
Monther Mohumad Tahat
Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor Darul Ehsan, Malaysia
LiveDNA: 962.1025
ORCID: 0000-0001-7180-1487
Kamaruzaman Sijam
Faculty of Agriculture, Universiti Putra Malaysia, 43400 UPM, Serdang, Selangor Darul Ehsan, Malaysia
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 7 | Page No.: 385-393







ABSTRACT
Ralstonia solanacearum (race 3 biovar 2) is a bacterial wilt causal agent of many plant species. Infects (potatoes Solanum tuberosum, eggplant Solanum melongena, peppers Capsicum annuum, tomatoes Lycopersicon esculentum, geraniums, Geranium carolinianum, ginger Zingiber officinale and a few weed species including bittersweet Celastrus orbiculatus, nightshade Solanum karsense and stinging nettle Urtica dioica. Ralstonia solanacearum can be infectious in the soil for years in the presence of a host. Race 3 biovar 2 is most commonly transmitted by contaminated soil, equipment, water and insect, or by transplantation of infected seeds or seedlings. Management requires use of resistance cultivars, clean and certified seed, good cultural practices, some chemicals fumigation, antagonistic microbes as a biological control like (Mycorrhizal fungi, Streptomyces sp. and Tricoderma sp.) transgenic resistant plant, cropping systems, soil amendments, integrated control, genetically engineered antagonistic and virulent mutants of R. solanacearum.
PDF Abstract XML References Citation
Received: August 16, 2010;
Accepted: September 09, 2010;
Published: August 29, 2012
How to cite this article
Monther Mohumad Tahat and Kamaruzaman Sijam, 2010. Ralstonia solanacearum: The Bacterial Wilt Causal Agent. Asian Journal of Plant Sciences, 9: 385-393.
DOI: 10.3923/ajps.2010.385.393
URL: https://scialert.net/abstract/?doi=ajps.2010.385.393
DOI: 10.3923/ajps.2010.385.393
URL: https://scialert.net/abstract/?doi=ajps.2010.385.393
INTRODUCTION
Ralstonia solanaceraum is an important pathogen of many crops. Formerly called Pseudomonas solanacearum. Historically, strains of R. solanacearum were classified into five races based loosely on host range and into five biovars based on differential ability to produce acid from a panel of carbohydrates (Denny, 2006).
The five races of R. solanacearum have different host ranges and geographic distributions. Race 1 is a poorly-defined group with a very wide host range and is endemic to the southern United States as well as Africa, Asia and South America. Race 2 principally attacks bananas and is found mainly in Central America and Southeast Asia. Race 3 is distributed worldwide and has primarily been associated with potato. Race 4 affects ginger in much of Asia and Hawaii and race 5 affects mulberries in China (Kelman, 1997; Denny, 2006). The origin of R. solanacearum is not clear, but Hayward (1991) suggests it predates the geological separation of the continents as the bacterium has been found in virgin jungle in South America and Indonesia. However, race 3 biovar 2 strains are believed to originate in the Andean highlands and this near-clonal subgroup is widely distributed in tropical ones throughout the world and some temperate regions such as Europe and northern Asia.
Cultural practices, crop rotation and host resistance may provide limited control of R. solanacearum (Kucharek, 1998; Pradhanang et al., 2003). Several plant essential oils and their components showed that some essential oils have significant efficacy against R. solanacearum in vitro and under glasshouse (Momol et al., 1999) and against several soilborne fungi of tomato (Momol et al., 2000). Disease incidences of 15 to 55% have been reported in fresh market tomato in Taiwan, causing losses exceeding 12 million U.S. dollars annually (Hartman et al., 1991). In Hawaii, ginger production was reported to have suffered losses of over 50% during 1998 and 1999 (Yu et al., 2003). Rapid early detection of bacterial wilt is not only in tubers or plant debris but also in soil or soil-related habitats is essential for disease management in the field to prevent losses and further pathogen spread (Janse et al., 1998).
