
P. Sharifi
Department of Plant Breeding and Agronomy, Faculty of Agriculture, Islamic Azad University, Rasht, Iran
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 5 | Page No.: 485-489







ABSTRACT
Present study determined the inheritance and heritability of cold tolerance at the germination stage in crosses between seven rice genotypes by partial diallel crosses between them. Analysis of variance indicated that there were highly significant differences among the replications, genotypes, General Combining Ability (GCA) and Specific Combining Ability (SCA) for percentage of reduction in Radicle Length (RL), percentage of reduction in plumule length (PR) and percentage of reduction in germination measure at percentage (GP). For all of three traits the higher magnitude of the dominance variances showed that non-additive gene action was predominant to additive one. The perusal of GCA effects reveals that Deilamani was an ideal general combiner for all characters and this parent is superior variety for them, whereas Hassani was a good general combiner for PL and GP. The estimates of SCA effects of top ranking crosses indicated that Neda X Hassani could be regarded as the most desirable cross combination for all of three traits. In this cross significant negative SCA effect indicated, demonstrating the existence of heterosis for cold tolerance.
PDF Abstract XML References Citation
How to cite this article
P. Sharifi, 2008. Inheritance of Cold Tolerance in Rice at the Germination Stage. Asian Journal of Plant Sciences, 7: 485-489.
DOI: 10.3923/ajps.2008.485.489
URL: https://scialert.net/abstract/?doi=ajps.2008.485.489
DOI: 10.3923/ajps.2008.485.489
URL: https://scialert.net/abstract/?doi=ajps.2008.485.489
INTRODUCTION
Rice (Oryza sativa L.), is a cold sensitive crop that has its origin in tropical or sub-tropical areas and low temperatures dramatically reduce its production (Kuroki et al., 2007). During the early growth stages of rice, the occurrence of low temperature stress inhibits germination, seedling establishment and eventually leads to non uniform crop maturation. Good cold tolerance at the seedling stage is an important character for stable rice production, especially in direct seeding fields (Cruz and Milach, 2004). Developing cold-tolerant variety is one of the most effective ways to avoid the low-temperature damage (Lou et al., 2007). For rice temperatures lower than 20°C decrease both the speed and percentage of germination (Xu, 2008).
Diallel mating designs are an important tool in plant breeding programs to obtain information on the inheritance of quantitative traits. A partial diallel consisting of crosses between numbers of parents is useful when there are distinct characters (Griffing, 1956). Diallel analysis provides information on average performance of individual lines in crosses known as General Combining Ability (GCA). It also gives information about the performance of crosses relative to the average performance of parents involved in the cross known as Specific Combining Ability (SCA). Heritability is defined as the ratio of variance due to hereditary difference and genotypic variance to the total phenotypic variance (Cruz et al., 2006). Diallel analysis leads to identification of parents with additive and non additive effects for specific characteristics. This in turn helps in choosing parents to be included in hybridization or population breeding programs (Murtaza et al., 2005).
Breeding programs for cold tolerance use cold tolerant donor parents of diverse origin, appropriate breeding methods, suitable selection criteria and reliable screening and testing techniques in laboratories, glasshouses and fields (Farrell et al., 2006) described tolerance screening at germination and discovered critical temperatures were 10°C for 9 day at germination stage. Prianka et al. (2000) studied genetic variability in 26 rice genotypes to identify traits associated with cold tolerance and indicated high heritability for germination percentage and moderate heritability for shoot: root ratio. The few studies conducted on diallel analysis of cold tolerance in rice are related to the vegetative stage (Cruz and Milach, 2004; Prianka et al., 2000; Cruz et al., 2006; Han et al., 2006). The mode of inheritance is also very important in defining the selection strategy to be applied in a breeding program. There are a few studies related to the genetic basis of rice cold tolerance at the germination stage, with the main available information being the papers published by Sthapit and Witcombe (1998) and Takahashi (1997). The data from these reports indicate that the low temperature germination of rice is controlled by four or more dominant genes and those both additive and non-additive effects are involved, although transgressive segregation and the absence of a maternal effect were also reported. Also Cruz et al. (2006) due to a diallel analysis showed that both additive and non-additive gene action were involved in cold tolerance traits at germination stage, such as percentage of reduction in coleoptile length and coleoptile growth, while the non-additive action was relatively more important for them.
