
E. Bisimwa
Earth and Life Institute, Applied Microbiology, Phytopathology, Universite catholique de Louvain, Croix du Sud 2 boite L 07.05.03., 1348 Louvain-la-Neuve/Belgium
J. Walangululu
Faculte des Sciences Agronomiques, Universite Catholique de Bukavu, BP 285 Bukavu, Democratic Republic of Congo
C. Bragard
Earth and Life Institute, Applied Microbiology, Phytopathology, Universite catholique de Louvain, Croix du Sud 2 boite L 07.05.03., 1348 Louvain-la-Neuve/Belgium
Asian Journal of Plant Pathology
Year: 2012 | Volume: 6 | Issue: 1 | Page No.: 1-12







ABSTRACT
Cassava mosaic disease epidemiology and molecular characterization of the related viruses monitored during this study in the Sud-Kivu region, Democratic Republic of Congo. Collected epidemiologic data showed a negative correlation with the altitude (elevation) on the disease incidence and its vector population. Disease incidence was 9.5 to 37.8% associated to less than 3 insects per leaf when the elevation is higher than 1500 m while 65 to 100% and more than 10 insects per leaf were recorded where elevation was less than 1000 m. No impact of rainfall has been recorded. Associated molecular cassava begomovirus diagnostic focused on AC2 and AC4 genes revealed only two species occurring in this area, African Cassava Mosaic Virus (ACMV) and East African cassava mosaic virus-Uganda (EACMV-UG) with more EACMV-UG and mixed infections in low elevation area while ACMV were predominant in high elevation sites. Phylogeny analysis revealed a dual coexistence of indigenous and Ugandan spread among ACMV isolates while all EACMV isolates were restricted to Ugandan spread (95 to 100% of amino acids identity). No recombination was revealed in this study, isolates diversity was restricted to nucleotide substitution.
PDF Abstract XML References Citation
Received: November 22, 2011;
Accepted: December 12, 2011;
Published: March 20, 2012
How to cite this article
E. Bisimwa, J. Walangululu and C. Bragard, 2012. Occurrence and Distribution of Cassava Mosaic Begomovirus Related to Agro-ecosystems in the Sud-kivu Province, Democratic Republic of Congo. Asian Journal of Plant Pathology, 6: 1-12.
URL: https://scialert.net/abstract/?doi=ajppaj.2012.1.12
URL: https://scialert.net/abstract/?doi=ajppaj.2012.1.12
INTRODUCTION
Cassava (Manihot esculenta Crantz, Family Euphorbiaceae) is a major food crop in Sub-Saharan Africa and nourishes more than 60% of the Democratic Republic of Congo (DRC) population (FAOSTAT, 2009). Its production is constrained by several pests and diseases among which Cassava Mosaic Disease (CMD) transmitted by whiteflies, Bemisia tabaci, is the most important (Legg et al., 2006).
CMD is well known since the 1930s (Kufferath and Ghesquiere, 1932) but its impact on yield harvest was relatively stable. Before the 1990s, only two virus species were known as CMD causal agents. Actually, seven begomovirus species widely distributed in Sub-Saharan Africa are recognized: the African Cassava Mosaic Virus (ACMV) (Hong et al., 1993; Swanson and Harrison, 1994), the East African Cassava Mosaic Virus (EACMV) and EACMV-like strains (Deng et al., 1997; Zhou et al., 1997; Legg and Ogwal, 1998; Legg, 1999; Legg et al., 2001, 2004, 2006), the East African Cassava Mosaic Cameroon Virus (EACMCV) (Fondong et al., 2000; Ndunguru et al., 2005), the East African Cassava Mosaic Malawi Virus (EACMMV) (Zhou et al., 1998), the East African Cassava Mosaic Zanzibar Virus (EACMZV) (Maruthi et al., 2004), the South African Cassava Mosaic Virus (SACMV) (Berrie et al., 2001; Ranomenjanahary et al., 2002; Mabasa and Rey, 2007), the Indian cassava mosaic virus (Adjata et al., 2008, 2009). Recently, the South East African Cassava Mosaic Virus (SEACMV) was diagnosed in Madagascar and is proposed as a new species by Harimalala et al. (2011).