Disease causal agent: Ralstonia solanacearum is a highly heterogeneous bacterial pathogen that causes severe wilting of many important plants (Smith et al., 1995). The disease is also called Southern bacterial blight, R. solanaceous wilt, Southern bacterial wilt and many other common names in countries where it occurs (Buddenhagen and Kelman, 1964). It is an aerobic obligate organism; strains of the pathogen have minimum, optimum and maximum temperature of 10, 35 and 41°C respectively (Kelman, 1953). Ralstonia solanacearum is a gram-negative, non-spore forming rod, about 0.5-0.7 μm x1.5-2.0 μm with a single polar flagellum (Sneath et al., 1986). The bacterium is aerobic and its colonies on solid media are small, irregularly round, white in reflected light and tan in transmitted light (Hayward, 1991). Four races and five biovars were classified based on oxidation of sugar and sugar alcohol (Strider et al., 1981). Different races of R. solanacearum have different host range. For example, race 1 can affect many flowering crops, race 2 can affect banana plant and race 3 can affect potato, tomato and some other Solanaceae, race 4 can affect ginger and some wide plant (Denny and Hayward, 2001). R. solanacearum have variation in metabolic activity into 5 or 6 different biovars. It is a complex species with considerable diversity, although workers have variously divided the species into "group, strains, pathovars, biotype and race (Strider et al., 1981).
Ralstonia solanacearum classification:
Kingdom: Bacteria
Phylum: Proteobacteria
Class: Betaproteobacteria
Order: Burkholderiales
Family: Burkholderiaceae
Genus: Ralstonia
Species: Solanacearum
Other names for R. solanacearum
Bacillus solanacearum Smith 1896
Bacillus solanacearum
Burkholderia solanacearum
Burkholderia solanacearum (Smith, 1896) Yabuuchi and Arakawa (1993)
Pseudomonas solanacearum (Smith, 1896) Smith (1914)
Ralstonia solanacearum (Smith, 1896) Yabuuchi et al. (1995) (http://www.ncipmc.org/ralstonia)
Recently amore phylogenetically meaningful system has classified R. solanacearum into four major genetic groups called phylotypes that reflect the geographical origin and ancestral relationships between strains (Fegan and Prior, 2005). Ralstonia solanacearum race 3 biovar 2 is a soil-borne pathogen that persists in wet soils, depth soil layers (>75 cm) and reservoir plants. The tetrazolium medium (TZC), described by Kelman and Person (1954) is the best for culturing R. solanacearum. The organism produces two easily distinguishable type of colonies; one is small, flat, red and butyrous (Chen and Echandi, 1982) (Fig. 1a) while the other colony is large, elevated, mostly white with light pink centers and full of fluid using Casamino Acid Peptone Glucose (CPG) (Fig. 1b) (Cuppels et al., 1978).
The most important race of R. solanacearum is race 3 biovar 2 (Table 1), which has valuable agricultural hosts.
| Table 1: | The most important races, host range, geographical distribution and biovars of Ralstonia solanacearum |
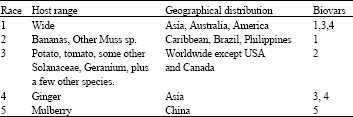 | |
| Adapted from (Denny and Hayward, 2001) | |
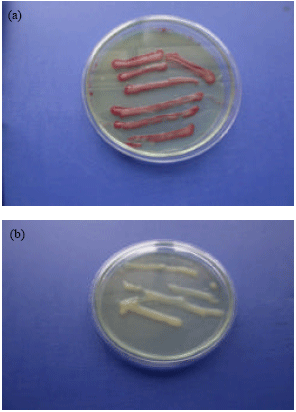 | |
| Fig. 1: | (a) Casamino Acid Peptone Glucose (CPG), (b) Tetrazolium Media (TZC) |
Occurrence and host range: Bacterial wilt caused by R. solanacearum has been described on a wide range of hosts in many tropical and subtropical regions (Agrios, 2005). In the absence of susceptible crops, alternative weed hosts and non-host plants play important roles for the survival of R. solanacearum strains (Granada and Sequeria, 1983). Ralstonia solanacearum causes lethal wilting disease in more than 200 plant species (Denny, 2000), while more than 450 plant species were listed as host plants for R. solanacearum including many important and economic crops (Hayward, 1991). Host range of R. solanacearum includes several hundred species representing 44 families of plants and many newly recognized hosts. R. solanacearum biovar 3 has been described on some woody perennial hosts including cashew (Anacardium occideutale) and custard apple (Annona spp.) (Mayers and Hutton, 1987).
