
J.K. Langat
Department of Plant Science and Crop Protection, University of Nairobi,P.O. Box 30197-00100, Nairobi, Kenya
J.W. Kimenju
Department of Plant Science and Crop Protection, University of Nairobi,P.O. Box 30197-00100, Nairobi, Kenya
G.K. Mutua
Department of Plant Science and Crop Protection, University of Nairobi,P.O. Box 30197-00100, Nairobi, Kenya
W.M. Muiru
Department of Plant Science and Crop Protection, University of Nairobi,P.O. Box 30197-00100, Nairobi, Kenya
W. Otieno
Kenya Plant Health Inspectorate Service, P.O. Box 49592-00100, Nairobi, Kenya
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 5 | Page No.: 467-472







ABSTRACT
This study was carried out with the aim of evaluating the effect of ecologically sound approaches for nematode management on non-target organisms, free-living nematodes. The materials tested were sugarcane bagasse, molasses, tea and flower composts, neem (Achook), a biological agent (Paecilomyces lilacinus) and fenamiphos (Nemacur). The treatments were administered before planting carnation var. White Natila in flower beds that were naturally infested with nematodes. Application of bagasse, molasses, tea and flower composts resulted in increased abundance of free-living nematodes compared to the control where nothing was applied. Bacterial feeders, fungal feeders, and predators comprised 73, 14 and 13%, respectively of the free-living nematodes recovered. Members of the genus Rhabditis were the most abundant (10%) among the bacteriovores while Mononchus (10%) and Aphelenchoides (14%) dominated among the predators and fungivorous trophic groups, respectively. The highest numbers of free-living nematodes were recorded at 90 Days after Planting (DAP) in plots treated with bagasse and molasses but the numbers declined at 180 DAP. A steady increase in numbers of free-living nematodes was observed in plots treated with tea and flower composts up to 180 DAP. Significant reductions in abundance of free-living nematodes were recorded in plots treated with fenamiphos and neem. This study has established that application of organic substrates serve as a stimulus to processes leading to build-up of free-living nematodes. The organic substrates can strongly be recommended for use in sustainable carnation production systems.
PDF Abstract XML References Citation
How to cite this article
J.K. Langat, J.W. Kimenju, G.K. Mutua, W.M. Muiru and W. Otieno, 2008. Response of Free-Living Nematodes to Treatments Targeting Plant Parasitic Nematodes in Carnation. Asian Journal of Plant Sciences, 7: 467-472.
DOI: 10.3923/ajps.2008.467.472
URL: https://scialert.net/abstract/?doi=ajps.2008.467.472
DOI: 10.3923/ajps.2008.467.472
URL: https://scialert.net/abstract/?doi=ajps.2008.467.472
INTRODUCTION
The horticulture sub-sector has registered a phenomenal growth of 15-20% in the past decade to rank third as a foreign exchange earner in Kenya. It is currently the fastest growing agricultural sub-sector, with annual earnings of US$ 300 million (Waceke et al., 2005). Fruits, vegetables and cut-flower production are the main sectors of horticultural production in Kenya. The cut flower industry dominates the horticultural exports, earning US$ 500 million in 2003 and provides direct employment to over 500,000 people (Manda and Sen, 2005).
