
Solomon Admassu
Ethiopian Institute of Agricultural Research,Awassa National Maize Research Project, Awassa, Ethiopia
Mandefro Nigussie
Ethiopian Institute of Agricultural Research, Melkasa Research Center, Nazreth, Ethiopia
Habtamu Zelleke
Department of Plant Science, Faculty of Agriculture, University of Haramaya, Dire Dawa, Ethiopia
Asian Journal of Plant Sciences
Year: 2008 | Volume: 7 | Issue: 2 | Page No.: 163-169







ABSTRACT
Fifteen maize genotypes were tested at nine different locations in 2005 under rainfed condition to determine stable maize genotypes for grain yield and determine genotypes with high yield and form homogenous grouping of environments and genotypes. The experiment was conducted using Randomized Complete Block Design with three replications. There was considerable variation among genotypes and environments for grain yield. Stability was estimated using the Additive Main Effects and Multiplicative Interactions (AMMI). Based on the stability analysis, genotypes 30H83, BH-540, Ambo Synth-1, AMH-800 and BHQP-543 were found to be stable for grain yield. The first two Interaction Principal Component axis (IPCA1 and IPCA2) were significant (p<0.01) and cumulatively contributed 70.27% of the total genotype by environment interaction. The coefficient of determination (R2) for genotypes 30H83 was as high as 0.92, confirming its high predictability to stability. Among the genotypes, the highest grain yield was obtained from genotype 30H83 and BH-541 (8.98 and 8.05 t ha-1) across environments. Clustering of AMMI-estimate values grouped genotypes in to four clusters and the environment in to three clusters. Environment Goffa was unique as it is grouped differently from all other environments.
PDF Abstract XML References Citation
How to cite this article
Solomon Admassu, Mandefro Nigussie and Habtamu Zelleke, 2008. Genotype-Environment Interaction and Stability Analysis for Grain Yield of Maize (Zea mays L.) in Ethiopia. Asian Journal of Plant Sciences, 7: 163-169.
DOI: 10.3923/ajps.2008.163.169
URL: https://scialert.net/abstract/?doi=ajps.2008.163.169
DOI: 10.3923/ajps.2008.163.169
URL: https://scialert.net/abstract/?doi=ajps.2008.163.169
INTRODUCTION
Information about phenotypic stability is useful for the selection of crop varieties and breeding programs. Plant breeders invariably encounter GenotypexEnvironment Interactions (GEI) when testing varieties across a number of environments. Depending up on the magnitude of the interactions or the differential genotypic responses to environments, the varietal ranking can differ greatly across environments (Kaya et al., 2002). The phenotypic performance of a genotype is not necessarily the same under diverse agro-ecological conditions. The concept of stability has been defined in several ways and several biometrical methods including univariate and multivariate ones have been developed to assess stability (Crossa, 1990; Ngeve and Bouwkamp, 1993; Sneller et al., 1997; Scapim et al., 2000). A combined analysis of variance can quantify the interactions and describe the main effects. However, analysis of variance is uninformative for explaining GEI. Other statistical models for describing GEI such as the Additive Main Effects and Multiplicative Interaction (AMMI) model are useful for understanding GEI.
The AMMI model is a hybrid that involves both additive and multiplicative components of the two-way data structure. AMMI biplot analysis is considered to be an effective tool to diagnose GEI patterns graphically. In AMMI, the additive portion is separated from interaction by Analysis of Variance (ANOVA). Then the Principal Component Analysis (PCA), which provides a multiplicative model, is applied to analyze the interaction effect from the additive ANOVA model. The biplot display of PCA scores plotted against each other provides visual inspection and interpretation of the GEI components. Integrating biplot display and genotypic stability statistics enables genotypes to be grouped based on similarity of performance across diverse environments (Thillainathan and Fernandez, 2001).