The reason for these highly different and heterogeneous bacterial pathogen is not obvious; nevertheless it is assumed that specific pathogenic strains for certain hosts may have evolved only in certain parts of the world and are not found elsewhere or these hosts may only be susceptible where a number of environmental factors such as temperature, rainfall, soil type, inoculums and other soil biological factors are conductive to disease expression coincide (Hayward, 1991). There is an extensive literature on the disease, particularly in terms of host range, geographic distribution, occurrence of various biovars, strains and the ability to survive in soil and in association with plants debris and weed hosts (Persley, 1992).
Disease distribution: The first record of bacterial wilt Ralstonia solanacearum (Smith, 1896) Pseudomonas solanacearum E. F. Smith) in the world was reported by Burrill in 1890 in Japan. That was found to be on tuber rot of potato (Gota, 1992). Across the world there are differences between R. solanacearum races and biovars depending on the geographical distribution (Hayward, 1991; Buddenhagen and Kelman, 1964). Biovar 1 is predominant in USA and biovar 3 in Asia, whereas biovar 2 and 5 occur in Australia (Pitkethley, 1981) and China (He et al., 1983). It is also found that biovar 4 occurs in India and Indonesia. In Africa the bacterial wilt disease was recorded in Egypt, Libya, South Africa, Zambia and Burundi (OEPP/EPPO, 1999). In the Philippines all of biovars 1-4 have been found and here as elsewhere in Asia biovar 3 is a predominant biovar in lowland regions. Strawberry is a host in Japan and Taiwan but not in the southeastern USA (Hayward, 1991). Around 43 plant species were found as a plant host of the bacterial wilt disease in Malaysia especially biovar 3 race 2 (Abdullah, 1982). Bacterial wilt disease was also observed in Cameron Highland/Malaysia at about 1545 M above the sea level. In addition, the disease has been observed and the pathogen was isolated from the infected crop plants and weeds at the farm of Universiti Putra Malaysia, Selangor (Abdullah, 1988, 1992).
Ralstonia solanacearum is listed as a quarantine organism in the European Union (EU) (Anonymous, 1995). In EPPO region the bacterial wilt disease was found in Belgium, Spain, Netherland, Germany, United Kingdom and Hungary (OEPP/EPPO, 1999). Bacterial wilt causal agent can spread between countries by water, latently infected planting materials and soil. One plant species that is seriously affected by bacterial wilt is tomato and the efforts to grow tomato widely in the tropics have generally been hampered by this disease (Hayward, 2000). Two new hosts for Ralstonia solanacearum: davana (Artemisia pallens) and coleus (Coleus forskohlii) were recorded by Chandrashekara and Prasannakumar (2010), both are important crops in medicinal and aromatic industries in India. Coleus and davana plants showing typical wilt symptoms were found.
Disease symptoms: Ralstonia solanacearum is characterized by sudden wilting of foliage and the young plant is affected more. The symptoms occur as discoloration of the vascular system from pale yellow to dark (Gota, 1992). The plant infected with R. solanacearum may express all or none of these symptoms, even under typical environmental conditions that are ideal for the pathogen and typically this is a commonly observed condition known as latency. The pathogen enters roots through wounds caused by transplanting, cultivation, nematode, insects and through natural wounds. Then it starts to multiply rapidly in the vascular system, finally the xylem elements are filled with bacterial cell and slime (Kelman and Sequeira, 1965). Molecular aptamers are single-stranded oligonucleotides that can specifically bind with high affinity to a variety of molecules ranging from macromolecules to small compounds. Aptamers potentially could be produced and used in the future as reproducible, fast and highly-specific diagnostic tools for R3bv2. (Champoiseau et al., 2009a).
Successful and efficient colonization requires production of molecular mass Extracellular Polysaccharide (EPS) in a high amount and multiple extracellular protein (EXPs) (Denny, 2000). The incidence the disease infection may range from a few scattered plants or loci of infection in fields where low or erratic natural infestations occur to the rapid death of the plants (Kelman and Sequeira, 1965). The bacterium rapidly spreads upward in the vascular system from secondary roots to larger roots and then to the stem. After that, the plant starts to suffer from wilting irreversibly. Older plant leaves first show wilting before the youngest leaves or one sided wilting and stunting and finally the plant wilts permanently and dies (Agrios, 2005).
