
Bishal Gole Tamang
Tribhuvan University, Institute of Agriculture and Animal Science, Rampur, Chitwan, Nepal
Raj Kumar Niroula
Biotechnology Unit, Nepal Agricultural Research Council, Khumaltar, Lalitpur, Nepal
Resham Babu Amgai
Biotechnology Unit, Nepal Agricultural Research Council, Khumaltar, Lalitpur, Nepal
Bindeswar Prasad Sah
Biotechnology Unit, Nepal Agricultural Research Council, Khumaltar, Lalitpur, Nepal
Surya Kant Ghimire
Tribhuvan University, Institute of Agriculture and Animal Science, Rampur, Chitwan, Nepal
Asian Journal of Plant Sciences
Year: 2011 | Volume: 10 | Issue: 7 | Page No.: 347-356







ABSTRACT
Flooding is notorious to rice (Oryza sativa L.) cultivation at any stages threatening the life of >100 millions people in Asia annually. Compare to other cereals rice has unusual capacity to tolerate both short and long term flooding attributed to coordinated efforts of several morphological, anatomical, biochemical and action of adaptive genes. In this study three hundred and thirteen Nepalese rice landrace collected from diverse geographic regions were completely submerged for 14 days. Status of Sub1A gene was monitored in tolerant rice accessions identified here. Diverse rice accession possessed differential elongation growth response and their survival ability ranged from 0-100%. Some of the rice accessions performed better than the tolerant check; FR13A. Monitoring of Sub1A gene presence revealed that some of the tolerant rice accessions are lacking the Sub1A gene and SublA-1 allele. Four accessions; Bhaatsaar, Kariyaparewa pakha, Sauthari and Karangi possessed the better survival response than the FR13A and remaining accessions had poorer response. Among tolerant accessions; Kariyaparewa pakha and Sauthari were lacking the Sub1A-1 allele though followed the quiescence growth response. Interestingly, eight rice accessions with Sub1A-1 allele were found to be intolerant to 14 days of complete submergence. Unlike FR13A all the tolerant and intolerant rice accessions displayed the bleaching of chlorophyll pigment resulting the lower SPAD reading. The detailed morpho-physiological and molecular analysis unveiled that both tolerant and intolerant rice accessions harbored Sub1A gene and adapt the both quiescence and escape strategies in response to flash flooding. Beside Sub1A gene, there might be participation of other stress responsive factors that probably functions in close coordination with Sub1 haplotype. In conclusion, Sub1A-1 alone is not a major contributing factor to confer submergence tolerance in diverse rice accessions, thus haplotype based profiling followed by cloning and sequencing are suggested.
PDF Abstract XML References Citation
Received: August 06, 2011;
Accepted: October 23, 2011;
Published: December 09, 2011
How to cite this article
Bishal Gole Tamang, Raj Kumar Niroula, Resham Babu Amgai, Bindeswar Prasad Sah and Surya Kant Ghimire, 2011. Determination of Adaptive Mechanisms for Flash Flooding Tolerance in Nepalese Cultivated Rice Genepool based on Morpho-physiological and Molecular Analysis. Asian Journal of Plant Sciences, 10: 347-356.