Carnation (Dianthus caryophyllus L.) is grown on more than 500 ha mainly under greenhouse conditions. Production of the crop has been increasing steadily in response to the expanding market demand (Waceke et al., 2001). Ultimately, Kenya has become the European Union`s biggest source of the carnations, having overtaken Israel as market leader. Nematodes have been rated among the principal constraints to carnation production since they damage roots and reduce the ability of plants to take up water or nutrients from soil (Masse et al., 2002). In ornamentals, the problem is compounded by foliar nematodes (Aphelenchus spp., Aphelenchoides spp. and Ditylenchus spp.) which cause qualitative losses. Losses due to nematodes in cut flower production are estimated to be 10-20% worldwide (Agudelo, et al., 2006). Chemical nematicides have dominated in the management of the nematodes but environmental concerns have provided an added impetus to the search for alternative strategies (Wang, 2004). Biological control, combined with other feasible strategies is an area that is fast gaining popularity as a sustainable strategy in nematode management (Mostafa, 2001; Kiewnick and Sikora, 2004). This approach optimizes ecological synergies between biological components of the ecosystem, enhancing biological efficiency of soil processes in order to maintain soil fertility, productivity and crop protection in a new approach referred to as ecologically based pest management, which aims at minimizing adverse effects on non-target species and the environment (Steinberger et al., 2001). While plant parasitic nematodes have a negative impact on the plant, free living nematodes are known to play a role in nutrient cycling among other benefits. According to Sánchez-Moreno and Navas (2007), abundance and diversity of free-living nematodes is a good indicator of soil health, given their role in decomposition and regulation of bacterial and fungal microbes (Al-Sayed et al., 2007). It is, therefore, imperative to conserve biodiversity of these free-living nematodes. Whilst large data sets are available on the effect of organic substrates on plant parasitic nematodes, few investigations have examined their effects on the free-living nematode fauna (Ekschmitt et al., 2001). This study was undertaken with the aim of determining the effects of treatments targeting plant parasitic nematodes on free-living nematodes.
MATERIALS AND METHODS
The study was carried out between June 2006 and July 2007 at Kericho, Kenya. The experiments were conducted under greenhouse conditions in an area situated at an altitude of 1950 m a.s.l and receives mean annual rainfall of 2000 mm with mean annual day and night temperatures of 25 and 11°C, respectively. The soil type is sandy loam humic nitisols with an inherent pH of 4-4.5 but it is usually amended using agricultural lime to pH 6 to make it suitable for growing of carnations.
Parallel flower beds, measuring 100 cm in width and raised to a height of 25 cm, were made in a greenhouse, leaving a path of 50 cm between them. Experimental plots, measuring 100x400 cm, were marked out in the flower beds. The treatments tested were bagasse, tea and flower composts, molasses (a by-product of sugarcane processing), neem, Paecilomyces lilacinus (PL plus) and fenamiphos (Nemacur) while untreated plots were included as a control. Bagasse, tea and flower composts were sun dried to a constant weight. The composts and bagasse were applied at the rate of 300 t ha-1 as recommended for greenhouse usage (McSorley and Gallagher, 1995). Molasses was applied at the rate of 667 mL m-3 (Schenck, 2001) while neem was applied following the manufacturer`s recommendation at 1.5 mL m-2 dissolved in 1000 L of water.
Paecilomyces lilacinus (PL-plus) was applied at the rate of 2 kg ha-1 while fenamiphos was applied 1 week before planting at the rate of 30 g m-2. Spray carnations cv. White Natila seedlings were transplanted into the plots. Treatments were arranged in a completely randomized design with 6 replications.
Characterization of nematodes in the research site: Five soil sub-samples were randomly collected to a depth 30 cm from the middle rows of each plot. The nematodes were extracted from 200 cm3 soil using the procedure described by Hooper et al. (2005). The sub-samples were thoroughly mixed to form a composite sample before being placed in plastic sampling bags, transported to the laboratory and stored at 4°C. Soil sampling was done at planting, 90 and 180 days after planting. Nematodes from each sample were fixed using rapid Seinhorst technique and thereafter mounted on Cobb-type aluminium double cover glass slides that allow examination from either side (Siddiqi, 2000). Identification of the nematodes was based on morphological characteristics and pictorial keys using a high power microscope (Hunt et al., 2005). After identification, the nematodes were assigned to trophic groups as described by Yeates et al. (1993). Nematode abundance was transformed to Log (x+1) and different treatments were compared by two-way Analysis of Variance (ANOVA). Comparison of the means was done with the Least Significant Difference (LSD) test.
RESULTS
Effect of organic substrates and bio-control agents on non-parasitic nematodes in carnations: Application of bagasse, molasses, tea and flower composts as organic amendments led to significant (p≤0.05) increase in numbers of free-living nematodes in the soil (Table 1). A decline in nematode numbers was recorded in plots treated with fenamiphos and neem, compared to the control. The differences in mean numbers of free-living nematodes were not significant in plots that were treated with P. lilacinus, compared to the control.