Massive invasion of the cortex might have resulted in the appearance of water-soaked lesions on the external surface of the stem; if an infected stem is cut crosswise tiny drops of dirty white or yellowish viscous ooze exude from several vascular bundles (Champoiseau et al., 2009b). The pathogen ingresses a plant through the roots, penetrate the xylem, systemically colonize the stem and causes wilt symptoms (Kelman, 1953). This bacterium causes wilt by infecting plants through roots and colonizing stem vascular tissue. Although diseased plants can be found scattered in the field. Under natural conditions, the initial symptom in mature plants is wilting of upper leaves during hot days followed by recovery throughout the evening and early hours of the morning. The wilted leaves maintain their green color as disease progresses. Under hot humid conditions disease complete wilting occurs and the plant will die. The brown discoloration shown in the lower stem vascular tissues (Kucharek, 1998).
CONTROL METHODS
It is difficult to control bacterial wilt disease in the soil (Jones, 1997). Various control strategies were developed to control and suppress this disease including host-plant resistance and biological control (Dalal et al., 1999). Potential biological agents were used to control bacterial wilt of tomato (Lycopersicon esculentum) include vesicular-arbuscular mycorrhizae (VAM) (Halos and Zorrilla, 1979) and some naturally occurring antagonistic rhizobacteria such as Bacillus sp. (Silveira et al., 1995), Pseudomonas sp. (Guo et al., 2001).
The most important and applicable methods are:
Resistance cultivars: The best strategy to control bacterial wilt caused by R. solanacearum is breeding for resistance cultivar (Persley, 1992). A virulent mutant of R. solanacearum has been used as a bio-control agent for the virulent pathogen (Trigalet and Trigalet-Demery, 1990). Abdalla and Abdulla (1998) found that the degree of susceptibility to bacterial wilt is significantly different among six tomato cultivars which were tested and this indicated that the additive genes were more important than the non-additive genes. Thus, in breeding programs, selection for disease resistant plants (genotype) after each generation is recommended. In the United States, Southern tomato transplant growers sometimes prevent the disease by avoiding infected fields (Hayward, 1964). The infection by R. solanacearum can be controlled by the use of hot pepper accessions (Capsicum annuum L.), sweet pepper (Capsicum annuum L.) in Japan. Bacterial multiplication in stems of resistant tomato plants was suppressed owing to the limitation of pathogen movement from the protoxylem or the primary xylem to other xylem tissues. The limitation was most conspicuous in Hawaii 7996. Grafting experiments indicated that the percentage of wilting of Ponderosa scions was less on Hawaii 7996 rootstocks than that on the most resistant rootstock (LS-89) used in Japan. Hawaii 7996 could be an alternative genetic source for breeding for resistance to bacterial wilt (Nakaho et al., 2004).
Cultural practices: Crop rotation, intercropping or incorporation, green manure and planting a susceptible crop such as mungbean before the cultivation have been practiced (Hartman et al., 1993). Crop rotation with a non-susceptible crop provides some control, but this measure is difficult to use because of the wide host range of the pathogen (Kelman, 1953). Islam and Toyota (2004) demonstrated that the bacterial wilt of tomato was suppressed in the poultry and farmyard manure (FYM) added soils and higher microbial activity was likely responsible. In Nepal, the importance of crop rotation and resistance cultivars were studied by Adhikari and Basyat (1998). They reported that the appearance of bacterial wilt symptoms were delayed by 1-3 weeks and the wilt severity was reduced by 20-26% when a susceptible tomato was grown after corn (Zea mays), lady’s finger (Abelomoschus esculentum), cowpea (Vigna unguiculata), or resistant tomato. Finally it is found that crop rotation with corn, lady’s finger, cowpea and resistance cultivars appeared to be useful management strategies to control tomato bacterial wilt. The infection of potato plant by Ralstonia solanacearum can be significantly reduced by using non-susceptible crops and crop rotation for 5-7 years (Smith et al., 1995).
