
A. Promkhambut
Department of Plant Science and Agricultural Resources, Faculty of Agriculture,Khon Kaen University 40002, Thailand
A. Younger
School of Agriculture, Food and Rural Development, Agriculture Building, Newcastle University,Newcastle Upon Tyne, NE1 7RU, United Kingdom
A. Polthanee
Department of Plant Science and Agricultural Resources, Faculty of Agriculture,Khon Kaen University 40002, Thailand
C. Akkasaeng
Department of Plant Science and Agricultural Resources, Faculty of Agriculture,Khon Kaen University 40002, Thailand
Asian Journal of Plant Sciences
Year: 2010 | Volume: 9 | Issue: 4 | Page No.: 183-193







ABSTRACT
The aim of this study was to investigate the effect of waterlogging on morphological and physiological traits of sorghum (Sorghum bicolor L. Moench) cultivars. Four sorghum cultivars, cv. Wray, Keller, Bailey (sweet cultivar) and cv. SP1 (forage cultivar) at five expanded leaf stage were subjected to 20 days of waterlogging and drained pots were kept as the control. Twenty days of waterlogging did not cause a significant difference in shoot and root biomass among cultivars. Flooding reduced leaf area (69%), plant height (30%) and youngest leaf expansion rate of all cultivars but severely reduced in SP1 (35-80%). Flooding promoted leaf senescence of all cultivars and biomass allocation to shoot (increase in shoot/root) in Wray, Keller and Bailey, but increased biomass partitioning to root in SP1. The initiation of new nodal root was noted in SP1, whereas the ability to maintain root surface area by increase in longest root length and nodal root development near soil surface was found in Wray. Photosynthetic rate, stomatal conductance and transpiration rate were severely reduced under waterlogging conditions of sweet cultivars (65-78%), but enhanced over the control in forage cultivar (56%). The ability to conserve root surface area, allocate more biomass to shoot during waterlogging and develop root near soil surface may support new growth in Wray, whereas the ability to maintain leaf gas exchange parameters in SP1 was due to the active nodal root growth. Nevertheless, there was no relationship between photosynthetic rate and shoot growth of sorghum under anaerobic conditions.
PDF Abstract XML References Citation
Received: March 12, 2010;
Accepted: May 28, 2010;
Published: June 19, 2010
How to cite this article
A. Promkhambut, A. Younger, A. Polthanee and C. Akkasaeng, 2010. Morphological and Physiological Responses of Sorghum
(Sorghum bicolor L. Moench) to Waterlogging. Asian Journal of Plant Sciences, 9: 183-193.
DOI: 10.3923/ajps.2010.183.193
URL: https://scialert.net/abstract/?doi=ajps.2010.183.193
DOI: 10.3923/ajps.2010.183.193
URL: https://scialert.net/abstract/?doi=ajps.2010.183.193
INTRODUCTION
One of the considered influencing factors for the recent world food crisis is renewable energy from agriculture products. It has been criticized as a humiliation by diverting crops from food to bio-fuel feedstock and resources competition, water or land, leaving 800 million people in hunger or undernourishment worldwide. Second-generation bio-fuel technologies, based on lignocellulosic feedstock, have been proposed to reduce this competition. Native or external plant species with fast growing rate, high net energy yield and low input requirements, particularly capable of being grown on marginal lands, are being intensively researched such as sugarcane (Saccharum officinarum L.) (Rabelo et al., 2008) or switchgrass (Panicum virgatum L.) (Schmer et al., 2008). One potential energy crop is Sorghum bicolor (L.) Moench. (Reddy et al., 2005; Corredor et al., 2008). In addition, land use intensification such as sequential cropping, intercropping or crop rotation of energy crop with food crop is one of the solutions (Malezieux et al., 2009). However, in the tropical regions, most of the crops grown during the summer-rainy season frequently suffer intermittent or long-term waterlogging or flooding due to excess irrigation, storms, poor soil drainage or overflowing of the rivers. If this energy crop is incorporated into the paddy fields, a massive occupied in tropical and sub-tropical areas, waterlogging has a particularly heavy effect on the crop, since repeated puddling breaks capillary pores, reduces void ratio, destroys soil aggregates and disperses fine clay particles as well as when a rising water table and rainfall intensity combine with low evapotranspiration (Polthanee, 1997).
These lead energy crops to be more commonly subjected to waterlogging or flooding. Therefore, information about sorghum responses to waterlogging is essential.
Growth and physiological processes are detrimentally affected by waterlogging. Waterlogging decreases the leaf elongation rate and leaf area of plants (Orchard and Jessop 1984; Dias-Filho and de Carvalho, 2000; Malik et al., 2001, 2002; Henshaw et al., 2007) and consequently reduces plant height and ultimately suppresses root and shoot production (Haung et al., 1994).
Stomata closure, reduction of transpiration and inhibition of photosynthesis are common responses that can occur in hours or days, depending on the tolerance to waterlogging of each species and cultivar. When waterlogging is prolonged, waterlogging-intolerant plants such as wheat (Triticum aestivum) (Malik et al., 2001) or maize (Zea mays L.) (Zaidi et al., 2003) drastically reduce their physiological activities and are often killed in a short time, whereas, in waterlogging-tolerant plants the same parameters could even be enhanced or have less effect due to the ability of roots to acclimate to waterlogging, such as by the ability to produce adventitious roots and aerenchyma formation (Dias-Filho and de Carvalho, 2000; Pang et al., 2004; Striker et al., 2005; Irving et al., 2007; Li et al., 2007; Mollard et al., 2008).