Following the spread of the epidemic CMD severe, Legg and Fauquet, (2004) have proposed a geographic distribution map on which ACMV occurred in 20 countries, EACMV in 10, EACMV-UG in 11, EACMCV in 5, SACMV in 2, EACMZV in 2 and EACMMV in 1. Since this period the only recorded changes are the ICMV presence in Togo (Adjata et al., 2008, 2009), SACMV in Madagascar and the proposed new species SEACMV in Madagascar (Harimalala et al. 2011).
The begomovirus species diversity is depending on several phenomenon among which recombination (Zhou et al., 1997), pseudo-recombination (Fondong et al., 2000; Pita et al., 2001a) and nucleotides substitution (Duffy and Holmes, 2009) are involved in new species and strains emergence. Despite the DRC second position of African producers, cassava mosaic begomovirus occurrence is less documented and reported. Neuenschwander et al. (2002) study in western DRC, Monde et al. (2010) in Yangambi region, Janssens (2001) and Muyolo (1987) represent the easily available information on cassava in DRC.
Regarding the rapid spread of CMD severe and virus diversity throughout Eastern and Central Africa, this study was planned to provide a preliminary diagnostics and distribution of the Cassava Mosaic Begomoviruses (CMBs) based on field data collection and molecular diagnosis. AC2, involved as Post Transcriptional Gene Silencing (PTGS) suppressor and AC4, an activator of the virus pathogenicity (Vanitharani et al., 2004; Vanitharani et al., 2005) were considered in molecular analysis related to their probable role in plant-virus interaction in expressing the disease symptoms. Additionally, AC2, AC4 and the coat protein are the most diversified zones on DNA of begomovirus genome. Based on the AC2 and AC4 sequences analysis, virus diversity and related causes were assessed.
MATERIALS AND METHODS
Field data and leaf samples collecting: Surveys were conducted in the South-Kivu province, Eastern D.R. of Congo in different cassava growing villages. From Bukavu town, the administrative centre of the province, five directions (Southern, Western, Northern, North-Western and the costal lake-Kivu). Twenty two villages and a total of 150 fields were covered during the survey (data not shown).
Within the selected fields, all the plants on the diagonals were systematically observed independently of the field square. For each plant examined, CMD incidence, CMD severity, the whitefly number (counted from the five top youngest leaves) and the type of infection (whitefly infection or cutting-borne infection) were recorded as proposed by Hahn et al. (1980) and adapted by Okao-Okuja et al. (2004) and Sseruwagi et al. (2004).
Leaf samples were systematically collected in the survey area on diseased plants. Samples were kept in alcohol prior to DNA extraction, Polymerase Chain Reaction (PCR) diagnosis and sequencing realised in the plant pathology laboratory at the Universite catholique de Louvain in Belgium.
Total DNA extraction, PCR amplification and sequencing: Total DNA was extracted from collected leaf samples using the protocol described by Dellaporta (Dellaporta et al., 1983) and the FastDNAR Kit with FastPrepR instruments (Qbiogene, inc., CA).
DNA-A AC2 and AC4 regions specific primers described by Monde et al. (2010) were used.
The mix PCR was prepared in a final volume of 50 μL using H2O depc 26.25 μL, MgCl2 25 mM 5 μL, GoTaq® 5x flexi buffer 10 μL, Dntp 100 mM 1.5 μL, each upstream and downstream primer 1 μL, GoTaq®DNA polymerase (Promega) 5 units μL-1 0.25 μL, extract DNA samples 5 μL. The PCR was cycling by a thermocycler ICycler Biorad® version 4.006 at 94°C for 2 min for denaturizing, followed of 38 cycles of amplification at 94°C for 30 sec for denaturizing, hybridization at 58°C for 30 sec; 72°C for 1 min for elongation. The final elongation has been done at 72°C for 7 min. The revelation was made in 1.2% agarose gel under UV after electrophoresis at 120 V in ethidium bromide. Direct sequencing using Biosequencer Genetic analyser 3100 was made with PCR product diluted tenfold in distilled water. Each amplicon was sequenced many times forward and reverse. Partial sequences produced were translated in amino acids by a molecular toolkit (www.vivo.colostate.edu) and submitted to EMBL EBI server. NCBI accessions numbers were attributed with accessions FN433646 to FN433697. Concatenated amino acids of AC2 and AC4 from the NCBI Genbank and those collected in DR Congo were used to carry out phylogenetic trees using MEGA4.0 software (Tamura et al., 2007). The evolutionary history was determined using the Neighbour-Joining method with the complete deletion option and 1000 replicates for bootstrap values. The expressed mutations in amino acids sequences are shown in front of respective phylogenetic trees by alignment (Fig. 1 and 2).