Nematodes from 14 genera were recovered from carnation plots treated with organic substrates and the bio-control agent, P. lilacinus (Table 2). Among the free-living nematodes, bacterial feeders accounted for 73% while fungal feeders and predators accounted for 14 and 13%, respectively. The bacterivorous nematodes were members of the genera Acrobeles, Rhabditis, Cephalobus, Prodorylaimus, Bunonema, Eucephalobus, Heterocephalobus, Plectus, Nygolaimus and Chromadora. The predators were members of the genera Mononchus and Labronema while the fungivores were assigned to the genera Aphelenchoides and Aphelenchus. Members of the genus Rhabditis were predominant among the bacterial feeding nematodes, representing 10% of the nematodes while Mononchus (10%) and Aphelenchoides (14%) dominated the predacious and fungivorous trophic groups, respectively. The treatments had a significant effect on the free-living nematodes, with the exception of those in the genera Cephalobus and Chromadora.
| Table 1: | Effect of different treatments on the free-living nematodes in carnation production systems |
 | |
| *Mean followed by different letter(s) are significantly different along the row (p≤0.05) | |
| Table 2: | Occurrence and abundance of free living nematodes following application of the various organic substrates in carnations |
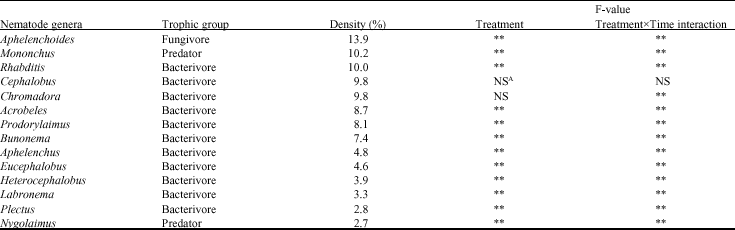 | |
| ANS: Not Significant, **Significant differences | |
| Table 3: | Effect of different treatments on free-living nematodes after varying durations from planting to 180 days after planting |
 | |
| *Means followed by different letter(s) are significantly different along the rows | |
| Table 4: | Effect of different treatments, applied to control parasitic nematodes, on free living nematodes in the soil |
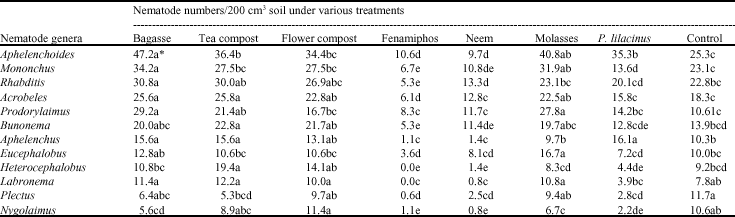 | |
| *Means within the same column followed by different letter(s) are significantly different (p≤0.05) | |
The treatments had variable effects on numbers of free-living nematodes over time, from application during planting throughout to 180 days after planting (Table 3). With the exception of fenamiphos and neem, all the other treatments led to a sharp increase in numbers of free-living nematodes within the first 90 DAP. The nematode numbers continued to increase in plots treated with tea and flower composts, fenamiphos as well as in the control up to 180 days.
Free-living nematodes from different genera had varying responses to the various treatments aimed at controlling plant-parasitic nematodes (Table 4). The organic amendments namely bagasse, molasses, tea and flower composts induced an increase in numbers of nematodes from most of the genera of free-living nematodes. Bagasse led to an increase in all the nematodes, except Nygolaimus and Eucephalobus spp. While most organic substrates had a positive effect on the populations of bacterivorous nematodes, P. lilacinus (biocontrol agent) greatly increased the populations of fungivorous nematodes in the genera Aphelenchus spp. and Aphelenchoides spp. Plots treated with fenamiphos and neem had significantly lower numbers of free living nematodes compared to the other treatments.