The AMMI model combines the analysis of variance for the genotype and environment main effects with principal components analysis of the genotype environment interaction (Kaya et al., 2002). The results can be graphed in a useful biplot that shows both main and interaction effects for both genotypes and environments (Annicchiarico, 2002). AMMI combines Analysis of Variance (ANOVA) into a single model with additive and multiplicative parameters.
AMMI is ordinarily the model of first choice when main effects and interaction effects are both important, which is a case in most yield trials (Mandel, 1971). If, for example, only main effects (additive structure) are present in the data, then the AMMI can be reduced to an ANOVA model. Whereas, if non-additive structure is only present then the PCA model is reflected. AMMI results can be readily used to diagnose these and other sub cases (Gabriel, 1978). The pattern portion of GEI sum of squares captured by the regression approach (heterogeneity among regressions) can at best capture only the amount of GEI sum of squares modeled by the simplest AMMI model. Therefore, AMMI analysis can potentially glean more patterns from the GEI than the regression approach (Sneller et al., 1997). In addition, the AMMI analysis can be applied to data sets where regression analysis may be inappropriate.
The combination of analysis of variance and principal components analysis in the AMMI model, along with prediction assessment, is a valuable approach for understanding GEI and obtaining better yield estimates. Therefore, the objectives of this study were to estimate Genotype-Environment (GE) interactions, to determine stable maize genotypes for grain yield and to determine genotypes with high yields, depending on the differential genotypic responses to environments and to form homogenous grouping of environments and genotypes.
MATERIALS AND METHODS
Fifteen maize genotypes were evaluated at nine locations in 2005 crop under rainfed condition. Randomized Complete Block Design (RCBD) with three replications was used. Each plot had four rows of 5.1 m length with spacing of 75 cm between rows and 30 cm between plants. Two seeds were planted per hill and then thinned to one plant per hill to have a final plant density of about 44,444 plants ha-1. To reduce border effects, data were recorded from the two central rows of each plot. Other management practices were done as recommended for each location. Fifteen maize genotypes of diverse origin were included in the study. The genotypes include top crosses, single crosses, three-way crosses and synthetics (Table 1).
The locations where the experiment was conducted were different in soil type, altitude and mean annual rainfall and considered as individual environment (Table 2). Several traits were assessed but only data for grain yield (t ha-1, at 12.5% grain moisture, estimated on the basis of two row plot) is reported here.
Analysis of variance for each environment was done for grain yield and other traits, using the SAS computer program (SAS, 2001). Bartlett`s test was used to assess homogeneity of error variances prior to combine analysis over environments. Genotypexenvironment interaction was quantified using the most common procedure; i.e., pooled analysis of variance, which partitions the total variance into its components (genotype, environment, genotypexenvironment interaction and pooled error). Environments were considered as random factors while the effect of genotypes was regarded as fixed. AMMI was used to test the stability of genotypes.
| Table 1: | Description of maize genotypes used for the study |
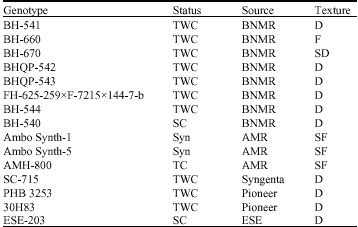 | |
| SF: Semi Flint; D: Dent; SD: Semi-Dent; F: Flint; TWC: Three-Way Cross; SC: Single Cross hybrid; TC: Top Cross; Syn: Synthetic; ESE: Ethiopian Seed Enterprise; QPM: Quality Protein Maize; BNMR: Bako National Maize Research and AMR: Ambo Maize Research | |
| Table 2: | Description of the test locations |
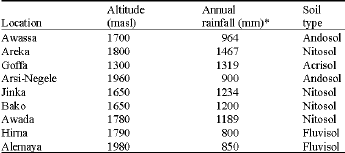 | |
| *: Mean of 10 years | |
RESULTS AND DISCUSSION
The AMMI analysis of variance for grain yield of 15 maize genotypes evaluated at nine locations showed that the total SS 31.3% was attributed to environmental effects for genotypic effects 24.2 and 22.8% was due to genotypexsignificant MS of environment indicated that the environments were diverse, with large differences among environmental means causing most of the variation in grain yield, which is in harmony with the findings by Taye et al. (2000), Kaya et al. (2002) and Alberts (2004). This indicated that the overwhelming influence that environments have on the performance of maize genotypes. Sneller et al. (1997), Tiruneh (1999), Taye et al. (2000) , Abush (2001) Kaya et al. (2002) and Alberts (2004) also reported similar results whose all the genotypes, environmental and genotypexenvironment effects were declared significant in the ANOVA of AMMI. Mean grain yield of the maize genotypes varied among environments ranging from 5.99 t ha-1 for environment Jinka to 8.98 t ha-1 for environment Bako. The mean grain yield of the 15 genotypes ranged from 5.63 to 8.98 t ha-1 and the highest grain yield was obtained from genotype 30H83 and BH-541 (Table 4).