Concatenated amino acids sequences of AC2 and AC4 of the following NCBI accessions were used: Cameroon [CM] [AF366902]1; Nigeria [NG] [AJ427910]1; Uganda [UG] [AF126800, AF126802]1; [AJ717533, AJ717534, AJ618958, AM502326, AM502327, AM502329, AM502334]2; Ivory Coast [IC] [AF259894]1 and Tanzania [TZ] [AY795988]2.
| • | (1): ACMV and (2): EACMV |
Molecular diversity of the analysed sequences were reported in the geographic map of Sud-Kivu (Fig. 3) established in basis of general map of soil occupation of DR Congo provided by UCL-Geomatics, Louvain la Neuve, Belgium, 2007 where only elavation is highlightened.
RESULTS AND DISCUSSIONS
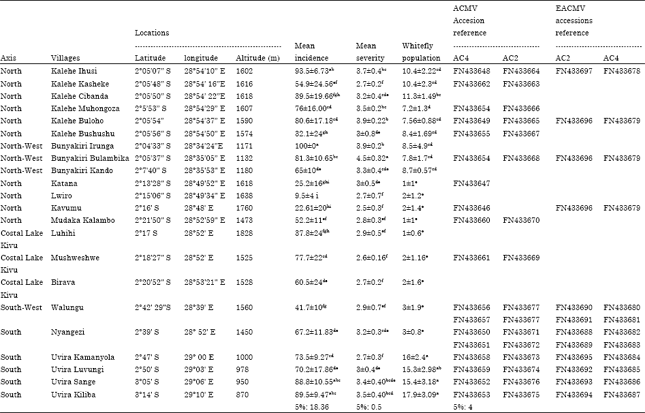
CMD incidence, severity, infection type and whitefly population: CMD incidence, its severity, whitefly population and virus diagnostics are presented in Table 1. The field data collected showed a clear geographic and ecological distribution of CMD incidence, its severity and the whitefly population in the surveyed area. CMD mean incidence values were moderate to high, ranging from 9.48 to 100% with the highest values recorded in the low altitude of Uvira (high temperature values, grassland and intensive agriculture environment) and Bunyakiri (high temperature values, secondary forest) where more fields were recorded with 85 to 100% of diseases plants than anywhere else (Table 1).
 | |
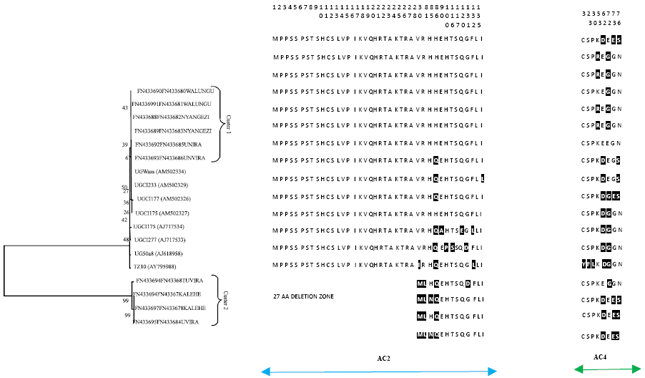
| Fig. 1: | Phylogenetic and molecular analysis based on amino acids in concatenated sequences of AC2 and AC4 for ACMV inferred from the Neighbour-joining method (Saitou and Nei, 1987) using MEGA software version 4.0 with complete deletion option. Bootstrap values (1000 replicates) are shown as percentages in the branches (Tamura et al., 2007). The phylogenetic tree was constructed with AC2 and AC4 sequences from Cameroon [CM] AF366902; Ivory Coast [IC] AF259894; Nigeria [Ng] AJ427910; Uganda [UG] AF126800, AF126802 and Democratic Republic of Congo FN433646, FN433647, FN433648, FN433649, FN433650, FN433651, FN433652, FN433653, FN433654, FN433655, FN433656, FN433657, FN433658, FN433659, FN433660, FN433661, FN433662, FN433663, FN433664, FN433665, FN433666, FN433667, FN433668, FN433669, FN433670, FN433671, FN433672, FN433673, FN433674, FN433675, FN433676, FN433677 |
 | |