Rice husks, corn stalke, cow manure, oyster shell powder and mineral ash have been used by improved mixture called Sun and Hung (SH) (Sun and Hung, 1985). The application of the organic amendment (Wydra et al., 2005)and compost released biologically active substances from crop residues and soil microorganisms such as allelochemicals (Bailey and Lazarovits, 2003). The Sun hemp mixture reduced the bacterial population after 4 weeks of incubation at 2, 6 and 10% rates of incorporation of inoculated plants wilted and died after the second week (Hartman et al., 1993). The addition of household compost resulted in enhanced decline rate of Ralstonia solanacearum population (Schonfeld et al., 2003). Corn stalk, rice straw and tree bark were used for the suppression of tomato bacterial wilt caused by R. solanacearum in Malaysia and the results showed that all compost used significantly reduced the disease severity index but the disease incidence was the lowest in bark compost (Masyitah, 2004). Abdullah et al. (1983) found that soil type and moisture levels individually as well as in combination had a significant effect on the severity of the bacterial wilt of groundnut. Studying the distribution of disease in Malaysia related to the soil types illustrated that the tobacco plants were infected more when grown on organic soils and light, heavy and intermediate types of mineral soils. Also disease incidence in organic soils (more than 65% organic matter) is normally low (Abdullah, 1988). Cow manure fertilization suppressed brown rot caused by R. solanacearum in most soils with a clear shift in rhizospere bacterial community. Stenotrophomonas maltophilia, isolated from the rhizosphere of eggplant in the Egyptian Delta, was antagonistic to R. solanacearum in vitro; its antagonistic activity was not Fe-siderophore dependent. A selected S. maltophilia strain survived longer and reduced R. solanacearum survival more in Egyptian than in Dutch clay soils and suppressed potato brown rot in Egyptian soil (by at least 36%) but not in Dutch soil (Messiha, 2006). The populations of native Ralstonia spp. were reduced from 2.4-7x108 colony forming units (CFU) g-1 to 0-115 CFU g-1. Heat treatment reduced bacterial wilt incidence by 50-75% (Kongkiattikajorn et al., 2006).
Chemical methods: Disease control using chemicals has been difficult because of the localization of the pathogen inside the xylem and its ability for survival at depth in the soil. In addition, this kind of control is not economically feasible in the field. Some scientists reported that there are no bactericides available for chemical control of the bacterial wilt disease (Hartman et al., 1993), while others, reported that it is difficult to control bacterial with chemicals (Grimault et al., 1992). The bactericides Terlai have been tested in Taiwan under both green house and field conditions (Hartman et al., 1993) and it was found that chemical control through soil fumigation and antibiotics (Penicillin, Ampicillin, Tetracycline and Streptomycin) has shown little suppression of Ralstonia solanacearum (Murakoshi and Takahashi, 1984). Some fumigation bactericides increased the extent of wilt caused by Ralostonia solanacearum such as chloroform (Shiomi et al., 1999). Antagonistic, avirulent bacteriocin-producing Ralstonia solanacearum strains were investigated for potential biological control of tomato (Lycopersicon esculentum) bacterial wilt in Brazil, it was demonstrated that none of the bacteriocin-producing strains was inhibited by its own bacteriocin (Araujo et al., 2004).
During 1992-1995 crop season, the field trials on the management of bacterial wilt Ralostonia solanacearum of tomato revealed that Streptomycin proved to be most effective as it gave highest disease control (79.5%) and lead to maximum yield production (274.60 ha-1) compared to asafetida (Ferula foetida) (70.30%) and turmeric powder (Curcuma longa Linn.) (69%) (Mazumder, 1998). Many plant species produce volatile essential oil compounds. These oils are considered to play a role in host defense mechanisms against plant pathogens (Mihaliak et al., 1991). Essential oils and their components, usually from medicinal plants, have been recognized as having fungicidal effects (Wilson et al., 1997), but their efficacy as a biofumigant on R. solanacearum has not been studied prior to 1999.
Different sugars and amino acids were added to a conducive soil to study their effects on bacterial wilt of tomato caused by Ralstonia solanacearum YU1Rif43. Most of the seeds failed to germinate in soils to which serine, glycine and alanine were applied at the same rate. At 2.5 mg g-1, the inhibitory effect on tomato germination disappeared except for methionine. The compounds that showed the most suppressive effect were glucose, proline, glutamine, serine, arginine and lysine. The pathogen utilized glucose, proline and glutamine, but not serine, arginine and lysine (Posas et al., 2007).