| Table 3: | Additive main effects and multiplicative interaction analysis of variance for grain yield of genotypes across environments |
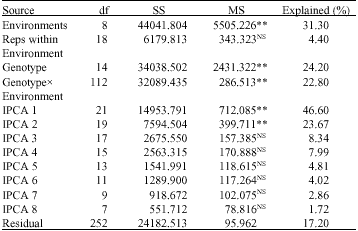 | |
| NS: Non-Significant, **: Significant at p<=0.01 level, respectively, Grand mean = 7.25 t ha-1, R2 = 0.8279, CV = 13.5% | |
| Table 4: | IPCA1, IPCA2, R2 and grain yield t ha-1 for 15 genotypes |
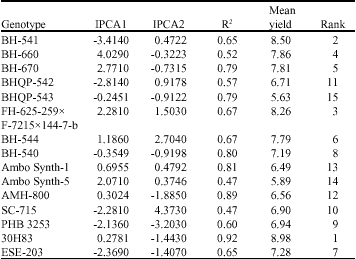 | |
Results from AMMI analysis (Table 3) also showed that the first Interaction Principal Component Axis (IPCA1) of the interaction captured 46.6% of the interaction sum of squares. Similarly, the second Interaction Principal Component Axis (IPCA2) explained a further 23.67% of the GEI sum of squares. The mean square for IPCA1 and IPCA2 were significant at p<=0.01 and cumulatively contributed to 70.27% of the total GEI. The F-test at p<=0.01 suggested that the two principal component axes of the interaction were significant for the model with 40°C of freedom. Hence, the AMMI with only two interaction principal component axes was the best predictive model, which is in agreement with Zobel et al. (1988) and Annicchiarico (2002). Further interaction principal component axes captured mostly noise and therefore did not help to predict validation of observations. Thus, the interaction of the 15 genotypes with nine environments was best predicted by the first two principal components of genotypes and environments.
Most accurate model for AMMI can be predicted by using the first two PCAs (Gauch and Zobel, 1996; Yan et al., 2000; Annicchiarico, 2002). Conversely, Sivapalan et al. (2000) recommended a predictive AMMI model with the first four PCAs. These results indicate that the number of the terms to be included in an AMMI model cannot be specified a priori without first trying AMMI predictive assessment.
By plotting both the genotypes and the environments on the same graph, the associations between the genotypes and the environments can be seen clearly (Fig. 1). The IPCA scores of genotypes in the AMMI analysis are an indication of the stability or adaptation over environments (Gauch and Zobel, 1996; Purchase, 1997; Alberts, 2004). The greater the IPCA scores, the more specific adapted is a genotype to certain environments. The more the IPCA scores approximate to zero, the more stable or adapted the genotype is over all the environments sampled.
A biplot is generated using genotypic and environmental scores of the first two AMMI components (Vergas et al., 1999). A biplot has four sections, depending upon signs of the genotypic and environmental scores. In Fig. 2, the sites fell into four sections: the best genotype with respect to site Alemaya, Awada and Arsi-Negelle were genotypes BH-544 and FH-625-259 xF-7215x144-7-b: Genotypes BH-660, BH-670 and AMH-800 were best for sites Bako and Areka on the other hand the best genotypes for Hirna, Awassa and Jinka were BH-541, BHQP-542 and SC-715, while Genotype PHB-3253 and ESE-203 were best for site Goffa.