DOI: 10.3923/ajps.2011.347.356
URL: https://scialert.net/abstract/?doi=ajps.2011.347.356
DOI: 10.3923/ajps.2011.347.356
URL: https://scialert.net/abstract/?doi=ajps.2011.347.356
INTRODUCTION
World rice (Oryza sativa L.) production is limited largely by several forms of climate change directed environmental stresses, of which flooding induced submergence is a major constraint worldwide (Xu et al., 2006). Annually about 16% of the world rice areas are unfavorably submerged and affecting the livelihood of more than 100 millions people (Hossain and Abedin, 2004). Rice is unusually adapted to semiaquatic environments because of its well-developed aerenchyma tissues, that facilitate oxygen diffusion through continuous air spaces from shoot to root and avoid O2 deficiency in roots. However, complete submergence due to frequent flooding can adversely affect the plant growth and yield (Fukao et al., 2006; Xuet al., 2006). In some of the rainfed paddy areas, the levels of flood water rise progressively during the rice growing season and can reach up to 7 meters whereas in others, flash flooding can fully submerge the plants for a few days to weeks (Voesenek and Bailey-Serres, 2009). Submergence especially limits the oxygen diffusion rate by 10-4 fold slower than in air resulting anaerobic metabolism and energy crisis (Bailey-Serres and Voesenek, 2008; Licausi and Perata, 2009). As a result, most of the flood-prone areas are planted with five times less yielding landraces that display a remarkable ability to adapt either in deepwater or in flash flooding conditions (Voesenek and Bailey-Serres, 2009). These traditional rice varieties are tolerant to flooding stress due to various adaptive mechanisms to survive periods of hypoxia or anoxia (Mackill et al., 1996; Hattori et al., 2009). An adaptation to submergence involves alterations in molecular, biochemical, physiological, genetical and anatomical/ morphological attributes in rice plants. These include energy generation through fermentative metabolism, aerenchyma development in parenchymal tissues that improves access to O2, activation of ethylene promoted gibberellin (GA) mediated internode elongation in deepwater rice to shootup the foliage above the water surface for gas exchange and restricting growth and conserving precious energy until floodwater recedes in lowland rice (Bailey-Serres and Voesenek, 2008; Hattori et al., 2009). These abilities have enabled the rice crop to cultivate worldwide ranging from rain-fed and irrigated lowlands to deepwater (Khush, 1997).
Recent discovery indicated that both quiescence and escape adaptive mechanisms displayed by different rice accessions are under genetic control (Xu et al., 2006; Hattori et al., 2009; Lee et al., 2009) and explicated in terms of the major Sub1 QTLs and qTIL12 (SK1 and SK2), respectively (Xu et al., 2006; Hattori et al., 2009). These QTLs, were identified in two different ecotypes of rice cultivated in different water regime, encode different subgroup of ERFs (Ethylene Responsive Factors) genes whose expression is activated by plant hormone ethylene. Analysis of near-isogenic and transgenic lines confirmed that Sub1A-1 restricts the escape strategy whereas SNORKELs trigger antithetically through increase GA production and responsiveness and enable the rice plant to survive in deepwater conditions. Submergence induced ethylene activates Sub1A-1, limiting ethylene production by feedback mechanism and promoting GA repressors SLR1 and SLRL1 (Fukao and Bailey-Serres, 2008; Nagai et al., 2010). Recent findings based on comparative microarray and metabolite studies also confirmed that Sub1A-1 regulates numerous transcription factors associated with stress tolerance responses, supporting the past speculation based on physiological study (Jung et al., 2010). Likewise mechanisms of seed germination and underwater seedling growth in the flooded paddy has also recently revealed in which tolerance is governed by CIPK15 gene encoding Calcineurin B-like-interacting protein kinases (Lee et al., 2009). All three adaptive genes express independently of each other and are considered crucial to cope the different types of flooding stress in the rice field. Despite these knowledges on adaptive mechanisms at the gene and protein level, our understanding of the diversity in mechanisms by which different rice accessions survive under varying nature of complete submergence is still to be discovered. Moreover, the potentiality of local Nepalese rice accessions has not been assessed yet against the flash flooding. Therefore, this study was undertaken to dissect the morpho-physiological and molecular mechanism using diverse germplasm representing all Nepalese rice ecosystem. In this research, we tried to elucidate the underlying diverse mechanisms that facilitate the rapid recovery of rice seedlings after flash flooding.