Biological control: Biological control can be defined as the direct and accurate management of common components of ecosystem to protect plants against pathogens. It is acceptable as a key practice in sustainable agriculture (Azcon-Augiler and Barea, 1996). Biological control preserves environmental quality by reducing the dependency on chemical input and maintaining sustainable management practices (Barea and Jeffries, 1995). Wall and Sanchez (1992) reported that when bacteriophages, which are capable of attacking the bacterial wilt pathogen, used as a bio-control agent showed that bacteriophages play an important role in suppressing the population dynamics of R. solanacearum. Plant growth promoting bacteria (PGPR) strains were reported to be a promising bio-control agent to control R. solanacearum. It was found that they were able to reduce the disease in different levels and increased the yield of tomato plant (Guo et al., 2004; Kenichi, 2004).
118 strains of rhizobacteria, were screened against an Ethiopian R. solanacearum strain. On the basis of in vitro screening, six strains (RP87, B2G, APF1, APF2, APF3 and APF4) shown good inhibitory effect were selected for in planta testing in a greenhouse. The study showed that APF1 and B2G strains reduced significantly disease incidence and increased weight of tomato plants. Area under Disease Progress Curves (AUDPC) was reduced by 60 and 56% in plants inoculated with APF1 and B2G strains, respectively. APF1 was found to be the most beneficial strain in disease suppression and also growth promotion resulting in 63% dry weight increase compared to untreated control (Lemessa and Zeller, 2007).
The effect of Biological Soil Disinfestation (BSD) was tested in glass vessels, microplots and in an accidentally infested commercial field. BSD is based on production of toxic organic acids through anaerobic digestion of fresh organic matter. BSD was accomplished by incorporating grass or potato haulms in soil and covering the soil with airtight plastic survival of R. solanacearum in soil and potato tubers was significantly reduced in the BSD treatment (>93%) and not in separate grass amendment -or plastic cover treatments (Messiha, 2006). There have been a few studies of the potential role of AMF for the protection of plants from bacterial wilt (Hayward, 1991). Glomus mosseae was able to suppress the infection of tomato plant by R. solanacearum in the glasshouse conditions (Tahat et al., 2010). Zhu and Yao (2004), found that the Localized and systemic increase of phenol in tomato roots induced by Glomus versiforme inhibits Ralstonia solanacearum.
CONCLUSION
Bacterial wilt, caused by Ralstonia solanacearum, is responsible for severe losses to many important crops, mainly Solanaceous plants and bananas, This microorganism is the causal agent of bacterial wilt, moko disease in banana southern wilt of geranium and potato brown rot. Develop disease management to control bacterial wilt on tomato, potato and geraniums including: 1-Screen additional chemical and biological control products. 2-Exclude the pathogen from tomato transplants, potato seeds and geranium cuttings and develop vegetative plant material certification schemes. 3-Study the effects of crop rotation, cover crops and mulches on pathogen dynamics and disease incidence 4- Develop additional DNA and immunological detection tools that can be used reliably to distinguish subgroups of R. solanacearum, especially Race 3 biovare 2, from other endemic strains of Ralstonia. 5-Educate county extension, growers and crop advisors in sampling, monitoring and management of related diseases.
ACKNOWLEDGMENT
The authors would like to thank the University Putra Malaysia for supporting this work.
REFERENCES
- Abdullah, H., 1992. Bacterial wilt in Malaysia: Hosts, disease incidence and geographical distribution. ACIAR. Proc., 45: 334-337.
Direct Link - Abdullah, H., L.M. Maene and H. Naib, 1983. The effects of soil types and moisture levels on bacterial wilt disease of groundnut (Arachi hypogaea). Pertanika, 6: 26-31.
Direct Link - Adhikari, T.B. and R.C. Basnyat, 1998. Effect of crop rotation and cultivar resistance on bacterial wilt of tomato in Nepal. Can. J. Plant Pathol., 20: 283-287.
CrossRefDirect Link - Agrios, G.N., 2005. Plant Pathology. 5th Edn., Elsevier, Cambridge, Massachusetts, ISBN-13: 9780080473789, Pages: 922.
Direct Link - Azcon-Aguilar, C. and J.M. Barea, 1997. Arbuscular mycorrhizas and biological control of soil-borne plant pathogens-an overview of the mechanisms involved. Mycorrhiza, 6: 457-464.
CrossRefDirect Link - Bailey, K.L. and G. Lazarovits, 2003. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res., 72: 169-180.