Growth related to physiological response under waterlogging stress in sorghum has not yet been elucidated. Understanding how plants respond to waterlogging is important in determining their potential for use in habitat prone to this stress.
Therefore, the objectives of this study were to: (1) investigate morpho- and physiological changes in sorghum (Sorghum bicolor L. Moench) in response to waterlogging; and (2) compare the waterlogging tolerance of four sorghum cultivars, Wray, Keller, Bailey and Supanburi 1(SP1) during early vegetative stage.
MATERIALS AND METHODS
The experiment was conducted at the Moorbank Botanical Garden at the University of Newcastle (UK) School of Biology, from February to April 2008 (54.987°N and 1.635°W ). Seeds of sweet sorghum (Sorghum bicolor (L.) Moench) cv. Wray, Keller, Bailey and forage sorghum (Sorghum bicolor (L.) Moench) cv. Suphanburi 1 (SP1) (Supanburi Field Crop Research Centre, Thailand) were sown in 15 cm diameter pots with 450 g of 6:2:1 (peat: unsterile sand: potting base) culture. The potting base contained 7.5% nitrogen, 3.6% phosphorus and 5.2% potassium. Prior to planting, pots were watered to Field Capacity (FC) and then three to five sweet sorghum seeds were drilled into each pot at 2 cm depth. At the two-leaf stage, plants were thinned to one plant per pot. Water was applied daily to FC by weighing random selected pots on a balance. Waterlogging was applied at the vegetative stage (five fully expanded leaves) of each cultivar (Vanderllip and Reeves, 1972). Pots containing growing plants were placed in a larger pot and water was applied daily to the soil surface for 20 days. Control pots were free-draining and were watered daily. To create similar growing conditions between waterlogged and control pots, control pots were also placed in the empty larger pots (Irving et al., 2007). Pots were arranged in environmentally controlled conditions, where the temperature was 13/26°C (min/max) and with a 14 h photoperiod with the additional light of 400-watt high-pressure sodium bulbs (Pang et al., 2004). The pots were arranged in a 2x4 factorial randomized complete block design with four replications of waterlogging treatment and two replications of control treatment (Irving et al., 2007).
Plant growth measurements: During the waterlogging period, the length of the youngest expanding leaf was measured daily using a ruler (Dias-Filho and da Carvalho, 2000). Plant height (cm plant-1), senescent leaf number per plant and leaf area (cm2 plant-1) were recorded at four-day intervals. A non-destructive leaf area measurement method was adopted, according to Wright (1981). At the end of the waterlogging treatment, 20 Days after Waterlogging (DAW), plants were destructively harvested. The length of the longest nodal root and nodal root number were recorded (Mcfarlane et al., 2004). Root, leaf and culm parts were dried at 80°C for 48 h and then the plant’s dry weight was recorded. Shoot: root biomass ratio was calculated (Ye et al., 2003).
Photosynthetic measurements: Photosynthetic rate (A, μmol m-2 sec-1), stomatal conductance (gs, mol m-2 sec-1) and transpiration rate (E, mmol m-2 sec-1) were collected with a LCi portable photosynthesis system (ADC Bioscientific Ltd., Hoddesdon, 2004, UK). Measurements were taken from the youngest fully expanded leaf of each plant (Pang et al., 2004). These measurements were made from 11.00 to 13.00 hrs, with the following specifications/adjustments: leaf surface area, 6.25 cm2; ambient CO2 concentration, 385 μmol mol-1; and temperature of leaf chamber varying from 32.37°C to 34.33°C. Photosynthetically Active Radiation (PAR), provided by 400-watt high-pressure sodium bulbs, was set at the leaf surface to 700 μmol m-2 sec-1 (Ashraf and Ur-Rehman, 1999). Measurements were conducted at 0, 4, 8, 12, 16 and 20 Days after Waterlogging (DAW).
Statistical analysis: The data for all physiological and growth parameters were subjected to Analysis of Variance (ANOVA) using Statistix 8 software (Anonymous, 2003). The mean values were compared using the least significant difference test.
RESULTS
Shoot growth response to waterlogging: Waterlogging significantly reduced plant height and the adverse effects were more pronounced when duration of waterlogging increased. At 4 Days after Waterlogging (DAW), cv. Wray and Bailey could maintain plant height to control plants but cv. Keller and SP1 showed a 10% reduction from their respective controls. However after 8 DAW onwards, a higher height reduction with longer waterlogging duration was found. At 20 DAW, height of Keller was reduced to 30% (Fig. 1c), Bailey was reduced to 33% (Fig. 1e) and SP1 was reduced to 35% (Fig. 1g) as compared to the control. In contrast, cv. Wray after 12 DAW onwards had height reduction maintained at 18-20% lower than the control (Fig. 1a). Reduction in plant height was consistent with the decrease in Youngest Leaf Expansion Rate (YLER). Waterlogging significantly reduced YLER of all cultivars, being lowest in cv.