MATERIALS AND METHODS
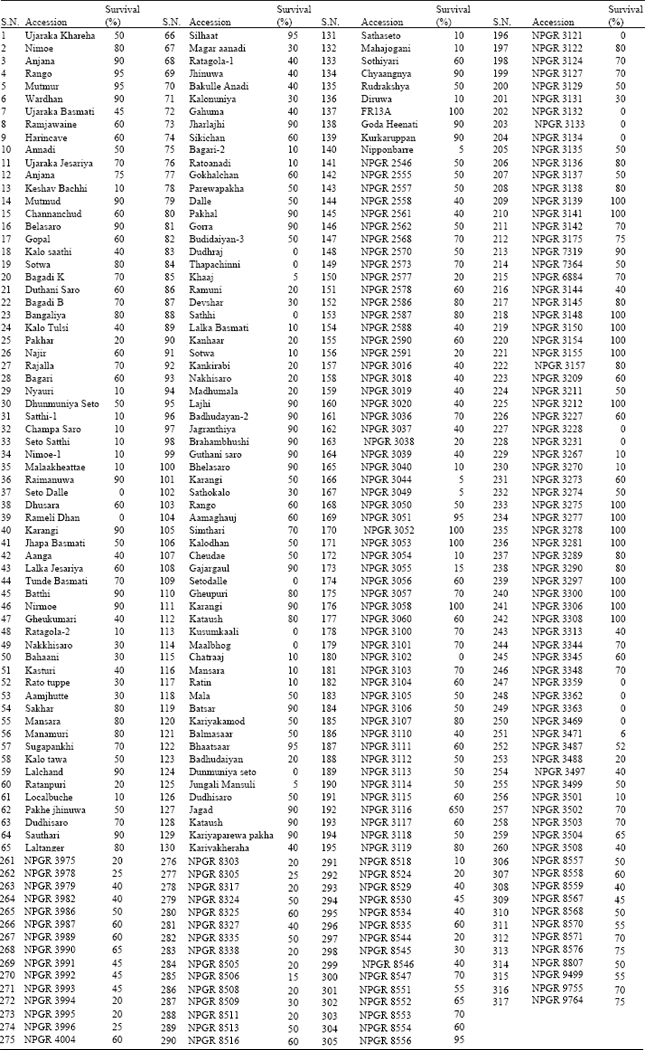
Seeds of 313 rice accessions, collected from different parts of the country was reterived from National Genetic Resource Center, Nepal Agricultural Research Council (NARC), comprising landrace from Terai, Hills and Mountains were subjected to submergence experiment. For the screening of submergence tolerant plants, seeds of each rice accession were surface sterilized and soaked over night in Petri dishes containing sterile water wetted filter paper. Petridishes containing seed were left for 2-3 days in dark with a temperature of 28±2°C for germination. Approximately 120 pre-germinated caryopses were sown in two rows in 5 kg plastic tray (0.38x0.27x0.07 m3) supplied with puddled soil without fertilizers. Each tray consisted of six accessions and performed the experiment in CRD (Completely Randomized Design) with three replications (Fig. 1a). Seedlings were allowed to grow for 14 days at 28±2°C with a 12 h photoperiod. Seedlings when reached the age of 12 days they were thinned to 50 seedlings/row. Fourteen days old seedlings were completely submerged in a 3,300 l water tank (2.44x1.22x1.12 m3) for another 14 days (Fig. 1b). Throughout the experiment the water depth was maintained at 1.05 m. The survival ability was scored after 7 days of desubmergence and survival percent over universal check (FR13A) was calculated as: Survival % over check = (% survival of accessions/% survival of check)x100.
Fourty-four accessions were selected based on their survival percentage (>90 %) and quiscence growth during preliminary glasshouse screening. To determine the molecular mechanism underlying the tolerance in those selected accessions, presence of Sub1A gene was monitored using six sets of gene specific primers as described previously (Xu et al., 2006). To verify the results of preliminary screening and Sub1A monitoring, the accessions with and without Sub1A but having more than 90% survival ability were further subjected to air versus submergence experiment following CRD with three replications, 15 seedlings/replication in 2 kg plastic pots including three rice varieties; FR13A, Goda Heenati and Kurkaruppan as tolerant and Nipponbarre (japonica) as intolerant control. Due to the insufficient amount of seeds, 16 rice accessions were excluded from this experiment. Thus, this confirmatory (pot) experiment only consisted of 28 rice accessions.