DISCUSSION
This study has demonstrated that amending soils with organic substrates as well as with biological agents contribute to a change in nematode community structure by increasing the abundance of free-living nematode populations. Populations of free-living nematodes have been shown to increase rapidly following the addition of organic substrates (Akhtar and Malik, 2000; Agyarko and Asante, 2005). The mode of action of organic substrates leading to stimulation of free-living nematodes is complex and dependent on the nature of the substrate (El-Sherif et al., 2007). These mechanisms stem from the decomposition process that leads to changes in the physical and chemical properties of the soil. According to Sanchez and Navas (2007), the nematode community structure is strongly impacted by changes in soil systems since nematodes are highly dependent on soil properties. When incorporated into the soil, organic substrates undergo a series of processes that release NH4+, formaldehyde, phenols and volatile fatty acids, among other compounds (Wang et al., 2004). The compounds may act individually or collectively to stimulate build-up of beneficial microbes including free-living nematodes (Desaeger and Rao, 2002). According to Akhtar and Malik (2000), there could be a correlation between increase in NH4+ and an increase in numbers of free-living nematodes following addition of organic substrates. In addition, free-living nematodes may accelerate the decomposition of soil organic matter and increase mineralization of nitrogen and phosphorous thus triggering a chain reaction that favours increase of the nematodes (Kimenju et al., 2004).
From this study, the diversity of free-living nematodes recovered from the carnation production system was lower compared to previous studies in other production systems (Yeates et al., 1999; Zolda, 2006). This can be attributed to the fact that cut-flower production is characterized by usage of enormous amounts of agrochemicals, mainly in the form of fertilizers and pesticides (Tenenbaum, 2002). Loss of biodiversity has indeed been attributed to the adverse effects that are associated with agrochemicals, especially the use of nematicides (Yeates et al., 1999). Fumigation of soil to control soilborne pathogens and nematodes is recognized as one of the most serious threats to the beneficial organisms such as free-living nematodes (Pinkerton et al., 2000). Bacterial feeders dominated the trophic groups isolated in this study and this finding is consistent with reports from other workers (Wasilewska, 1997; Ekschmitt et al., 2001). According to Zolda (2006), the elevated numbers of bacterial feeding nematodes can be attributed to increased food resources for the microorganisms in the soil. Indeed, bacterivorous nematodes responded quickly to increased food supply (Yeates et al., 1999). In addition, decomposition pathways in agricultural systems are mostly driven by bacteria which serve as a stimulus to increased numbers of bacterial feeding nematodes (McSorley and Frederick, 2000). Rapid changes in numbers of bacterivorous nematodes can be anticipated, given their short generation time of 3-4 days (Ruess, 2003).
The dominance of fungal feeding genera (Aphelenchoides spp. and Aphelenchus spp.) in soils amended with P. lilacinus (PL Plus) is noteworthy because fungivorous genera are normally found at lower densities than bacteriovores and predators (Knudsen and Bae, 2001). Paecilomyces lilacinus is an ubiquitious soil hyphomycete which parasitizes eggs of root-knot nematodes thus regulating populations of the nematodes in field soil (Schenck, 2004). Numerous species of fungivores have been found in soil with the most common genera being Aphelenchus and Aphelenchoides (Yeates et al., 1999). A fungal-dominated decomposition pathway is however likely, considering that species of one of the most prevalent genus in this study, namely Acrobeles as well as plant parasitic Filenchus and Tylenchus may also feed on fungi (Zolda, 2006). The relative increase in fungal biomass occasioned by application of P. lilacinus, could have resulted in the relative increase of fungal feeding nematodes. Kiewnick and Sikora (1989) reported that numbers of the mycophagous nematodes, Aphelenchoides sp. and Aphelenchus avenae, increased several-fold within a few days after adding flax to roots that had been precolonized by Rhizoctonia solani. Moreover, abundance of bacterivorous nematodes has been shown to reduce bacterial biomass, occasioning a relative increase of fungal biomass (Soylu et al., 2005). Bae et al. (2001) observed many nematodes associated with Trichoderma hyphae presumably feeding and numerous nematode eggs adhering to hyphae. This led to the conclusion that populations of fungivorous nematodes may increase rapidly following addition of fungi as biocontrol agents.