Estimation of environmental indices (Ij) were used to classify environments into three classes viz., positive significant as good (favorable environments), positive or negative non-significant as average environments and negatively significant as poor (unfavorable) environments (Table 6). Based on the results of the analysis Bako, Awassa and Hirna were favorable environments with environmental index of positive and significant. Awada, Jinka and Goffa were poor (unfavorable) environments with negative and significant environmental index, while Alemaya and Arsi-Negelle were average environments.
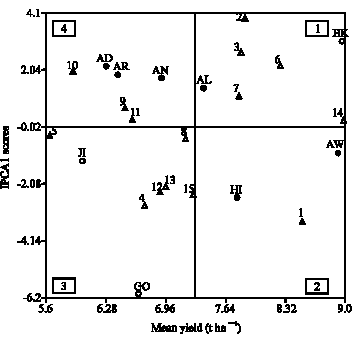 | |
| Fig. 1: | AMMI1 biplot of main effects and interactions. Where, 1 = BH-541; 2 = BH-660; 3 = BH-670; 4 = BHQP-542; 5 = BHQP-543; 6 = FH-625-259xF-7215 x144-7-b; 7 = BH-544; 8 = BH-540; 9 = Ambo Synth-1; 10 = Ambo Synth-5; 11 = AMH-800; 12 = SC-715; 13 = PHB 3253; 14 = 30H83, 15 = ESE-203; AW = Awassa; AR = Areka; GO = Goffa; AN = Arsi-Negelle; JI = Jinka; BK = Bako; AD = Awada; HI = Hirna and AL = Alemaya |
Table 4 and 5 shows the AMMI analysis with the IPCA1 and IPCA2 scores for the genotypes and environments, respectively. When looking at the environments it was clear that there is a good variation in the different environments sampled, ranging from the lower yielding environments in quadrants 3 and 4 and the high yielding environments in quadrants 1 and 2. With respect to the test sites, Goffa, was most discriminating as indicated by the longest distance between its marker and the origin (Fig. 2). However, due to its large PCA2 score, genotypic differences observed at Goffa may not exactly reflect the genotypes in average yield overall sites. Site Awassa was not the most discriminating, but genotypic differences at Awassa should be highly consistent with those averaged over sites, because it had near zero PCA2 scores compared to the other.
Awassa and Bako were the most favorable environments for all genotypes with nearly similar yield response but slight difference in interaction (Fig. 1). Jinka and Awada were the least favorable environments for all genotypes, with different interaction and different yield response.
The genotypes had considerably less variation than the environments around the mean yield of 7.25 t ha-1. The genotypes 30H83, BH-541 and FH-625-259x F-7215x 144-7-b are mainly adapted to higher yielding environments. Considering only the IPCA1 scores, genotypes BH-660, BH-541 and BH-670 was unstable genotypes and also adapted to the higher yielding or more favorable environments. Genotypes adapted to lower yielding environments were BH-540, AMH-800 and Ambo Synth-1. The most stable genotypes based on IPCA1 scores, were BH-540, 30H83, AMH-800, Ambo Synth-1 and BHQP-543.