| Fig. 2: | Phylogenetic and molecular analysis based on amino acids in concatenated sequences of AC2 and AC4 for EACMV-UG inferred from the Neighbour-joining method (Saitou and Nei, 1987) using MEGA software version 4.0 with complete deletion option. Bootstrap values (1000 replicates) are shown as percentages in the branches (Tamura et al., 2007). The phylogenetic tree was constructed with AC2 and AC4 sequences from Uganda [Ug]AM502327, AM502329, AM502326, AM502334, AJ717534, AJ717533, AJ618958; Tanzania [Tz]AY795988 and Democratic Republic of Congo FN 433678, FN433679, FN433680, FN433681, FN433682, FN433683, FN433684, FN433685, FN433686, FN433687, FN433688, FN433689, FN433690, FN433691, FN433692, FN433693, FN433694, FN433695, FN433696, FN433697 |
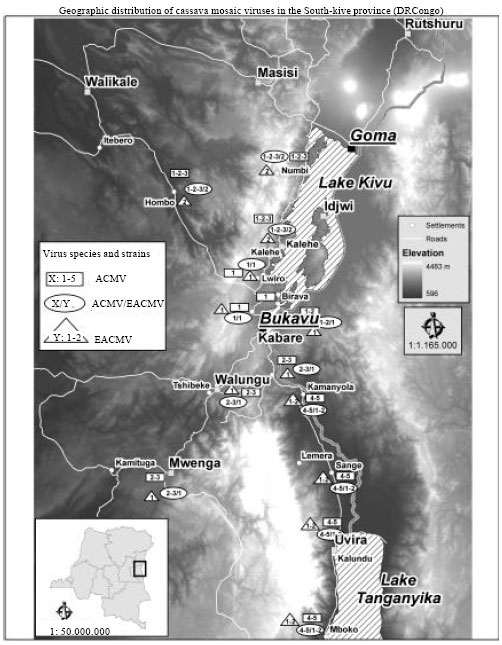 | |
| Fig. 3: | Cassava mosaic Begomovirus distribution on the Sud-Kivu map. The location of sampling were made in the basis of the general map of DR Congo. Five different isolates of ACMV (1 to 5) and two isolates of EACMV-UG (1 and 2) were identified by molecular analysis based on AC2 and AC4 genes. Fractional numbers (X/Y) represent the dual infections by both ACMV and EACMV-UG |
In those villages more than 50% of infected plants were recorded with high severity score 3-5 and cassava asymptomatic plants in the fields were rare. The high incidence and severity values were positively correlated to whitefly population (7-17.9 insects per leaf) depending on the local environment temperature.
| Table 1: | CMD incidence, severity and whitefly population and CMD diagnostic in the South-kivu province. Number with different succeeding alphabets are significantly different at p = 0.05 level, mean incidence was estimated as a percentage of CMD diseased plants in a field and in a village. Mean severity was estimated by using a 1-5 scale proposed by Sseruwagi et al. (2004) where score 1 is attributed to a cassava plant CMD symptom free and 5 to a cassava plant with CMD severe symptoms. Whitefly population was estimated as a mean whitefly number per leaf on the 5 top cassava leaves |
 | |
| Mean values with same letters are not signifficantly LSD different LSD at p = 0.05 | |
Xie et al. (2011) have mentioned that whitefly fecundity and development were significantly increased at 24°C comparing to 15, 18 and 21°C.
However, great values of CMD incidence, severity and whitefly population were recorded at Kalehe, despite its location in altitude, moderate temperature and intensive agriculture environment. The lowest values were observed in the high altitude (Lwiro, Kavumu, Katana, Luhihi) where 9.5 to 37.8% of infected plants were recorded. The lowest value was observed in Lwiro (9.5%) where most of the cassava plants on the fields were asymptomatic, CMD severity score moderate (2-3) and whitefly population low (2 insects per leaf).