The study revealed that neem and fenamiphos caused a reduction in numbers of free-living nematodes. Pesticidal properties of azadirachtin, the active ingredient in neem, have been clearly documented (Akhtar and Malik, 2000; Agyarko and Asante, 2005). According to Agyarko and Asante (2005), neem based products reduced egg hatch and the mobility of nematode juveniles. Apart from azadirachtin, several other compounds namely salannin, nimbidin, thionemone ammonia, phenol, formaldehyde and fatty acids are released during decomposition of neem-based products (Yasmin et al., 2003). It is possible that these compounds are individually or collectively detrimental to free-living nematodes, thus accounting for the decline in their numbers in treated plots. In conclusion, organic substrates are increasingly gaining popularity as components of integrated pest management, developed with the goal of reducing chemical usage in the control of plant parasitic nematodes. The fact that addition of the organic substrates results in build-up of beneficial organisms such as free-living nematodes is an added advantage.
CONCLUSION
An increase in numbers of free-living nematodes was recorded in all plots where different organic substrates were applied with exception of neem. A decline in numbers of the nematodes was observed in plots treated with chemical nematicides (fenamiphos). It is recommended, the organic amendments should be incorporated in carnation production given the range of other benefits which include disease suppression, increased soil water holding capacity, improvement of soil fertility and structure.
ACKNOWLEDGMENTS
The authors are grateful to James Finlay (K) Ltd. for providing financial and material support towards this study and the University of Nairobi for providing laboratory space and equipment.
REFERENCES
- Agudelo, P.A., S.A. Lewis and M.A. Abril, 2006. First report of root-knot nematode Meloidogyne javanica on chrysanthemum in Colombia. Plant Dis., 90: 828-828.
Direct Link - Akhtar, M. and A. Malik, 2000. Roles of organic soil amendments and soil organisms in the biological control of plant-parasitic nematodes: A review. Bioresour. Technol., 74: 35-47.
CrossRefDirect Link - Al-Sayed, A.A., A.M. Kheir, H.I. El-Naggar and H.H. Kesba, 2007. Organic management of Meloidogyne incognita on grapes in relation to host biochemistry. Int. J. Agric. Res., 2: 776-785.
CrossRefDirect Link - Agyarko, K. and J.S. Asante, 2005. Nematode dynamics in soil amended with neem leaves and Poultry manure. Asian J. Plant Sci., 4: 426-428.
Direct Link - Bae, Y.S., G.R. Knudsen and L.M.C. Dandurand, 2002. Influence of soil microbial biomass on growth and biocontrol efficacy of Trichoderma harzianum. Plant Pathol. J., 18: 30-35.
Direct Link - Desaeger, J. and M.R. Rao, 2000. The root-knot nematode (Meloidogyne spp.) problem in Sesbania fallows and the scope for management in Western Kenya. Agrof. Syst., 47: 273-288.
Direct Link - El-Sherif, A.G., A.R. Refaei, M.E. El-Nagar and H.M.M. Salem, 2007. Integrated Management of Meloidogyne incognita infecting eggplant by certain organic amendments, Bacillus thuringiensis and oxamyl with reference to N P K and Total chlorophyll status. Plant Pathol. J., 6: 147-152.
CrossRefDirect Link - Ekschmitt, K., G. Bakonyi, M. Bongers, T. Bongers and S. Bostro¨m et al., 2001. Nematode community structure as indicator of soil functioning in European grassland soils. Eur. J. Soil Biol., 37: 263-268.
CrossRef - Hooper, D.J., J. Hallmann and S.A. Subbotin, 2005. Methods for Extraction, Processing and Detection of Plant and Soil Nematodes. In: Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, Luc, M., R.A. Sikora and J. Bridge (Eds.)., 2nd Edn., CAB International Publisher, UK, ISBN: 9781845931445, pp: 53-86.