 | |
| Fig. 1(a-d): | Initial screening of different rice accessions against flash flooding. (a) 14 d old rice seedlings from different accession prior to submergence are laid out in CRD experiment inside the water tank, (b) morphological response of rice seedlings after submergence, (c) differential survival response of different rice accessions after desubmergence followed by 7 d recovery and (d) differential elongation response showing both quiescence and escape strategy adapted by different rice genotypes |
Height of the seedling and SPAD reading (SPAD 502, Minolta, Japan) were recorded from five randomly choosen seedlings/replication before and just after desubmergence to determine the elongation ability and total chlorophyll content. SPAD reading was recorded from three positions of a leaf. Survival ability of individual accession was scored after 14 days of submergence followed by 7 days of recovery.
Genomic DNA of rice accessions was prepared using modified CTAB method as described by Sul and Korban (1996). PCR reaction was conducted in the 15 μL volume containing 2 μL (100 ng) of genomic DNA, 1.5 μL (1 μM) of each primer, 7.5 μL of 2x GoTaq Green PCR Master Mix (Promega Corporation, Madison, WI, USA) and 2.5 μL PCR H2O. The reaction mixture without template DNA (12.5 μL) was dispensed in each PCR tube. Finally, the DNA template from respective rice accessions was added in PCR reaction. PCR mixture was amplified in a MJ Research PTC-100 ™ Programmable Thermal Controller (MJ Research, Inc, Watertown, MA, USA.) with the following temperature regimes: initial denaturation for 2 min at 95°C followed by 32 cycles of 95°C for 30 sec, annealing at 56-60°C depending on the primers TM for 1 min, extension at 72°C for 2 min and final extension at 72°C for 7 min followed by holding at 4°C as described by Xu et al. (2006). Amplified PCR products were separated in 2% analytical grade agarose gel (Promega Corporation, Madison, WI, USA) using horizontal gel electrophoresis unit in 1xTAE (0.11% Glacial Acetic acid, 0.5 M EDTA and 0.04M Tris base) buffer and run at 100v for 1 h. Gels were stained with 0.1 μg mL-1 ethidium bromide (Promega Corporation, Madison, WI, USA) and then visualized under UV trans illuminator gel documentation system (Wilber Lourmat, Marne-La-Valleen, France.) using 1 μg guide size DNA ladder (Genetix, Biotech Asia Pvt. Ltd). Mean and standard deviation for height and chlorophyll contents and percentage of survival over check were computed using MS EXCEL Software (2007). Monitoring of Sub1A was done based on the presence/absence of the band in the respective lane of the accessions.
RESULTS
Three hundred and thirteen Nepalese rice landrace collected from diverse geographic regions were completely submerged for 14 days.