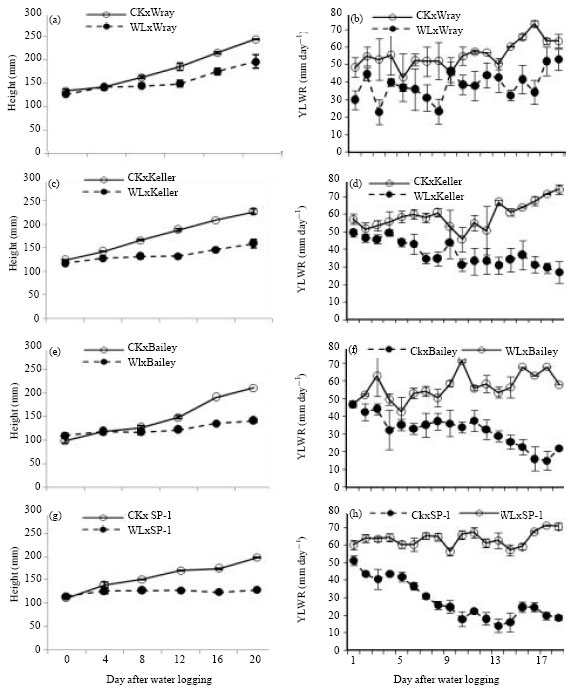 | |
| Fig. 1: | Effect of waterlogging on plant height (a, c, e, g) and Youngest Leaf Expansion Rate (YLER) (b, d, f, h) of 4 sorghum cultivars. Solid lines and open symbols represent control plants, broken lines and closed symbols represent waterlogged plants. Error bars represent standard errors |
Wray (Fig. 1b), while progressively decreasing in cv. Keller (Fig. 1d), Bailey (Fig. 1f) and being highest in SP1 (Fig. 1h). LA was significantly affected by waterlogging, with advancing reduced when waterlogging duration was extended. Different responses were noted among studied cultivars. Cv. Wray could maintain LA similar to control plants at 4 DAW and began to reduce at 8 DAW while the other three cultivars started to decline at 4 DAW onwards. After 12 DAW to 20 DAW LA reductions in cv. Wray were between 56-65% (Fig. 2a). However, in cv. Keller it increased from 43% at 12 DAW to 62% at 20 DAW (Fig. 2c), or 34 to 72% in cv. Bailey (Fig. 2e) and the highest persistence was in cv. SP1 (74 to 80%) (Fig. 2g). The reduction in LA was concurrent with the acceleration in leaf senescence. Waterlogging significantly increased the senescent leaf number of all studied cultivars in terms of the onset and change during the waterlogging period. Senescent leaf No. gradually increased at 8 DAW in cv. Wray and Bailey and reached a maximum at 16 DAW and 20 DAW, respectively (Fig. 2b and f). However, a sharp rise to maximum was noted in cv. Keller at 8 DAW onwards (Fig. 2d). Nevertheless, the development of senescent leaf No. was delayed in cv. SP1, started at 12 DAW. Shoot biomass was severely affected by 20 days of waterlogging with 71% compared to the control.
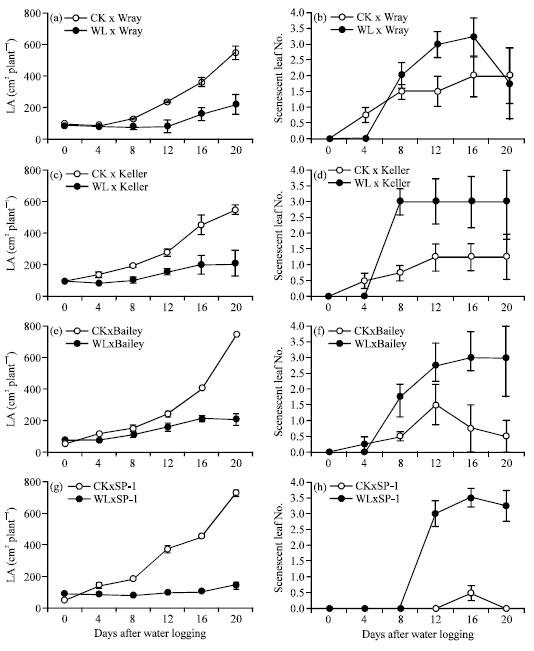 | |
| Fig. 2: | Effect of waterlogging on leaf area, LA, (a, c, e, g) and leaf senescent number (b, d, f, h) of 4 sorghum cultivars. Solid lines and open symbols represent control plants, broken lines and closed symbols represent waterlogged plants. Error bars represent standard errors |
This was due to the decrease in culm dry weight (DW) (68%) and leaf DW (72%). All cultivars had a similar response and no interaction between water regimes and cultivars was found (Table 1). However, under waterlogging conditions, cv. Wray showed relatively higher shoot DW, followed by cv. Keller and Bailey and lowest in cv. SP1.
Root growth: Waterlogging significantly reduced Nodal Root (NR) number plant-1, longest root length (LRL) and consequent root DW, compared to the control plants. NR number plant-1, LRL and root DW were reduced to 24, 28 and 71% compared to the control plants, respectively (Table 1). The differences in cultivar responses were found in all root growth parameters. Interestingly, compensation between NR number and LRL was observed. The lowest NR number was found in cv. Wray (16 plant-1), but it had the longest LRL (399 mm plant-1) (Table 1). Interactions between water regimes and cultivars on NR number and LRL were also noted. Cv. Bailey had the highest NR number (30 plant-1), whereas the lowest was found in waterlogged Wray (13 plant-1) (Fig. 3a). LRL was highest in the control treatment of cv. Wray (502.75 mm plant-1) and lowest in waterlogged cv.