Direct Link - Hunt, D.J., M. Luc and R.H. Manzanilla-López, 2005. Identification, Morphology and Biology of Plant Parasitic Nematodes. In: Plant Parasitic Nematodes in Subtropical and Tropical Agriculture, Luc, M., R.A. Sikora and J. Bridge (Eds.), CABI International, Wallingford, England, ISBN-13: 978-1-84593-144-5, pp: 11-52.
CrossRefDirect Link - Kiewnick, S, and R. A Sikora, 2004. Optimizing the efficacy of Paecilomyces lilacinus (strain 251) for the control of root-knot nematodes. Commun. Agric. Applied Biol. Sci., 69: 373-380.
PubMedDirect Link - Kimenju, J.W., D.M. Muiru, N.K. Karanja, M.W. Nyongesa and D.W. Miano et al., 2004. Assessing the role of organic amendments in management of root-knot nematodes on common bean, Phaseolus vulgaris L. Trop. Microbiol. Biotechnol., 3: 14-23.
Direct Link - Manda, D.K. and K. Sen, 2004. The labour market effects of globalization in Kenya. J. Int. Dev., 16: 29-43.
CrossRef - Masse, D., E. Pate, N. Ndiaye-Faye and P. Cadet, 2002. Effect of fallow improvement on the nematode community in the Sudanian region of Senegal. Eur. J. Soil Biol., 38: 205-211.
CrossRef - Mostafa, F.A.M., 2001. Integrated control of root-knot nematodes, Meloidogyne spp. Infecting sunflower and tomato. Pak. J. Biol. Sci., 4: 44-46.
CrossRefDirect Link - Wang, K.H., R. McSorley and N. Kokalis-Burelle, 2006. Effects of cover cropping, solarization and soil fumingation on nematode communities. Plant Soil, 286: 229-243.
CrossRef - Ruess, L., 2003. Nematode soil faunal analysis of decomposition pathways in different ecosystems. J. Nematol., 5: 179-181.
CrossRef - Sa´nchez-Moreno, S. and A. Navas, 2007. Nematode diversity and food web condition in heavy metal polluted soils in a river basin in southern Spain. Eur. J. Soil Biol., 43: 166-179.
CrossRef - Soylu, S., E.M. Soylu, S. Kurt and O.K. Ekici, 2005. Antagonistic potentials of rhizosphere-associated bacterial isolates against soil-borne diseases of tomato and pepper caused by Sclerotinia sclerotiorum and Rhizoctonia solani. Pak. J. Biol. Sci., 8: 43-48.
CrossRefDirect Link - Steinberger, Y., W. Liang, E. Savkina, T. Meshi and G. Barness, 2001. Nematode community composition and diversity associated with a topoclimatic transect in a rain shadow desert. Eur. J. Soil Biol., 37: 315-320.
CrossRef - Tenenbaum, D., 2002. Would a rose not smell as sweet?: Problems stem from the cut flower industry. Environ. Health Perspect., 110: A240-A247.
Direct Link - Waceke, J.W., S.W. Waudo and R. Sikora, 2001. Response of Meloidogyne hapla to mycorrhiza fungi inoculation on pyrethrum. Afr. J. Sci. Technol., 2: 63-70.
Direct Link - Yasmin, L., M.H. Rashid, M.N. Uddin, M.S. Hossain, M.E. Hossain and M.U. Ahmed, 2003. Use of neem extract in controlling root-knot nematode (Meloidogyne javanica) of sweet-gourd. Plant Pathol. J., 2: 161-168.
Direct Link - Yeates, G.W., 2003. Nemtodes as soil indicators: Functional and biodiversity aspects. Biol Fert. Soils, 37: 199-210.
CrossRef - Yeates, G.W. and T. Bongers, 1999. Nematode diversity in agroecosystems. Agric. Ecosyst. Environ., 74: 113-135.
CrossRefDirect Link - Zolda, P., 2006. Nematode communities of grazed and ungrazed semi-natural steppe grasslands in Eastern Austria. Pedobiol., 50: 11-22.
CrossRef