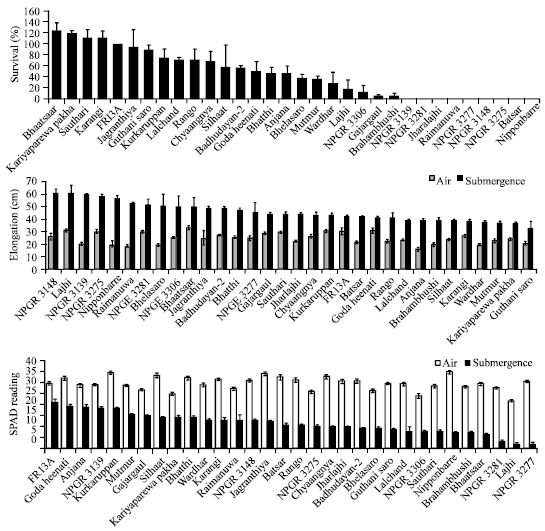 | |
| Fig. 2(a-c): | Morphological and physiological response of different rice accessions after 14 days of complete submergence. (a) Survival response of selected rice accessions (with and without Sub1A gene) after 7 d of recovery. Mean and standard deviation were calculated from three replications with 15 seedlings/replication and is presented in the graph, (b) The elongation growth response of rice seedlings under 14 d of complete submergence. Both quiescence and escape adaptive mechanisms are shown in the graph. Data were collected as described earlier, (c) Graph showing SPAD reading to determine the chlorophyll content before and after submergence |
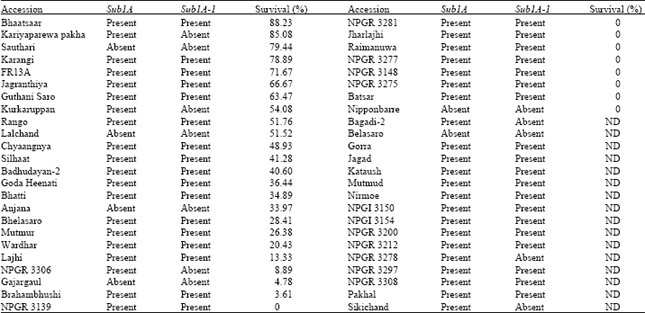
Depending upon the genetic makeup, these accessions possessed differential growth response and survival ability (Fig. 1c-d). In the initial screening, the submergence tolerance response was varied from 0-100% (Table 1) and some of the accessions were better than tolerance check; FR13A (Table 1, Fig. 2a, 3). Based on the result of large scale screening, 44 tolerant rice accessions were selected for further experiment without knowing the molecular mechanisms behind the tolerance. To know the molecular regulation of the tolerance in those selected rice accessions, the presence of major genetic regulator; Sub1A gene was monitored using Sub1A gene and Sub1A-1 allele specific primer and found that some of the tolerance rice accessions lacking both Sub1A gene or SublA-1 allele (Table 2, Fig. 2a). Though 44 rice accessions were selected for Sub1A presence monitoring, unfortunately high stringent submergence experiment using all accessions could not be performed due to lack of adequate number of rice seeds. Therefore, the comparative high throughput screening was restricted to only 28 accessions. In the repeated experiment some of the rice accessions found to be tolerant in earlier experiment showed the intolerancy (Fig. 2a). Four accessions; Bhaatsaar, Kariyaparewa pakha, Sauthari and Karangi possessed the better survival response than the FR13A and rest of the accessions had poorer response. Among tolerant accession; Kariyaparewa pakha and Sauthari were lacking the Sub1A-1 allele and also followed the quiescence growth response (Fig. 3a, b, 2b).
| Table 1: | List of rice accessions used and their survival ability |
 | |
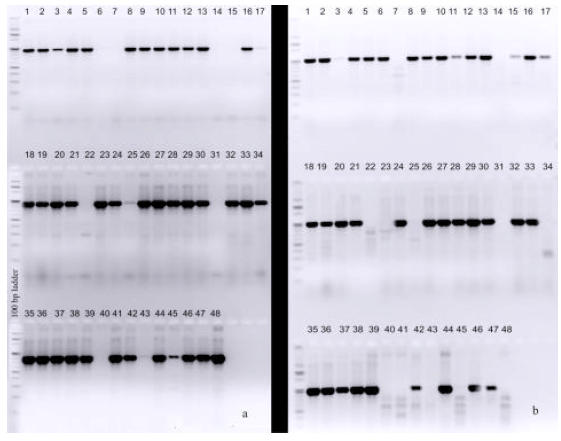 | |
| Fig. 3(a-b): | Monitoring of Sub1A gene in 44 selected rice accessions. Six sets of Sub1A locus specific primer were used as reported earlier (Xu et al., 2006). (a) DNA amplification profiling to detect the presence and absence of Sub1A gene using Sub1-A-7 primer couple, (b) Detection of Sub1A-1 allele in tolerance and intolerance rice accessions using allele specific primer pair; Sub1-A-1. The detail information about the primer used are provided in the Table 3. Number in each lane