The objectives of this study were to determine the general and specific combing ability of seven rice genotypes, the choice of best parents (between studied parents) for cold resistance programs and the inheritance and heritability of cold tolerance at the germination stage and.
MATERIALS AND METHODS
Genetic material and crosses: The experiment carried out in the farm of Rice Research Institute of Iran (RRII) in Rasht, which were located between 49°E longitude and 37°N latitude at an altitude of 7 m below the mean sea level in north of Iran. Seven rice (Oryza sativa L.) varieties containing Deilmani, Hassani, Shahpasand, Saleh, Sepidrod, Neda and IRFAON-215 were crossed in a partial diallel at the 2006. The F1 seeds from seven parents were grown during the spring 2007 and the F1 plants were further advanced to the F2 generation. Reciprocal crosses were not performed because there was no indication of maternal inheritance for cold reaction (Takahashi, 1997). The cold tolerance evaluation was carried out in laboratory of department of agronomy and plant breeding in Islamic Azad University of Rasht.
Cold tolerance evaluation: Rice cold tolerance at germination was considered by the method of Cruz et al. (2006) that evaluated the percentage of reduction in Radicle Length (RL) and Plumule Length (PL) and germination measure at percentage (GP) due to cold after a cold stress. Before the beginning of the experiments, all the seeds of the parents and F2 were washed with aqueous ethanol (70% v/v) for 30 sec followed by immersion in aqueous sodium hypoclorite (5% v/v) for 20 min to prevent contamination and then washed six times with sterile distilled water. Traits was measured by placing 30 seeds of each of the seven parents and 21 F2 hybrids in Petri dishes containing germination paper moistened with sterile distilled water and germination allowed to proceed at 25°C for seven days (control) and 11°C for 30 days (cold treatment). In each of these treatments the experiment was conducted in a randomized block design with three replicates, where the blocks corresponded to different shelves in the germination chamber and each replication was the mean of the measurements for the seeds in a same Petri dish. At the end of each germination period the radicle and plumule length and germination of each seed was measured and the final data were expressed as percentage of reduction in these traits, obtained by comparing the cold-treated seeds with the control seeds for each genotypes.
Statistical analysis: Data for percentage of reduction in radicle and plumule length and germination for the genotypes was normally distributed and were thus submitted to analysis of variance and comparison of means using LSD test (α = 0.01).
Data were analyzed using SAS (SAS, 2002). Analysis of Cold tolerance parameters was conducted using the DIALLEL-SAS procedure developed by Zhang et al. (2005), according to Griffing`s (1956) Method II, Model I. The DIALLEL-SAS program evaluated main genotype effects contain GCA and SCA. For a diallel mating from a set of inbred lines, the generation means (Yhij) of maternal line i and paternal line j in block h of can be partitioned as:
Where:
| μ | = | The population mean |
| Gij | = | Genotypic effect |
| Bh | = | The block effect |
| ehij | = | Residual effect |
The following ratio was estimated to investigation of weight of additive or dominance effects (Baker, 1978):
The significance of the GCA of the parents and SCA, of their hybrids was calculated and the values of these components were estimated.
RESULTS
Analysis of variance (Table 1) indicated that there were highly significant differences among the replications, genotypes, General Combining Ability (GCA) and Specific Combining Ability (SCA) for all of traits contain percentage of reduction in radicle length (RL%), percentage of reduction in plumule length (PR%) and percentage of reduction in germination measure at percentage (GP%). The significant GCA and SCA observed in this study indicated the importance of both additive and non-additive gene actions in the inheritance of the traits (Table 1).