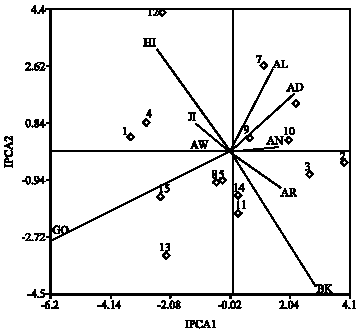 | |
| Fig 2: | Interaction biplot for the AMMI2 model. Where, 1 = BH-541; 2 = BH-660; 3 = BH-670; 4 = BHQP-542; 5 = BHQP-543; 6 = FH-625-259xF-7215x144-7-b; 7 = BH-544; 8 = BH-540; 9 = Ambo Synth-1; 10 = Ambo Synth-5; 11 = AMH-800; 12 = SC-715; 13 = PHB 3253; 14 = 30H83, 15 = ESE-203; AW = Awassa; AR = Areka; GO = Goffa; AN = Arsi-Negelle; JI = Jinka; BK = Bako; AD = Awada; HI = Hirna and AL = Alemaya |
Since IPCA2 scores also play a significant role (23.67%) in explaining the GEI, the IPCA1 scores were plotted against IPCA2 scores to further explore adaptation (Fig. 2). SC-715 was the most unstable in addition, PHB-3253, BH-541, BH-660, BH-544 and BHQP-542 were unstable to a lesser extent. BH-540, BHQP-543, 30H83, AMH-800 and Ambo Synth-1 were stable, when plotted on the IPCA1 and IPCA2 scores.
Adaptation of the genotypes based on the AMMI 2 model: The AMMI model shown patterns and relationships of genotypes and environments successfully. The hybrid that best adapted to most environments was 30H83 inclining average to favorable environments and it was also stable to all environments. BH-541 was better performing in the lower to high yielding environments (Table 6). FH-625-259xF-7215x144-7-b showed adaptation to specific environments. It is clear that the AMMI model can be used to analyze the GxE and identify the superior genotypes. It can also be used in the selection of the best environments for genotype evaluation.
Cluster analysis: Genotypes were clustered using the AMMI2 estimated values of nine environments used as attributes and, conversely, the environments were grouped using estimated value of AMMI2 for the 15 genotypes. Dendrogram for clustering of cultivars and environments are shown in Fig. 3. At the two group level of genotype clustering genotypes, BH-660, BH-670, FH-625-259xF-7215x144-7-b, BH-544 and BH-541 were discriminated from the remaining. These genotypes are characterized by high yield (above the grand mean) with high positive interaction. In contrast, the second genotypes, BHQP-542, BHQP-543, Ambo Synth-1, Ambo Synth-5, AMH-800 and SC-715 are low yielders (below the grand mean) with a IPCA score ranging from -3 to 3.
| Table 5: | IPCA1, IPCA2 scores and environmental index for nine locations |
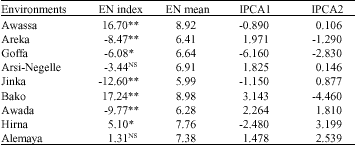 | |
| NS, *, **: Non Significant and significant at p<=0.05 and 0.01 level, respectively, EN mean: Environmental mean and EN index: Environmental index | |
Centroid clustering methods strongly recommended a class of four for this data set. Therefore, splitting the down the main first branch of the dendrogram resulted in two subclusters, while splitting down the second main branch resulted also in two subclusters (Fig. 3a). The first subcluster of the first group comprised genotypes BHQP-543, Ambo Synth-1, Ambo Synth-5 and AMH-800, low yielders and having low interaction. Except genotype BHQP-543 (quality protein maize) all of these genotypes are highland materials developed for highland altitude. The second sub-cluster (BHQP-542, BH-540, SC-715, PHB-3253 and ESE-203) is characterized by genotypes with low yield and negative interactions (Fig. 3a).
The splitting down of the second main branch at two cluster levels simply isolated genotype No.1 and 14 from the rest. As it was observed in the biplot, these genotypes were highest yielders and most adapted to many environments and could probably be the reason for the AMMI2 estimate clustering to group them differently from the other high yielding genotypes. The second sub-cluster (BH-660, BH-670, FH-625-259xF-7215x144-7-b and BH-544) was characterized by genotypes with high yield and high interaction. Three way cross hybrids, BH-660 and BH-670 have the same single cross female parent and they only differ in their male parents.