| Table 1: | Effects of 20 days of waterlogging on above-ground and below-ground growths of 4 sweet sorghum cultivars |
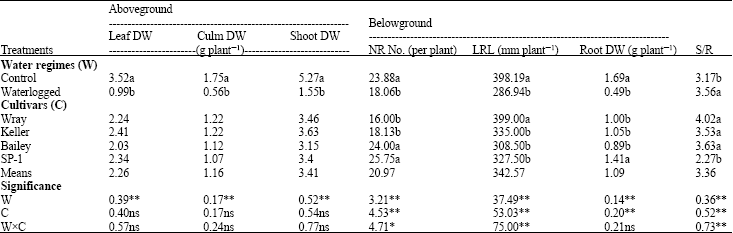 | |
| *,**Significant at p<0.05 or 0.01, respectively and ns; not significantly different at 0.05 of probability. Values followed by the same letter in the same column are not significantly different at 0.05 and 0.01 probability. DW: dry weight | |
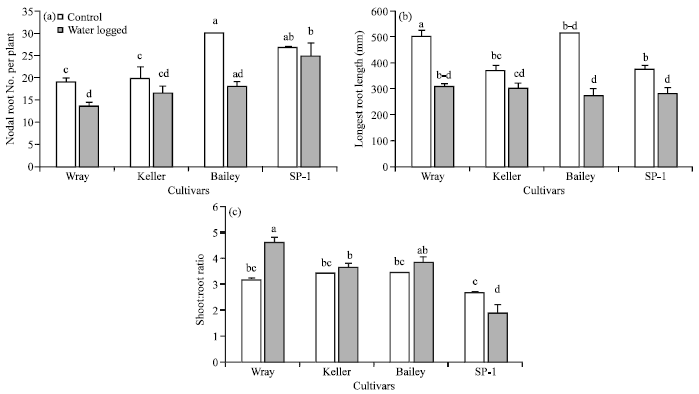 | |
| Fig. 3: | (a) Effects of 20 days of waterlogging on nodal root number per plant, (b) longest root length and (c) shoot/root ratio of 4 sorghum cultivars. Values followed by the same letter in each growth stage are not significant different at 0.05 and 0.01 probability. Error bars represent standard error |
Bailey (272 mm plant-1) (Fig. 3b). However, no interaction was noted in terms of root DW.
Shoot/root ratio: Shoot /root ratio (S/R) was significantly higher (3.56) in plants under waterlogged conditions than in freely drained pots (3.17) (Table 1). A statistically different interaction was found. Further analysis showed that waterlogged SP1 had the lowest S/R, while the highest was found in waterlogged Wray (Fig. 3c).
Physiological response to waterlogging: Leaf gas-exchange characteristics of all sorghum cultivars were significantly affected by waterlogging varying in cultivars studied. The intensification was more pronounced with longer duration of waterlogging. Photosynthetic rate (A) of sweet sorghum, cv. Wray, Keller and Bailey, started to decline at 4 DAW, with 29, 58 and 31%, respectively, compared to their controls but unchanged in cv. SP1. The degree of the reduction was maintained to 8 DAW and accelerated to 57% at 12 DAW and was lowest at 20 DAW 78% in cv. Wray (Fig. 4a). In cv. Keller, the adverse effect was alleviated at 8 DAW (35%) and 12 DAW (40%). However, prolonged waterlogging duration reduced A of Keller to 65-70% (Fig. 4c). In cv. Bailey, the response was found similar to cv. Wray (Fig. 4e). In contrast to sweet sorghum, waterlogging reduced A of forage sorghum, cv.
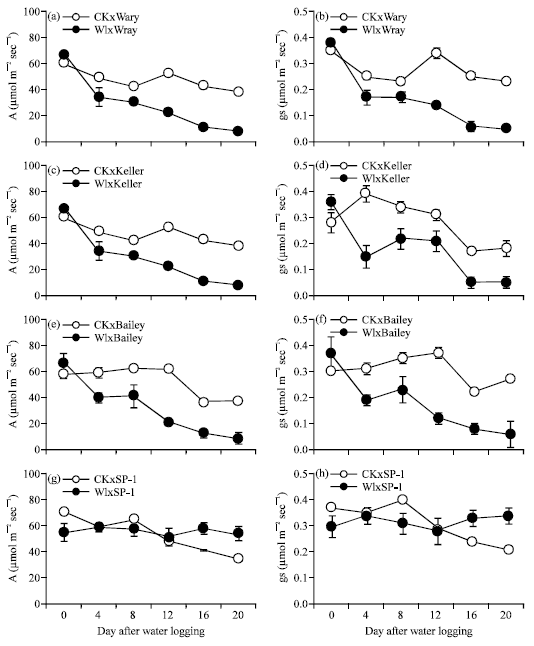 | |
| Fig. 4: | Effect of waterlogging on photosynthetic rate (A) (a, c, e and g) and stomatal conductance (gs) (b, d, f and h) of 4 sorghum cultivars. Solid lines and open symbols represent control plants, broken lines and closed symbols represent waterlogged plants. Error bars represent standard errors |
SP1 only at 8 DAW (11%). And starting from 12 DAW onwards, A was increased over the control, giving 56% higher than controls at 20 DAW (Fig. 4h).
Stomatal conductance (Fig. 4b, d, f and h) and transpiration rate (data not shown) followed a similar pattern to that observed for photosynthesis.
DISCUSSION
Four sorghum genotypes in this experiment responded to waterlogging similarly in terms of shoot and root biomass accumulations but study of root growth, dry matter partition and leaf gas exchange parameter showed drastic differences in their responses.
The significant shoot growth reduction (71%) was due to the restricted development of plant height (30%), LA (69%) and thus decreased dry matter accumulation in leaf (72%) and culm (68%). This finding is in agreement with the findings in wheat (Triticum aestivum) (Malik et al., 2001), mungbean (Vigna radiate L. Wilczek) (Ahmed et al., 2002), ryegrass (Lolium perenne L.) (Mcfarlane et al., 2004), maize (Zea mays L.) (Zaidi et al., 2003, 2004), barley (Hordeum vulgare) (Pang et al., 2004), buckwheat (Fagopyrum sp.) (Matsuura et al., 2005) and soybean (Glycine max (L.) Merrill ) (Henshaw et al., 2007).