Reduction of RL, PR and GP were inversely proportional to cold tolerance, i.e., the higher the reduction in traits the lower were the cold evaluated and, among the parents, the highest reduction in RL, PR and GP according to low temperature was in the cold sensitive genotype Neda and the lowest reduction in these traits were in the genotypes contain Saleh, Hassani and Deilamani, respectively (Table 2-4). The F2 hybrids indicated values ranging from 96.83% (IRFAON215 X Shahpasand) to 11.47% (Neda X Hassani) for GP (Table 2), from 92.06% (IRFAON215 X Shahpasand) to 21.38% (Deilamani X Hassani) for PL and from 92.83% (IRFAON215 X Shahpasand) to 35.53% (Deilamani X Shahpasand) for Rl. In the crosses Hassani X Shahpasand, Sepidrod X Shahpasand, Neda X Shahpasand, Saleh X Shahpasand, Sepidrod X Hassani, Saleh X Sepidrod, Neda X Sepidrod, Deilamani X Neda and IRFAON215 X Saleh the F2 hybrids presented a lower mean for RL than the parents (Table 2). Also in the crosses Deilamani X Hassani, Sepidrod X Hassani, Neda X Hassani, RFAON215 X Hassani I, Saleh X Hassani, Saleh X Sepidrod, Neda X Sepidrod, Deilamani X Sepidrod and IRFAON215 X Saleh the F2 mean for PR was lower than that of the parents (Table 3). For GP, crosses of Sepidrod X Shahpasand, Neda X Hassani and Neda X Sepidrod too, the F2 mean was lower than that of the parents (Table 4). These results indicated over-dominance towards cold tolerance. On the other hand, was not found higher mean for any of traits in the crosses than parents and so, over-dominance towards cold sensitivity the F2 did not indicated for these traits. The value of average degree of dominance (2.22, 2.32 and 4.33 for RL, PL and GP, respectively) too indicated the presence of over-dominance for all of traits.
| Table 1: | Analysis of variance in a 7X7 complete diallel following Griffing` Method II Model I and estimation of genetic parameters |
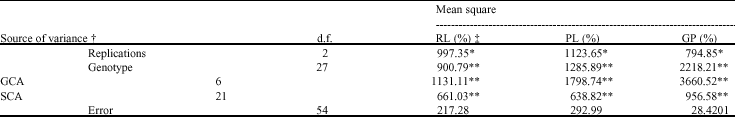 | |
| ** and * indicate significant at 1 and 5% levels. † GCA: General Combining Ability; SCA: Specific Combining Ability. ‡ RL: Radicle length reduction; PL: Plumule length reduction; GP: Germination percentage reduction | |
| Table 2: | Estimates of General Combining Ability (GCA), Specific Combining Ability (SCA) (above diagonal) of radicle length reduction and mean of them in F1 hybrids (below diagonal) and parents (diagonal) |
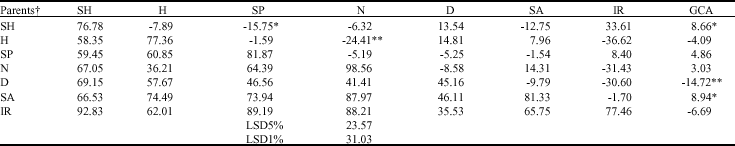 | |
| ** and * indicate significant at 1 and 5% levels, respectively. † SH: Shahpasand; H: Hassani; SP: Sepidrod; N: Neda; D: Deilmani; SA: Saleh; IR: IRFAON-215 | |
| Table 3: | Estimates of General Combining Ability (GCA), Specific Combining Ability (SCA) (above diagonal) of plumule length reduction and mean of them in F1 hybrids (below diagonal) and Parents (diagonal) |
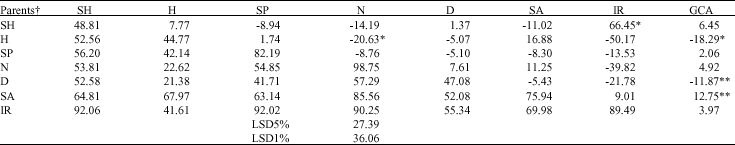 | |
| ** and * indicate significant at 1 and 5% levels, respectively. † SH: Shahpasand; H: Hassani; SP: Sepidrod; N: Neda; D: Deilmani; SA: Saleh; IR: IRFAON-215 | |
Negative values for the GCA and SCA effects revealed a contribution to cold tolerance at germination, or lower reduction in entire traits due to cold, while positive for the GCA and SCA effects indicate tendency towards cold sensitivity. For all of three traits the higher magnitude of the dominance variances than additive one and low value of genetic ratio (Table 5) showed that non-additive gene action was predominant to additive one.