 | |
| Fig. 3a: | Genotype dendrogram representing four genotype cluster. Where, FH-144-7-b = FH-625-259xF-7215x144-7-b |
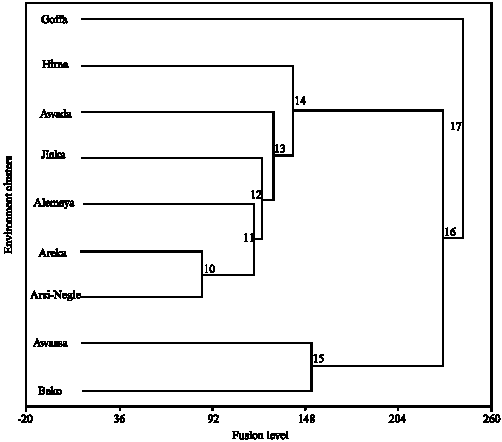 | |
| Fig. 3b: | Environmental dendrogram representing three clusters of test locations |
| Table 6: | Best five high yielding genotypes based on AMMI model selections |
 | |
| FH1447b = FH-625-259xF-7215x144-7-b; Gen1 = 1st yielder at a location; Gen2 = 2nd yielder; Gen3 = 3rd yielder; Gen4 = 4th yielder; Gen5 = 5th yielder at a location | |
Environments were clustered using the nearest neighbor. The three clusters sufficiently described this data set (Fig. 3b). The first group comprised of only environment Goffa and the second group comprised of all environments except Goffa. Goffa is unique environment, where there was high and erratic rainfall and situated at low altitude as compared to others. The splitting down of the second main branch at two cluster levels simply isolated environment Awassa and Bako together which are high yielding (above grand mean) environments. This could also be attributed to similarities between the two locations in altitude, humidity and length of growing seasons.
REFERENCES
- Gabriel, K.R., 1971. The biplot graphic display of matrices with application to principal component analysis. Biometrika, 58: 453-467.
CrossRefDirect Link - Kaya, Y., C. Palta and S. Taner, 2002. Additive main effects and multiplicative interactions analysis of yield performance in bread wheat genotypes a cross environments. Turk. J. Agric., 26: 275-279.
Direct Link - Mandel, J., 1971. A new analysis of variance model for non-additive data. Technometrics, 13: 1-18.
Direct Link - Ngeve, J.M. and J.C. Bouwkamp, 1993. Comparison of statistical methods to assess yield stability in sweetpotato. J. Am. Soc. Hortic. Sci., 118: 304-310.
Direct Link - Scapim, C.A., V.R. Oliveira, A.D.L. Braccini, C.D. Cruz, C.A.D.B. Andrade and M.C.G. Vidigal, 2000. Yield stability in Maize (Zea mays L.) and correlations among the parameters of the eberhart and russell, Lin and Binns and Huehn models. Genet. Mol. Biol., 23: 387-393.
CrossRefDirect Link - Sivapalan, S., L.O. Brien, G. Ortiz-Ferrara, G.L. Hollamby, I. Barclay and P.J. Martin, 2000. An adaptation analysis of Australian and CIMMYT/ICARDA wheat germplasm in Australian production environments. Aust. J. Agric. Res., 51: 903-915.
CrossRefDirect Link - Sneller, C.H., L. Kilgore-Norquest and D. Dombek, 1997. Repeatability of yield stability statistics in soybean. Crop Sci., 37: 383-390.
CrossRefDirect Link - Thillainathan, M. and G.C.J. Fernandez, 2001. SAS applications for Tai's stability analysis and AMMI model in GenotypeHEnvironmental Interaction (GEI) effects. J. Hered., 92: 367-371.
Direct Link - Vargas, M., J. Crossa, F.A. van Eeuwijk, M.E. Ramirez and K. Sayre, 1999. Using partial least squares regression, factorial regression and AMMI models for interpreting genotype x environment interaction. Crop Sci., 39: 955-967.
CrossRefDirect Link - Yan, W., L.A. Hunt, Q. Sheng and Z. Szlavnics, 2000. Cultivar evaluation and mega-environment investigation based on the GGE biplot. Crop Sci., 40: 597-605.
CrossRefDirect Link