The pronounced reduction in shoot growth in this study is described by its high susceptibility to waterlogging at vegetative stage of S. bicolor due to its lack of nodal root development as compared to other growth stages (Our experiment, unpublished data). In this study, even though plant height and leaf area were reduced, waterlogging did not cause a significant reduction to biomass of shoot and root among cultivars. This indicates that these cultivars are tolerant to waterlogging conditions. Our previous experiment demonstrated that continuous flooding sweet sorghum from 30 DAE (8-10 leaf stage) until harvest only reduced shoot biomass and stalk yield to 20 and 22%, respectively. However, to investigate up to what extent they can withstand continuous flooding at the early vegetative stage further experiment is required. No significant difference in shoot biomass but significant reduction in shoot length and LA in response to waterlogging is also reported in Hibicus esculentus (Ashraf and Arfan, 2005).
The present study showed that even with a lack of differences in total biomass accumulation between aerobic and anaerobic treatment, significant differences in biomass allocation patterns was found. This is consistent with the response to waterlogging of Sporobolus virginicus (L.) Kunth (Naidoo and Mundree, 1993).
Results of this experiment indicate that the biomass partitioned to root in cv. SP1 is utilized for the initiation of new nodal roots. However, there was a competition for assimilates between shoot and root growth, resulting in slow shoot growth. This is in contrast to cv. Wray, where more assimilates was allocated to support shoot growth. This supports the result of Ye et al. (2003), who indicated that a shift of biomass from root to shoot is an adaptation to prolonged waterlogging in the higher waterlogging tolerance mangrove species (Kandelia candel). The significant reduction in photosynthate allocation to belowground parts, while maintaining aboveground biomass accumulation is reported as an effective metabolic strategy to reduce belowground oxygen demand and to increase the potential of shoots to transport oxygen to root of Sporobolus virginicus (L.) Kunth (Niadoo and Mundree, 1993). Therefore, it could possibly be concluded that in sorghum biomass portioning to shoot is an acclimation response to long-term waterlogging at early growth stage.
Leaf elongation rate has been proposed as an early detection mechanism for relative flood tolerance in grass species such as Brachiaria spp. (Dias-Filho and de Carvalho, 2000; Dias-filho, 2002) or maize (Zea mays L.) (Lizaso and Ritchie, 1997). Our results show that cv. Wray has the ability to extend the youngest leaf and produce new leaves, indicating that this cultivar is relatively tolerant to waterlogging. However in cv. SP-1, YLER was sharply decreased at day 2 after applying waterlogging and continued decreasing over time, indicating that cv. SP-1 is relatively intolerant to waterlogging. Cv. Keller (as well as cv. Bailey) has the ability to maintain YLER for at least five days after waterlogging and then gradually decrease. This may imply that these two cultivars are quite tolerant to short-term waterlogging. Coincident increasing in senescent leaf number of cv. Wray, Keller and Bailey during waterlogging implies that these new growths may be supported by the remobilization of nutrients from older parts. This finding is consistent with the response of lucerne (Medicago sativa) to waterlogging (Irving et al., 2007).
Adventitious root or nodal root development has been reported as the key root acclimation to waterlogging or flooding (Pardales et al., 1991; McDonald et al., 2002; Pang et al., 2004; Polthenee et al., 2008; Changdee et al., 2009). However, Van Noordwijk and Brouwer (1993) suggested that more developed roots may be less able to adapt morphologically (such as development of aerenchyma) under stress conditions. A relatively extensive aerenchyma spaces noted in cv. Wray than cv. SP1 in previous experiment may support the previous concept.
Zaidi et al. (2003) indicated that an early adventitious rooting is one of the crucial morphological traits of maize to tolerate excess soil moisture stress. This is in agreement with our results, indicating that sweet sorghum possesses this root acclimation trait, while fibre sorghum does not. In addition, our results showed that in cv. Wray even with lowest nodal root number, it had significant highest individual root length. This may presumably be a partial compensation to conserve root area for water and nutrient uptake during waterlogging. It was also observed that during waterlogging, the nodal root of cv. Wray was located near the soil surface indicating that this cultivar possesses root acclimation to reach atmospheric oxygen. This supports the results of Niadoo and Mundree (1993), who indicated that waterlogging tolerant Sporobolus virginicus (L.) possesses morphological responses to waterlogging such as fewer, but taller and more mature culm with greater aerenchyma spaces and production of aboveground adventitious roots close to the aerobic zone.
Thus, the present experiment indicates that the early nodal root development, the ability to maintain root surface area for water and nutrient uptake and to develop aerenchyma spaces in existing roots and nodal root development near the soil surface during the susceptible stage are root morphological acclimations to survive and concurrently sustain plant growth under prolonged waterlogging conditions.
The decreased photosynthetic rate, as well as transpiration rate, in our experiment may be partially regulated by stomatal closure, due to a high positive correlation between stomatal conductance and photosynthesis under both control and waterlogged conditions (Fig. 5). Other findings (Huang et al., 1994; Malik et al., 2001; Ashraf, 2003; Striker et al., 2005) have also shown that stomatal conductance is the major factor effecting photosynthesis under waterlogging conditions in plants.