Direct Link - Buddenhagen, I. and A. Kelman, 1964. Biological and physiological aspects of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol., 2: 203-230.
CrossRefDirect Link - Champoiseau, P., J.B. Jones, C. Allen and T. Momol, 2009. Description and strategies for best management of Ralstonia solanacearum Race 3 biovar 2 as a potential incitant of bacterial wilt of tomato. Proceedings of the 24th Annual Tomato Disease Workshop Held on, Nov. 3-5, State College, Pennsylvania, pp: 1-35.
Direct Link - Cuppels, D.A., R.S. Hanson and A. Kelman, 1978. Isolation and characterization of a bacteriocin produced by Pseudomonas solanacearum. J. Gen. Microbiol., 109: 295-303.
Direct Link - Denny, T.P., 2000. Ralstonia solanacearum. A plant pathology in touch with its host. Trinds Microbiol., 8: 486-489.
PubMed - Araujo, J.S., P. de Rodrigues, R. Ribeiro, L.D. de, K.S. Goncalves and J.C. Polidoro, 2004. Bacteriocin production by Brazilian isolates of Ralstonia solanacearum in vitro. ISHS. Acta Horticulturae International Symposium on Tomato Diseases. Brazil. http://www.actahort.org/members/showpdf?booknrarnr=695_37.
- Fegan, M. and P. Prior, 2005. How Complex is the Ralstonia solanacearum Species Complex? In: Bacterial Wilt Disease and the Ralstonia solanacearum Species Complex, Allen, C., P. Prior and A.C. Hayward (Eds.). American Phytopathological Society Press, St. Paul, MN., USA., ISBN: 0890543291, pp: 449-461.
- Granada, G.A. and L. Sequeira, 1983. A new selective medium for Pseudomonas solanacearum. Plant Dis., 67: 1084-1088.
Direct Link - Grimault, V. Schmit, J. and P. Prior, 1992. Some characteristics involved in bacterial wilt Pseudomonas solanacearum resistance in tomato. ACIAR Proc., 45: 112-119.
Direct Link - Guo, J.H., H.Y. Qi, Y.H. Guo, H.L. Ge, L.Y Gong, L.X. Zhang and P.H. Sun, 2004. Biocontrol of tomato wilt by plant growth-promoting rhizobacteria. Biol. Control, 29: 66-72.
CrossRefDirect Link - Hartman, G.L., W.F. Hong and T.C. Wang, 1991. Survey of bacterial wilt on fresh market hybrid tomatoes in Taiwan. Plant Prot. Bull., 33: 197-203.
Direct Link - Hayward, A.C., 1964. Characteristics of Pseudomonas solanacearum. J. Applied Bacteriol., 27: 265-277.
CrossRefDirect Link - Hayward, A., 2000. Ralstonia solanacearum. In: Encyclopedia of Microbiology, Lederberg, J. (Ed.) 2nd Edn., Academic Press, ISBN-10: 0122268008, pp: 32-42.
Direct Link - Hayward, A.C., 1991. Biology and epidemiology of bacterial wilt caused by Pseudomonas solanacearum. Annu. Rev. Phytopathol., 29: 65-87.
CrossRefDirect Link - He, L.Y., L. Sequeira and A. Kelman, 1983. Characteristics of strains of Pseudomonas solanacearum from China. Plant Dis., 67: 1357-1361.
CrossRefDirect Link - Islam, T.M. and K. Toyota, 2004. Suppression of bacterial wilt of tomato by Ralstonia solanacearum by incorporation of composts in soil and possible mechanisms. Microbes Environ., 19: 53-60.
Direct Link - Lemessa, F. and W. Zeller, 2007. Screening rhizobacteria for biological control of Ralstonia solanacearum in Ethiopia. Biol. Control, 42: 336-344.
CrossRefDirect Link - Mayers, P.E. and D.G. Hutton, 1987. Bacterial wilt, a new disease of custard apple: symptoms and Etiology. Ann. Applied Biol., 111: 135-141.
CrossRef - Mihaliak, C.A., J. Gershenzo and R. Croteau, 1991. Lack of rapidmonoterpene turnover in rooted plants, implications for theories of plant chemical defense. Oecologia, 87: 373-376.