The high incidence and CMD severity score observed in the low altitude area (Uvira, Kalehe and Bunyakiri) was recorded in a positive correlation with the EACMV presence, alone or in mixed infections, in cassava fields with 57.2; 64.2 and 63.7% EACMV-positive samples respectively. These zones can be considered with the high epidemic index 3 (Legg et al., 2006). All sampled fields were predominantly grown with local and susceptible genotypes, among which Nambiyombiyo, M’Bailo and Cizinduka are most cultivated in altitude zones whereas Nakarasi and Naunde are most cultivated in Uvira zones.
The CMD severity score all around the covered area was ranged from 2.3 at Kavumu to 5 at Bunyakiri and Uvira with a mean score of 3.5 for the three most affected zones (Uvira, Kalehe and Bunyakiri). There was no genotypes variability on the incidence and severity, homogenous results have been recorded for all the local genotypes observed.
The whitefly number counted on upper cassava leaves varied from the different villages with low mean value of 1-3 insects per leaf in the high altitude (Katana, Luhihi, Lwiro, Kavumu, Nyangezi, Walungu and Mudaka) to 17.9 insects per leaf, the highest value observed in low altitude in Uvira-Kiliba. The positive correlation of white fly population, CMD incidence and severity and EACMV presence reaffirms the hypothesis EACMV to be easily transmitted by whitefly than ACMV (Colvin et al., 2004). The type of infection was homogenously distributed in each ecological zone, roughly 75 to 80% for cutting-borne infection and 20 to 25% for whitefly infection, except in Kamanyola where whitefly infection is 32.6 versus 67.4% for cutting-borne infection. These results suggest an ecological impact, mainly the temperature, on CMD vector development and its impact on the disease transmission and spread.
PCR diagnostic showed that only two species are occurring in the region, ACMV and EACMV species. They were diagnosed alone or in mixed infections in all cassava fields in the survey area except in the villages all along and near the lake Kivu (Luhihi, Mushweshwe and Birava) where only ACMV has been found (87%). EACMV-UG was found alone or in mixed infections with ACMV. Great rates of mixed infections were frequently associated to severe CMD symptoms observed at Bunyakiri, Kalehe and Uvira as it has been observed in Uganda, Kenya, Tanzania and Rwanda, suggesting a synergistic interaction (Harrison et al., 1997; Legg, 1999; Pita et al., 2001b; Fondong et al., 2000).
The blast of EACMV sequences showed high similarity to EACMV-UG (more than 98% of nucleotide and amino acids homology) (Fig. 2). This study is a report on EACMV-UG spread to Sud-Kivu province where it hasn’t been reported yet. The high frequency of EACMV-UG isolates in the north and south means the spread was realized from two different movements, at the north probably from Uganda when at the south, the spread is probably from Burundi, where it has been reported earlier (Bigirimana et al., 2004).
The concatenated AC2 and AC4 amino acids sequences alignment with Ugandan isolates showed that AC2 region of both ACMV and EACMV-UG is less diversified with less than 6% of amino acids substitution. The same observation was made for AC4 of EACMV-UG with less than 4% of amino acids substitution while the same region of ACMV was the most diversified with 6 to 11% of amino acids substitutions (Fig. 1 and 2). The AC4 of ACMV variability percentage is closed to what have been mentioned by Ndunguru et al. (2005) and Duffy and Holmes, (2009) for cassava begomovirus diversity and evolution and confirms ACMV occurring in the region for a long time, while the EACMV-UG less diversity is related to its recent introduction in the region.
The phylogenetic trees of different isolates established based on concatenated amino acids of AC2 and AC4 (Fig. 1 and 2) showed a single ACMV group of truncated AC4 gene presenting a premature codon stop and segregated in five different clusters while all EACMV-UG isolates were segregated in two different clusters.
The three first groups of ACMV are widely distributed in the region and linked to moderate CMD symptoms, while the groups 4 and 5 are restricted to Uvira zone and linked to severe CMD symptoms. The cluster 5 contains one isolate which is completely different from the other isolates and contains a specific fragment length of twenty five amino acids (22.9% of amino acids substitution) at the beginning of the AC2 region. The blast of the differential fragment didn’t reveal any recombination from a known begomovirus, nucleotides and amino acids substitution was the only one way of the virus diversity and evolution identified in this study.