The lower leaf photosynthesis of cv. Wray, Keller and Bailey may also possibly be regulated by sink strength in terms of root growth, since these cultivars have relatively low nodal root numbers compared to significantly higher ones in cv. SP1. Strong regulation of leaf photosynthesis by sink strength in wheat (Triticum aestivum) (Malik et al., 2002) and sugarcane (Saccharum officinarum L.) (McCormick et al., 2006) has also been reported. In this study, a positive significant correlation coefficient between photosynthetic rate and nodal root number of cv. SP1 may support this finding (Fig. 6). In addition, factors regulating photosynthesis in plants grown in waterlogged soil may be reduced CO2 transfer conductance from sub-stomatal cavities to the site of carboxylation, or activity of photosynthetic enzymes at the carboxylation point.
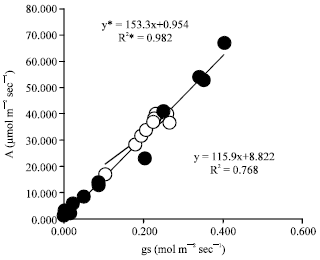 | |
| Fig. 5: | Relationships between photosynthetic rate (A) and stomatal conductance (gs) of sorghum. Open symbols represent control plants and filled symbols represent waterlogged plants. Asterisks represent responses in waterlogged plants |
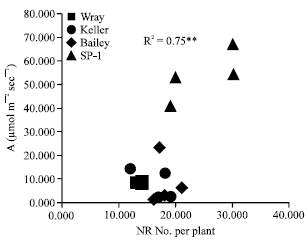 | |
| Fig. 6: | Relationships between NR number per plant and photosynthetic rate (A) under waterlogging conditions |
A marked increase in sub-stomatal CO2 (Ci) noted in waterlogged cv. Wray, Keller and Bailey in relation to control plants, compared with comparative levels between waterlogged and freely drained plant in cv. SP1 (data not shown), may confirm that point. This is consistent with the finding of Malik et al. (2001), who reported that this may be responsible for decreased photosynthesis in waterlogged wheat. However, to draw such relationship in sorghum in response to waterlogging, a further experiment is needed. Nevertheless, the observed response of photosynthesis to CO2 indicates that a relevant limitation to photosynthesis of S. bicolor under waterlogging conditions may be caused by a reduced capacity of RuBisCo for CO2 fixation, not translocation of CO2 such as occurs in drought stressed sweet sorghum (Massacci et al., 1996). Reduction of RuBisCO is closely related to total soluble protein content (Irving et al., 2007) and is logically related to total leaf nitrogen content. The concurrent higher SPAD Chlorophyll Meter Reading (SCMR) and photosynthetic rate (A) in cv. SP1, as well as a significant decrease in SCMR and A of cv. Wray, Keller and Bailey, throughout the experiment (data not shown), possibly implies that the leaf photosynthetic rate in waterlogged sorghum is partly regulated by chlorophyll.
However, in sweet sorghum, particular cv. Wray, it is likely that the growth did not relate to chlorophyll. This is consistent with the finding in Blue panicgrass response to waterlogging (Ashraf, 2003). This may be due to the remobilization of nitrogen from older plant parts to support shoot growth during waterlogging causing a reduction in photosynthetic rate. Under waterlogging or flooding conditions, a positive relation between photosynthetic capacity and growth has been reported in signal grass (Brachiaria brizantha (A. Rich.) Stapf) (Dias-filho, 2002), Blue panicgrass (Panicum antidotale Retz.) (Ashraf, 2003), barley (Hordeum vulgare) (Pang et al., 2004), bird's-foot trefoils (Lotus sp.) (Striker et al., 2005) or dallisgrass (Paspalum dilatatum) (Mollard et al., 2008), due to the development of nodal roots or adventitious roots, which form extensive aerenchyma spaces which create a relatively low-resistance internal pathway and by enhancing mass flow transport of oxygen, carbon dioxide and ethylene between plant parts above water and submerged tissues (Jackson and Colmer, 2005). This is in contrast to our findings. The present study showed that under waterlogging conditions cv. SP1 produces nodal roots similar to control plants and increased photosynthetic rate higher than the control at the end of the experiment. However, its height, LA, leaf DW and shoot DW had highest decrease. In contrast to cv. Wray, waterlogging significantly reduced nodal root number and photosynthetic rate. However, its height, Youngest Leaf Expansion Rate (YLER), LA and shoot biomass was less affected than cv. SP1. This indicates that there is no relationship between growth and photosynthetic capacity in sorghum under waterlogging conditions. This supports the results of Ashraf and Arfan (2005), who found that under waterlogging Okra (Hibiscus esculentus) decreases photosynthetic rate but not its shoot biomass or wheat (Triticum aestivum) under salt stress (Hawkins and Lewis, 1993). The concomitant reduction in shoot biomass and unaffected leaf photosynthetic rate is found in Sporobolus virginicus (L.) Kunth in response to the combination effects of waterlogging and salt stress (Naidoo and Mundree, 1993), which is consistent to the response of forage sorghum, cv. SP1, in this experiment.
Nevertheless, in comparison to wheat response to severe waterlogging (Malik et al., 2001), photosynthesis is reduced to 82% compared with the control at 5 d after waterlogging. Our results indicate that sorghum has higher waterlogging tolerance than wheat since it took 20 days to suppress photosynthesis to a similar value (average 73% as compared to the control plants). Significantly higher plant height, or the ability to extend the youngest leaf expansion rate per day, results in relatively higher shoot biomass during waterlogging in cv. Wray and it can thus be concluded that cv. Wray may be the most waterlogging tolerant from an agronomic point of view, the maintenance of relatively high yield (Setter and Waters, 2003). Cv. Keller and Bailey are intermediate and cv. SP1 is sensitive to waterlogging. This indicates that sweet sorghum; especially cv. Wray has the potential to grow on waterlogging prone areas, whereas forage sorghum is more preferable on upland areas. In cv. SP1, leaf photosynthetic rate was less affected by short-term waterlogging and during the long-term waterlogging duration leaf photosynthetic rate as well as transpiration rate and stomatal conductance increased over the control. It can thus be concluded that cv. SP1 is the most waterlogging tolerant from a physiological point of view, survival or maintenance of high growth rate under waterlogging, relative to non-waterlogged conditions (Setter and Waters, 2003). Cv. SP1 may be the most valuable cultivar for use in further breeding programs. At the same time, from an agronomic point of view, cv. Wray may be the most suitable for immediate use by agronomists. Therefore, it can be seen that physiological measurements can help in assessing highly physiology tolerant cultivars, which only a high-yield basis assessment may simply discard.