represents the rice accessions as 1 = NPGR-3275, 2 = Goda Heenati, 3 = NPGR-3306, 4 = NPGR-3308, 5 = NPGR-3154, 6 = NPGR-3150, 7 = Nipponbarre, 8 = FR13A, 9 = NPGR-3148, 10 = NPGR-3277, 11 = NPGR-3200, 12 = NPGR-3297, 13 = NPGR-3139, 14 = NPGR-3278, 15 = NPGR-3212, 16 = NPGR-3281, 17 = Guthani Saro, 18 = Jharlajhi, 19 = Rango, 20 = Badhudayan-2, 21 = Mutmud, 22 = Anjana, 23 = Pakhal, 24 = Silhaat, 25 = Belasaro, 26 = Jagad, 27 = Kataush, 28 = Batsar, 29 = Brahambhushi, 30 = Mutmur, 31 = Lalchand, 32 = Jagranthiya, 33 = Wardhar, 34 = Gorra, 35 = Bhatti, 36 = Lajhi, 37 = Raimanuwa, 38 = Karangi=, 39 = Chyaangnya, 40 = Gajargaul , 41 = Kariyaparewa pakha 42 = Nirmoe, 43 = Sauthari, 44 = Bhaatsaar, 45 = Sikichand, 46 = Bhelasaro, 47 = Kurkaruppan and 48 = Bagadi-2 |
| Table 2: | Selected tolerant rice accession with their mean survival ability and status of Sub1A gene |
 | |
| ND: Not detected due to lack of enough number of rice seed to repeat the experiment | |
| Table 3: | List of primers used for Sub1A monitoring in selected Nepalese rice accessions |
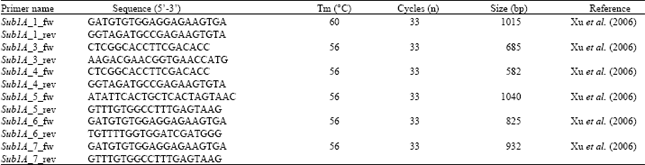 | |
Interestingly, eight rice accessions possessed the presence of Sub1A-1 allele but did not tolerate 14 days of complete submergence (Table 2, Fig. 2a). In this experiment, though accessions; NPGR-3148, -3139 displayed the presence of Sub1A-1allele, they followed the escape adaptive strategy resulting the very poor survival ability under flash flooding (Fig. 2b). The chlorophyll retention after submergence was highest in FR13A than the tolerant accessions identified in this study. Unlike FR13A all the tolerant and intolerant rice accessions displayed the bleaching of chlorophyll pigment resulting the lower SPAD reading (Fig. 2c).
DISCUSSION
Flash flooding tolerance response in rice genotypes that have been evaluated so far harbor Sub1A-1 allele of Sub1A locus and follow an energy saving quiescence growth mechanism (Xu et al., 2006; Bailey-Serres and Voesenek, 2008; Singh et al., 2010). Rice accessions with haplotypes other than Sub1A-1/Sub1C-1 are intolerant to submergence (Singh et al., 2010). Similarly, deepwater response in rice and hypoxia tolerance in Arabidopsis are controlled by SK1 and SK2 and RAP2.12, HRE1 and HRE2 belonging to different members of subgroup VII of the ERF transcription factor family, respectively (Hattori et al., 2009; Licausi et al., 2011). Despite these earlier findings (Fukao et al., 2006; Xu et al., 2006; Fukao et al., 2011), in the present study some of the rice accessions such as Kariyaparewa pakha and Sauthari lacking Sub1A-1 allele displayed the significant level of tolerance. Interestingly, contrasting response was also observed for rice accessions namely NPGR 3139, NPGR 3281, Jharlajhi, Raimanuwa, NPGR 3277, NPGR 3148, NPGR 3275 and Batsar. These rice accessions contained Sub1A-1 allele but the recovery rate was close to nil (Table 2, Fig. 3b).