The EACMV-UG isolates are closed to Uganda isolates and segregated in two clusters. The first cluster contains isolates widely distributed in the surveyed area which have homogenous AC2 region and weak AC4 amino acids substitution (3.8%) comparing to Ugandan isolates. The second cluster contains isolates restricted to kalehe and Uvira zones which present a deletion of twenty seven amino acids on the beginning of AC2 region, while AC4 region contains a weak amino acids substitution (3.8%). This particular isolates, both ACMV and EACMV-UG, restricted to Kalehe and Uvira is one of the probable causes of severe CMD symptoms observed in this area beside a great rate of ACMV/EACMV dual infections and the intensive cultivation of local and susceptible cassava genotypes.
The spread to Great Lake region (DRC, Rwanda and Burundi) of EACMV-UG from East African countries was expected to introduce the cassava begomovirus diversity reported by Ndunguru et al. (2005) in its origin region, in contrast no cassava begomoviruses species diversity has been reported by this study similar to the Rwandan and Burundian cases where both ACMV and EACMV-Ug only were reported (Bigirimana et al., 2004; Legg et al., 2001). In the East-Northern DRC, only those two species were reported by Monde et al. (2010) study. In relation to these observations, it can be assume that the Ugandan CMD spread to Great Lake countries was realized before recombination processes. However, recombinations result from the coexistence of several begomovirus in the same environment and no additional favourable conditions are reported to influence this phenomenon. In this way, frequent begomovirus recombinations reported in East Africa can easily occur in the Great lake region and there is no reason to not fear such threat in the Great Lake region where the two species coexistence is reported since ten years. Curiously, the first phenomenon of begomovirus recombination in Uganda was reported related to severe CMD disease (Deng et al., 1997; Zhou et al., 1997) only four years after Hong et al. (1993) report on the coexistence of ACMV and EACMV in East Africa region. Thus, there is no evident explanation of this and more care is needed to permanently monitor the disease epidemiology and aetiology in this region.
ACKNOWLEDGMENTS
This study was achieved under a Belgian technical cooperation (BTC) scholarship associated to technical support of the Earth and Life Institute Applied Microbiology-Phytopathology of the University catholic of Louvain in Belgium and the Catholic University of Bukavu in DR Congo. Authors are grateful to UCL-ENGE-Geomatics for providing the map.
REFERENCES
- Adjata, K.D., E. Muller, M. Aziadekey, Y.M.D. Gumedzoe and M. Peterschmitt, 2008. Incidence of cassava viral diseases and first identification of East African cassava mosaic virus and Indian cassava mosaic virus by PCR in Cassava (Manihot esculenta Crantz) fields in Togo. Am. J. Plant Physiol., 3: 73-80.
CrossRefDirect Link - Adjata, K.D., E. Muller, M. Peterschmitt, O. Traore and Y.M.D. Gumedzoe, 2009. Molecular evidence for the association of a strain of Uganda variant of east African cassava mosaic virus to symptom severity in cassava (Manihot esculenta Crantz) fields in togo. Am. J. Biochem. Biotechnol., 5: 196-201.
Direct Link - Berrie, L.C., E.P. Rybicki and M.E. Rey, 2001. Complete nucleotide sequence and host range of South African cassava mosaic virus: Further evidence for recombination amongst begomoviruses. J. Gen. Virol., 82: 53-58.
Direct Link - Bigirimana, S., P. Barubanze, R. Obonyo and J.P. Legg, 2004. First evidence for the spread of East African cassava mosaic virus: Uganda (EACMV-Ug) and the pandemic of severe cassava mosaic disease in Burundi. Plant Pathol., 53: 231-231.
CrossRefDirect Link - Colvin, J., C.A. Omongo, M.N. Maruthi, G.W. Otim Nape and J.M. Thresh, 2004. Dual begomovirus infection and high Bemisia tabaci populations: Two factors driving spread of cassava mosaic disease pandemic. Plant Pathol., 53: 577-584.
CrossRefDirect Link - Duffy, S. and E.C. Holmes, 2009. Validation of high rates of nucleotides substitution in Geminiviruses: Phylogenetic evidence from East African cassava mosaic viruses. J. Gen. Virol., 90: 1539-1547.