In conclusion, waterlogging reduced plant growth of four sorghum genotypes similarly but study of root growth, dry matter partition and leaf gas exchange parameter showed drastic differences in their responses. Cultivar, maintaining the higher shoot growth, was not associated with the ability to develop higher nodal root and retain or least affect leaf photosynthetic rate. But it is related to biomass partitioning to shoot during long- term flooding, early nodal root development, accompanied with the ability to conserve root surface area for water and nutrient uptake as well as the ability to remobilize nutrients from older parts to support shoot growth such as cv. Wray. In contrast, keeping high photosynthetic rate by partitioning biomass to develop new roots was at the expense of shoot such as cv. SP1. Nevertheless, to comprehend the physiological response of this cultivar, further study is needed.
ACKNOWLEDGMENTS
This research was financially supported by the Royal Golden Jubilee program of the Thailand Research Fund. The authors acknowledge the kind assistance of Brian Brown, technical manager from the School of Agriculture, Food and Rural Development and Moorbank Botanical Garden, School of Biology, University of Newcastle, UK, for providing equipment and facilities during the conduction of the experiment.
REFERENCES
- Ahmed, S., E. Nawata and T. Sakuratani, 2002. Effects of waterlogging at vegetative and reproductive growth stages on photosynthesis, leaf water potential and yield in mungbean. Plant Prod. Sci., 5: 117-123.
CrossRefDirect Link - Ashraf, M. and H. Ur-Rehman, 1999. Interactive effects of nitrate and long-term waterlogging on growth, water relations and gaseous exchange properties of maize (Zea mays L.). Plant Sci., 144: 35-43.
Direct Link - Ashraf, M., 2003. Relationships between leaf gas exchange characteristics and growth of differently adapted populations of Blue panicgrass (Panicum antidotale Retz.) under salinity or waterlogging. Plants Sci., 165: 69-75.
CrossRefDirect Link - Ashraf, M. and M. Arfan, 2005. Gas exchange characteristics and water relations in two cultivars of Hibicus esculentus under waterlogging. Biol. Plant, 49: 459-462.
CrossRefDirect Link - Corredor, D.Y., J.M. Salazar, K.L. Hohn, S. Bean, B. Bean and D. Wang, 2008. Evaluation and characterization of forage sorghum as feedstock for fermentable sugar production. Applied Biochem. Biotechnol., 158: 164-179.
CrossRef - Dias-Filho, M.B. and C.J.R. de Carvalho, 2000. Physiological and morphological responses of Brachiaria spp. to flooding. Pesq. Agropec. Bras., 35: 1959-1966.
CrossRef - Dias-Filho, M.B., 2002. Tolerance to flooding in five Brachiaria brizantha accessions. Pesq. Agropec. Bras., 37: 439-447.
CrossRef - Hawkins, H.J. and O.A.M. Lewis, 1993. Combination effect of NaCl salinity, nitrogen form and calcium concentration on the growth, ionic content and gaseous exchange properties of Triticum aestivum L. cv. Gamtoos. New Phytol., 124: 161-170.
CrossRefDirect Link - Henshaw, T.L., R.A. Gilbert, J.M.S. Scholberg and T.R. Sinclair, 2007. Soya bean (Glycine max L. Merr.) genotype response to early-season flooding: II. Aboveground growth and biomass. J. Agron. Crop Sci., 193: 189-197.
CrossRefDirect Link - Huang, B., J.W. Johnson, D.S. Nesmith and D.C. Bridges, 1994. Growth, physiological and anatomical responses of two wheat genotypes to waterlogging and nutrient supply. J. Exp. Bot., 45: 193-202.
CrossRefDirect Link - Irving, L.J., Y.B. Sheng, D. Woolley and C. Matthew, 2007. Physiological effects of waterlogging on two lucern varieties grown under glasshouse conditions. J. Agron. Crop Sci., 193: 345-356.
CrossRefDirect Link - Jackson, M.B. and T.D. Colmer, 2005. Response and adaptation by plants to flooding stress. Ann. Bot., 96: 501-505.
CrossRefDirect Link - Li, M., D. Yang and W. Li, 2007. Leaf gas exchange characteristics and chlorophyll fluorescence of three wetland plants in response to long-term soil flooding. Photosynthetica, 45: 222-228.
CrossRef - Lizaso, J.I. and J.T. Ritchie, 1997. Maize shoot and root response to root zone saturation during vegetative stage. Agron. J., 89: 125-134.
Direct Link - Malezieux, E., Y. Crozat, C. Dupraz , M. Laurans and D. Makowski et al., 2009. Mixing plant species in cropping systems: Concepts, tools and models: A review. Agron. Sustain. Dev., 29: 43-62.
CrossRef - Malik, A.I., D.T.D. Colmer, H. Lambers and M. Schortemeyer, 2001. Changes in physiological and morphological traits of roots and shoots of wheat in response to different depth of waterlogging. Aust. J. Plant Physiol., 28: 1121-1131.