 | |
| Fig. 4(a-d): | Representative photographs showing 7 d of recovery after 14 d of complete submergence in selected rice accessions |
The poor survival ability in those genotypes could be explained by extra costing of energy for rapid stem elongation response (Fig. 2b) which is to be considered disadvantage for faster regeneration (Perata and Voesenek, 2007; Colmer and Voesenek, 2009). Underwater elongation in rice is triggered by ethylene and GA (Fukao et al., 2006). It is suggested that, in the absence of Sub1A-1, Sub1C facilitates shoot elongation during drowning, through a GA-dependent mechanism (Fukao and Bailey-Serres, 2008). Sub1A-1 reverses the ethylene-dependent increase in GA responsiveness and consequent Sub1C mRNA accumulation. However, based on recombinant genetic studies, Sub1A seems to be the major determinant of submergence tolerance, as Sub1C gene expression does not significantly affect the level of tolerance (Septiningsih et al., 2009).
This study also clearly indicated that the chlorophyll retention capacity after submergence varied with the overall genetic make up rather than the presence of Sub1A-1 allele. FR13A was found to be robust interms of chlorophyll retention capacity and the slow depletion mechanism of chlorophyll could be plus that helps to replenish the energy crisis through instant photosynthesis during recovery (Fig. 2c, 4). Positive correlation between survival and chlorophyll content after submergence was also reported by Das et al. (2009). However, most tolerant rice accessions identified in this study such as Bhaatsaar, Karangi and Sauthari possessed rapid depletion of chlorophyll signifying that chlorophyll retention capacity of the genotypes is independent of submergence tolerance trait. Compare to tolerant check, four genotypes viz. Bhaatsaar, Kariyaparewa pakha, Sauthari and Karangi performed better however, the mechanism of regeneration was quite dissimilar (Fig. 4). In these accessions most of the older leaves died soon after the desubmergence but regeneration of new leaves was abruptly rapid (Fig. 4 and Table 2). Unlike these accessions, upon desubmergence the older leaves of FR13A showed low level of senescent and regained its normal growth very soon.
Based on submergence response and monitoring of Sub1A gene among the 28 rice accessions we hypothesized that in the absence of gene regulating elongation response, the survival ability is independent of Sub1A-1. On the other hand, in the presence of gene responsible for energy consumption process and absence of Sub1 haplotype (Sub1A-1/Sub1C-1), Sub1A-1 allelic form alone is not sufficient to confer the submergence tolerance. To validate these hypotheses Sub1 haplotypic analysis, transcriptomic and hormonal profilings are underway.
CONCLUSION
Following large scale screening of 313 diverse rice accessions we are able to identify four tolerant genotypes. These genotypes were with and without Sub1A-1 allele. From the perspective of rice improvement these germplasm constitute the great assets to broaden the narrow genetic base of submergence tolerance trait. Despite their potential importance, the exact mechanisms of tolerance in those accession are to be unraveled. Therefore, to dissect the underlying hidden mechanisms, further in-depth study using a combined developmental, physiological and omics approaches are suggested.
ACKNOWLEDGMENTS
This research was conducted from the financial support under the Generation Challenge Program (GCP) of Global Biodiversity Trust (Grant No. 10027). Authors are grateful to National Genetic Resource Center (NGRC/NARC Nepal) for providing the seed of the rice accessions included in this study. The laboratory facility of Biotechnology Unit (NARC-Nepal) and its supportive staffs are also gratefully acknowledged for their contribution in DNA extraction and data scoring.
REFERENCES
- Bailey-Serres, J. and L.A. Voesenek, 2008. Flooding stress: Acclimations and genetic diversity. Annu. Rev. Plant Biol., 59: 313-339.
CrossRefPubMedDirect Link - Colmer, T.D. and L.A.C.J. Voesenek, 2009. Flooding tolerance: Suites of plant traits in variable environments. Funct. Plant Biol., 36: 665-681.
CrossRefDirect Link - Das, K.K., D. Panda, R.K. Sarkar, J.N. Reddy and A.M. Ismail, 2009. Submergence tolerance in relation to variable floodwater conditions in rice. Environ. Exp. Bot., 66: 425-434.