Diallel analysis for RL revealed significant GCA and SCA mean squares, indicating that both additive and non-additive effects are involved (Table 2). The GCA effects were positively significant for Shahpasand and Saleh and negatively for Deilamani. A significant SCA effect was found for Sepidrod X Shahpasand and Neda X Hassani (Table 2). The results of diallel analysis for PL indicated significant GCA effects for Hassani and Deilamani (negatively) and Saleh (positively). For this trait too also showed negatively significance of SCA for Neda X Hassani and positively significance for IRFAON215 X Shahpasand (Table 3). The results of diallel analysis for GP indicated that GCA was significant in Shahpasand and IRFAON215 (positively) and in Hassani and Deilamani (negatively). SCA effects were negatively significant in the some of crosses such as Sepidrod X Shahpasand, Neda X Hassani and IRFAON-215 X Neda and positively for Saleh X Shahpasand, IRFAON215 X Shahpasand and IRFAON215 X Saleh (Table 4). Genetic variance estimates were superior to the environmental variance for all the crosses (Table 5) and consequently, nearly all of the crosses presented high values for broad sense heritability, the exception PL which presented moderate heritability. But all of the traits revealed low values of narrow sense heritability due to low value of additive effect.
| Table 4: | Estimates of General Combining Ability (GCA), Specific Combining Ability (SCA) (above diagonal) of germination percentage reduction and mean of them in F1 hybrids (below diagonal) and Parents (diagonal) |
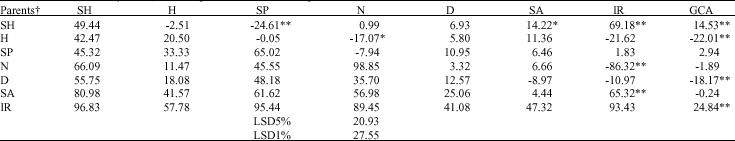 | |
| ** and * indicate significant at 1 and 5% levels, respectively. † SH: Shahpasand; H: Hassani; SP: Sepidrod; N: Neda; D: Deilmani; SA: Saleh; IR: IRFAON-215 | |
| Table 5: | Estimation of genetic parameters |
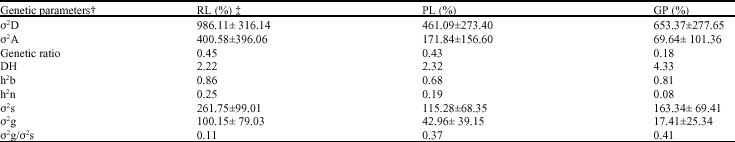 | |
| † σ2D: Dominance variance; σ2A: Additive variance; σ2s: SCA variance; σ2g: GCA variance; DH: Degree of dominance; h2n: Narrow-sense heritability; h2b: Broad-sense heritability. ‡ RL: Radicle length reduction; PL: Plumule length reduction; GP: Germination percentage reduction | |
DISCUSSION
Since most cold tolerance traits important for successful breeding are quantitative nature, the estimation of the general and specific combining ability, heritabilities and mode of inheritance contributes to their better understanding thereby enabling rational and targeted combining of desirable genes into future cultivars. Similarity of parents and progenies greatly depends on the knowledge of the relationship between the additive and non-additive components of variance and mitigates the selection of the investigated materials for creating hybrids or line cultivars.
Rice cold tolerance at the germination stage was evaluated using percentage of reduction in RL, PL and GP. These traits were chosen as they proved to be the most adequate ones to differentiate cold tolerant from cold sensitive genotypes at the germination stage (Cruz and Milach, 2004). Genetic effects were observed for all of three traits, indicating the greater importance of non-additive effects in comparison to additive effects in the crosses studied. In our present investigations, according to the diallel analysis, all of cold tolerance traits inherited over-dominantly in. According to the previous investigations, that discovered the inheritance of percentage of reduction in coleoptile and growth (Cruz et al., 2006) controlled by non-additive gene effects. In similar studies regarding rice cold tolerance at the vegetative and reproductive stages non-additive effects have also been reported to be more important than additive ones (Prianka et al., 2000). However, some previous investigations (Nishimura and Hamamura, 1993) of the inheritance of cold tolerance traits showed contradictory results, who have reported the predominance of additive gene action over dominance action for cold tolerance at the booting stage in japanica rice genotypes.
Broad sense heritability estimates based on variance of different generations were 86, 68 and 81%, for RL, PL and GP respectively (Table 5). The high heritability estimates indicated that cold tolerance is a heritable character. High heritability estimates for cold tolerance have been reported in rice (Prianka et al., 2000), who studied genetic variability in 26 rice genotypes to identify traits associated with cold tolerance and discovered high to moderate heritability for shoot: root ratio, cold tolerance, germination percentage, seedling vigor, survival percentage and chlorophyll content). Cold tolerant rice cultivars lose viability quickly when stored under ambient conditions. The report of Majumder (1999) indicated that there was increased unsaturation of polar lipids in cold tolerant. Thus, the seed deterioration in storage is associated with lipid per oxidation. .