CrossRef - Malik, A.I., T.D. Colmer, H. Lamber, T.I. Settler and M. Schortmeyer, 2002. Short-term waterlogging has long-term effects on the growth and physiology of wheat. New Phytol., 153: 225-236.
CrossRefDirect Link - Massacci, A., A. Battistelli and F. Loreto, 1996. Effect of drought stress on photosynthetic characteristics, growth and sugar accumulation of field-grown sweet sorghum. Aust. J. Plant Physiol., 23: 331-340.
CrossRefDirect Link - Matsuura, A., S. Inanage, T. Tetsuka and K. Murata, 2005. Differences in vegetative growth response to soil flooding between Common and Tartary Buckwheat. Plant Prod. Sci., 8: 525-532.
Direct Link - McDonald, M.P., N.W. Galwey and T.D. Colmer, 2002. Similarity and diversity in adventitious root anatomy as related to root aeration among a range of wetland and dryland grass species. Plant Cell Environ., 25: 441-451.
Direct Link - Mcfarlane, N.M., T.A. Ciavarella and K.F. Smith, 2004. The effects of waterlogging on growth, photosynthesis and biomass allocation in perennial ryegrass (Lolium perenne L.) genotypes with contrasting root development. J. Agric. Sci., 141: 241-248.
Direct Link - Mollard, F.O.P., G.G. Striker, E.L. Polschuk, A.S. Vega and P. Insausti, 2008. Flooding tolerance of Paspalum dilatatum (Poaceae: Paniceae) from upland and lowland positions in a natural grassland. Flora, 203: 548-556.
CrossRef - Naidoo, G. and S.G. Mundree, 1993. Relationship between morphological and physiological responses to waterlogging and salinity in Sporobus virginicus (L.) Kunth. Oecologia, 93: 360-366.
CrossRef - Orchard, P.W. and R.S. Jessop, 1984. The response of sorghum and sunflower to short-term waterlogging. I: Effects of stage of development and duration of waterlogging on growth and yield. Plant Soil, 81: 119-132.
CrossRef - Pang, J., M. Zhou, N. Mendham and S. Shabala, 2004. Growth and physiological responses of six barley genotypes to waterlogging and subsequent recovery. Aust. J. Agric. Res., 55: 895-906.
CrossRef - Pardales, Jr.J.R., Y. Kono and A. Yamauchi, 1991. Response of the different root system components of sorghum to incidence of waterlogging. Environ. Exp. Bot., 31: 107-115.
CrossRef - Polthenee, A., T. Changdee, J. Abe and S. Morita, 2008. Effects of flodding on growth, yield and aerenchyma development in adventitious roots in four cultivars of kenaf (Hibiscus cannabinus L.). Asian J. Plant Physiol., 7: 544-550.
CrossRefDirect Link - Rabelo, S.C., R.M. Filho and A.C. Costa, 2008. Lime pretreatment of sugarcane bagasse for bioethanol production. Applied Biochem. Biotechnol., 153: 139-150.
CrossRef - Reddy, B.V.S., S. Ramesh, R.P. Sanjana, P.M. Ramaih and K. Rajashekar, 2005. Sweet sorghum: A potential alternative raw material for bioethanol and bio-energy. Int. Sorghum Mill. Newslett., 46: 79-86.
Direct Link - Schmer, M.R., K.P. Vogel, R.B. Mitchell and R.K. Perrin, 2008. Net energy of cellulosic ethanol from switchgrass. Proc. Natl. Acad. Sci. USA, 105: 464-469.
CrossRefDirect Link - Setter, T.L. and I. Waters, 2003. Review of prospects for germplasm improvement for waterlogging tolerance in wheat, barley and oats. Plant Soil, 253: 1-34.
CrossRef - Striker, G.G., P. Insausti, A.A. Grimoldi, E.L. Ploschuk and V. Vasellati, 2005. Physiological and anatomical basis of differential tolerance to soil flooding of Lotus corniculatus L. and Lotus glaber. Mill. Plant Soil, 276: 301-311.
CrossRef - Vanderllip, R.L. and H.E. Reeves, 1972. Growth stages of sorghum (Sorghum bicolor (L.) Moench. Agron. J., 64: 13-16.
Direct Link - Van Noordwijk, M. and G. Brouwer, 1993. Gas-filled root porosity in response to temporary low oxygen supply in different growth stages. Plant Soil, 152: 187-199.
CrossRef - Ye, Y., N.F.Y. Tam, Y.S. Wong and C.Y. Lu, 2003. Growth and physiological responses of two mangrove species (Bruguiera gymnorrhiza and Kandelia candel) to waterlogging. Environ. Exp. Bot., 49: 209-221.
CrossRefDirect Link - Zaidi, P.H., S. Rafique and N.N. Singh, 2003. Response of maize (Zea mays L.) genotypes to excess soil moisture stress: Morpho-physiological effects and basis of tolerance. Eur. J. Agron., 19: 383-399.
CrossRef - Zaidi, P.H., S. Rafique, P.K. Rai, N.N. Singh and G. Srinivasan, 2004. Tolerance to excess moisture in maize (Zea mays L.): Susceptible crop growth stage and identification of tolerant genotypes. Field Crops Res., 90: 189-202.
CrossRef - Changdee,T., A. Polthanee, C. Akkasaeng and S. Morita, 2009. Effect of different waterlogging regimes on growth, some yield and roots development parameters in three fiber crops (Hibiscus cannabinus L., Hibiscus sabdariffa L. and Corchorus olitorius L.). Asian J. Plant Sci., 8: 515-525.
CrossRefDirect Link