CrossRefDirect Link - Fukao, K. and J. Bailey-Serres, 2008. Submergence tolerance conferred by Sub1A is mediated by SLR1 and SLRL1 restriction of gibberellin responses in rice. Proc. Natl. Acad. Sci., 105: 16814-16819.
Direct Link - Fukao, T., E. Yeung and J. Bailey-Serres, 2011. The submergence tolerance regulator SUB1A mediates crosstalk between submergence and drought tolerance in rice. Plant Cell, 23: 412-427.
CrossRef - Fukao, T., K. Xu, P.C. Ronald and J. Bailey-Serres, 2006. A variable cluster of ethylene response factor-like genes regulates metabolic and developmental acclimation responses to submergence in rice. Plant Cell, 18: 2021-2034.
CrossRefDirect Link - Hattori, Y., K. Nagai, S. Furukawa, X.J. Song and R. Kawano et al., 2009. The ethylene response factors SNORKEL1 and SNORKEL2 allow rice to adapt to deep water. Nature, 460: 1026-1030.
CrossRefDirect Link - Jung, K.H., Y.S. Seo, H. Walia, P. Cao and T. Fukao et al., 2010. The submergence tolerance regulator Sub1A mediates stress-responsive expression of AP2/ERF transcription factors. Plant Physiol., 152: 1674-1692.
PubMed - Khush, G.S., 1997. Origin, dispersal, cultivation and variation of rice. Plant Mol. Biol., 35: 25-34.
CrossRef - Lee, K.W., P.W. Chen, C.A. Lu, S. Chen, T.H. Ho and S.M. Yu, 2009. Coordinated responses to oxygen and sugar deficiency allow rice seedlings to tolerate flooding. Sci. Signal., Vol. 2.
CrossRef - Licausi, F. and P. Perata, 2009. Low oxygen signaling and tolerance in plants Adv. Bot. Res., 50: 140-198.
CrossRef - Licausi, F., M. Kosmacz, D.A. Weits, B. Giuntoli, F.M. Giorgi, L.A.C.J. Voesenek, P. Perata and J.T. van Dongen, 2011. Oxygen sensing in plants is mediated by an N-end rule pathway for protein destabilization. Nature.
CrossRef - Mackill, D.J., W.R. Coffman and D.P. Garrity, 1996. Rainfed Lowland Rice Improvement. International Rice Research Intitute, Manila, ISBN-971-22-0071-X,.
Direct Link - Nagai, K., Y. Hattori and M. Ashikari, 2010. Stunt or elongate? Two opposite strategies by which rice adapts to floods. J. Plant Res., 123: 303-309.
CrossRefDirect Link - Perata, P. and L.A. Voesenek, 2007. Submergence tolerance in rice requires Sub1A, an ethylene-response-factor-like gene. Trends Plant Sci., 12: 43-46.
PubMed - Septiningsih, E.M., A.M. Pamplona, D.L. Sanchez, R. Maghirang-Rodriguez and C.N. Neeraja et al., 2009. Development of submergence-tolerant rice cultivars: the Sub1 gene and beyond. Ann. Bot., 103: 151-160.
CrossRef - Singh, N., T.T. M. Dang, G.V. Vergara, D.M. Pandey and D. Sanchez et al., 2010. Molecular marker survey and expression analyses of the rice submergence-tolerance gene SUB1A. Theor. Appl. Genet., 121: 1441-1453.
CrossRef - Sul, I.W. and S.S. Korban, 1996. A highly efficient method for isolating genomic DNA from plant tissues. Plant Tiss. Cult. Biotech. 2: 113-116.
CrossRef - Voesenek, L.A.C.J. and J. Bailey-Serres, 2009. Genetics of high-rise rice. Nature, 4: 959-960.
CrossRef - Xu, K., X. Xu, T. Fukao, P. Canlas and R. Maghirang-Rodriguez et al., 2006. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature, 442: 705-708.
CrossRefDirect Link