CrossRef - Nakaho, K., H. Inoue, T. Takayama and H. Miyagawa, 2004. Distribution and multiplication of Ralstonia solanacearum in tomato plants with resistance derived from different origins. J. Gen. Plant Pathol., 70: 115-119.
CrossRefDirect Link - Posas, B.M., K. Toyota and T.M. Islam, 2007. Inhibition of bacterial wilt of tomato caused by Ralstonia solanacearum by sugars and amino acids. Microb. Environ., 22: 290-296.
CrossRefDirect Link - Pradhanang, P.M., M.T. Momol, S.M. Olson and J.B. Jones, 2003. Effects of plant essential oils on Ralstonia solanacearum population density and bacterial wilt incidence in tomato. Plant Dis., 87: 423-427.
Direct Link - Schonfeld, J., A. Gelsomino, L.S. van Overbeek, A. Gorissen, K. Smalla and J.D. van Elsas, 2003. Effects of compost addition and simulated solarization on the fate of Ralstonia solanacearum biovar 2 and indigenous bacteria in soil. FMES Microbiol. Ecol., 43: 63-74.
CrossRefDirect Link - Shiomi, Y., M. Nishiyama, T. Onizuka and T. Marumoto, 1999. Comparison of bacterial community structures in the rhizoplane of tomato plants grown in soils suppressive and conducive towards bacterial wilt. Applied Environ. Biol., 65: 3996-4001.
Direct Link - Da Silveira, E.B., R.L.R. de Mariano, S.J. Michereff and M. Menezes, 1995. Antagonism of Bacillus spp. against Pseudomonas solanacearum and effect on tomato seedling growth. Fitopatol. Bras., 20: 605-612.
Direct Link - Smith, J.J., L.C. Offord, M. Holderness and G.S. Saddler, 1995. Genetic diversity of Burkholderia solanacearum (Synonym Pseudomonas solanacearum) race3 in Kenya. Applied Environ. Microbiol., 61: 4263-4268.
Direct Link - Strider, D.L., R.K. Jones and R.A. Haygood, 1981. Southern bacterial wilt of geranium caused by Pseudomonas solanacearum. Plant Dis., 65: 52-53.
Direct Link - Tahat, M.M., K. Sijam and R. Othman, 2010. The role of tomato and corn root exudates on Glomus mosseae spores germination and Ralstonia solanacearum growth in vitro. Int. J. Plant Pathol., 1: 1-12.
CrossRefDirect Link - Trigalet, A. and D. Trigalet-Demery, 1990. Use of a virulent mutants of Pseudomonas solanacearum for the biological control of bacterial wilt of tomato plants. Physiol. Mol. Plant Pathol., 36: 27-38.
CrossRef - Wilson, C.L., J.M. Solar, A. El Ghaouth and M.E. Wisniewski, 1997. Rapid evaluation of plant extracts and essential oils for antifungal activity against Botrytis cinerea. Plant Dis., 81: 204-210.
CrossRefDirect Link - Wydra, K., R. Diogo, E. Dannon and J. Semaru, 2005. Soil Amendment with silicon and bacterial antagonists induce resistance against bacterial wilt caued by Raltoina solanacearum in tomato. Proceedings of the International Conference on Agricultur Resaerch for Development Tropentang, October 11-13, 2005, Stuttgart-Hohenheim, Germany, pp: 1-5.
Direct Link - Yabuuchi, E., Y. Kosako, I. Yano, H. Hotta and Y. Nishiuchi, 1995. Transfer of two Burkholderia and an Alcaligenes species to Ralstonia Gen. Nov.: Proposal of Ralstonia pickettii (Ralston, Palleroni and Doudoroff 1973) Comb. Nov., Ralstonia solanacearum (Smith 1896) Comb. Nov. and Ralstonia eutropha (Davis 1969) Comb. Nov. Microbiol. Immunol., 39: 897-904.
CrossRefDirect Link - Yu, Q., A.M. Alvarez, P.H. Moore, F. Zee and M.S. Kim et al., 2003. Molecular diversity of Ralstonia solanacearum isolated from ginger in Hawaii. Phytopathology, 93: 1124-1130.
Direct Link - Zhu, H.H. and Q. Yao, 2004. Localized and systemic increase of phenols in tomato roots induced by Glomus versiforme inhibits Ralstonia solanacearum. J. Phytopathol., 152: 537-542.
CrossRefDirect Link