Consequently, in this study the parents with negative GCA indicated that this parent is superior variety for cold tolerance, whereas can be distinguished inferior variety for cold tolerance due to negative GCA effects. In some of crosses significant negative SCA effect indicated, demonstrating the existence of heterosis for cold tolerance. Similarly, significance of the positive in the other crosses revealed heterosis for cold sensitivity at the germination stage. The ratio of estimated GCA to SCA variances indicated the preponderance of the latter component in controlling the expression of all the traits. The SCA variances involve both dominance and epistasis which together constitute the non additive type of gene action, whereas the GCA variance is a reflection of additive and additive X additive type of gene action. The non additive component of gene action could be optimized in upgrading the genetic potential of the crop by adopting reciprocal recurrent selection and hybrid technology, which has already become a field reality in rice. The perusal of GCA effects reveals that Deilamani was an ideal general combiner for all characters, whereas Hassani was a good general combiner for PL and GP showed negative significant GCA effects.
The estimates of SCA effects of top ranking crosses indicated that Neda X Hassani could be regarded as the most desirable cross combination for all of three traits. An important inference that can be drawn from these results is that cross combinations involving Hassani and Deilamani as one of the parents recorded desirable SCA effects for all or most of the traits studied.
REFERENCES
- Cruz, R.P. and S.C.K. Milach, 2004. Cold tolerance at the germination stage of rice: Methods of evaluation and characterization of genotypes. Sci. Agric., 61: 1-8.
Direct Link - Cruz, R.P., S.C.K. Milach and C.F. Luiz, 2006. Inheritance of rice cold tolerance at the germination stage. Genet. Molec. Biol., 29: 314-320.
Direct Link - Farrell, T.C., K.M. Fox, R.L. Williams and S. Fukai, 2006. Genotypic variation for cold tolerance during reproductive development in rice: Screening with cold air and cold water. Field Crops Res., 98: 178-194.
CrossRef - Griffing, B., 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci., 9: 463-493.
Direct Link - Han, L.Z., Y.Y. Zhang, Y.L. Qiao, G.L. Cao, S.Y. Zhang, J.H. Kim and H.J. Koh, 2006. Genetic and QTL analysis for low-temperature vigor of germination in rice. Acta Genet. Sinica, 33: 998-1006.
Direct Link - Kuroki, M., K. Saito, S. Matsuba, N. Yokogami, H. Shimizu and Y. Sato, 2007. A quantitative trait locus for cold tolerance at the booting stage on rice chromosome 8. Theor. Applied Genet., 115: 593-600.
CrossRef - Lou, Q., L. Chen, Z. Sun, Y. Xing, J. Li, X. Xu, H. Mei and L. Luo, 2007. A major QTL associated with cold tolerance at seedling stage in rice (Oryza sativa L.). Euphytica, 158: 87-94.
CrossRef - Majumder, M.K., 1999. Relationship between cold tolerance and ageing tolerance in rice. Indian Agric., 43: 61-166.
Direct Link - Murtaza, N., M. Kitaoka and G.M. Ali, 2005. Genetic differentiation of cotton cultivars by polyacrylamide gel electrophoresis. J. Centr. Eur. Agric., 6: 69-76.
Direct Link - Nishimura, M. and K. Hamamura, 1993. Diallel analysis of cool tolerance at the booting stage in rice varieties from Hokkaido. Jap. J. Breed., 43: 557-566.
Direct Link - Sthapit, B.R. and J.R. Witcombe, 1998. Inheritance of tolerance to chilling stress in rice during germination and plumule greening. Crop Sci., 38: 660-665.
Direct Link - Xu, L.M., L. Zhou, Y.W. Zeng, F.M. Wang, H.L. Zhang, S.Q. Shen and Z.C. Li, 2008. Identification and mapping of quantitative trait loci for cold tolerance at the booting stage in a japonica rice near-isogenic line. Plant Sci., 174: 340-347.
CrossRef - Zhang, Y., M.S. Kang and K.R. Lamkey, 2005. DIALLEL-SAS05: A comprehensive program for Griffing's and Gardner-Eberhart analysis. Agron. J., 97: 1097-1106.
CrossRefDirect Link