265.
Direct Link - Chaturvedi, H.C., S.K. Singh, A.K. Sharma and S. Agnihotri, 2001. Citrus tissue culture employing vegetative explants. Indian J. Exp. Biol., 39: 1080-1095.
Direct Link - Cotton, C.M., J.W. Gramshaw and L.V. Evans, 1991. The effect of a-Naphthalene Acetic Acid (NAA) and benzylaminopurine (BAP) on the accumulation of volatile oil component in cell cultures of tarragon (Artemisia dracunculus). J. Exp. Bot., 42: 377-386.
Direct Link - Giamakis, A., O. Kretsi, I. Chinou and C. G. Spyropoulos, 2001. Eucalyptus camaldulensis: Volatiles from immature flowers and high production of 1,8-cineole and β-pinene by in vitro cultures. Phytochemistry, 58: 351-355.
Direct Link - Harborne, J.B. and C.A. Williams, 2001. The phytochemical richness of the Iridaceaea and its systematic significance. Ann. Bot., 1: 43-50.
Direct Link - Magiatis, P., A. Michaelakis, A.L. Skaltsounis and S.A. Haroutounian, 2001. Volatile secondary metabolite pattern of callus cultures of Chamomilla recutita. Nat. Prod. Lett., 15: 125-130.
Direct Link - Rasol, M.K., H. Cipcic and D. Hagege, 1999. Isoesterases related to cell differentiation in plant tissue culture. Chem. Biol. Intera., 119-120: 587-592.
Direct Link - Sauer, U.G., 2004. Avoidance of animal experiments in the new EU chemicals regulation-opportunities and problems from the point of view of animal welfare. ALTEX: Alternat. Tierexp., 21: 9-14.
Direct Link - Shervington, A., R. Darwish and F. Afifi, 1997. Volatile oil produced by microbial infected Rosmarinus officinalis callus and suspension cultures. Alex. Pharm. Sci., 11: 81-84.
Direct Link - Shibli, R.A. and M.M. Ajlouni, 2000. Somatic embryogenesis in the endemic black Iris. Plant Cell Tissue Organ Cult., 61: 15-21.
Direct Link - Sokmen, M., J. Serkedijeva, D. Daferera, M. Gulluce and M. Polissiou et al., 2004. In vitro antioxidant, antimicrobial and antiviral activities of the essential oil and various extracts from herbal parts and callus cultures of Origanum acutidens. J. Agric. Food Chem., 52: 3309-3312.
Direct Link - Svobada, K.P., R.P. Finch, E. Carion and S.G. Deans, 1995. Production of volatile oils in tissue culture of Origanum vulgare and Tanacetum vulgare. Acta Hortic., 390: 147-152.
Direct Link