CrossRefDirect Link - Fondong, V.N., J.S. Pita, M.E.C. Rey, A. de Kochko, R.N. Beachy and C.M. Fauquet, 2000. Evidence of synergism between African cassava mosaic virus and a new double-recombinant Geminivirus infecting cassava in Cameroon. J. Gen. Virol., 81: 287-297.
CrossRefDirect Link - Hahn, S.K, E.R. Terry and K. Leuschner, 1980. Breeding cassava for resistance to cassava mosaic disease. Euphytica, 29: 673-683.
CrossRefDirect Link - Harrison, B.D., X. Zhou, G.W. Otim-Nape, Y. Liu and D.J. Robinson, 1997. Role of a novel type of double infection in the geminivirus-induced epidemic of severe cassava mosaic in Uganda. Ann. Applied Biol., 131: 437-448.
CrossRefDirect Link - Hong, Y.G, D.J. Robinson and B.D. Harrison, 1993. Nucleotide sequence evidence for the occurrence of three distinct whitefly-transmitted geminiviruses in cassava. J. Gen. Virol., 74: 2437-2443.
PubMedDirect Link - Legg, J.P. and S. Ogwal, 1998. Changes in the incidence of African cassava mosaic virus disease and the abundance of its whitefly vector along South-North transects in Uganda. J. Applied Entomol., 122: 169-178.
CrossRefDirect Link - Legg, J.P., 1999. Emergence, spread and strategies for controlling the pandemic of cassava mosaic virus disease in East and Central Africa. Crop Protect., 18: 627-637.
CrossRefDirect Link - Legg, J.P., F. Ndjelassili and G. Okao-Okuja, 2004. First report of cassava mosaic disease and cassava mosaic geminiviruses in Gabon. Plant Pathol., 53: 232-232.
CrossRefDirect Link - Legg, J.P., G. Okao-Okuja, R. Mayala and J.B. Mahinyuza, 2001. Spread into Rwanda of the severe cassava mosaic virus disease pandemic and the associated Uganda variant of the East African Cassava Mosaic Virus ( EACMV-Ug). Plant Pathol., 50: 796-796.
Direct Link - Legg, J.P. and C.M. Fauquet, 2004. Cassava mosaic geminiviruses in Africa. Plant Mol. Biol., 56: 585-599.
PubMed - Legg, J.P., B. Owor, P. Sseruwagi and J. Ndunguru, 2006. Cassava mosaic virus disease in East and Central Africa: Epidemiology and management of a regional pandemic. Adv. Virus Res., 67: 355-418.
CrossRefPubMedDirect Link - Maruthi, M.N., S. Seal, J. Colvin, R.W. Briddon and S.E. Bull, 2004. East African cassava mosaic Zanzibar virus: A recombinant begomovirus species with a mild phenotype. Arch. Virol., 149: 2365-2377.
CrossRefDirect Link - Monde, G., J. Walangululu, S. Winter and C. Bragard, 2010. Dual infection by cassava begomoviruses in two leguminous species (Fabaceae) in Yangambi, North-Eastern Democartic Republic of Congo. Arch. Virol., 155: 1865-1869.
PubMedDirect Link - Ndunguru, J., J.P. Legg, T.A.S. Aveling, G. Thompson and C.M. Fauquet, 2005. Molecular biodiversity of cassava Begomovirus in Tanzania: Evolution of cassava geminiviruses in Africa and evidence for East Africa being a center of diversity of cassava geminiviruses. Virol. J., 2: 21-21.
PubMedDirect Link - Neuenschwander, P., J. Hughes, F. Ogbe, J.M. Ngatse and J.P. Legg, 2002. Occurrence of the Uganda variant of East African Cassava Mosaic Virus (EACMV-Ug) in Western Democratic Republic of Congo and the Congo Republic defines the westernmost extent of the CMD pandemic in East/Central Africa. Plant Pathol., 51: 385-385.
Direct Link - Okao-Okuja, G., J.P. Legg, L. Traore and A.M. Jorge, 2004. Viruses associated with cassava mosaic disease in Senegal and Guinea Conakry. J. Phytopathol., 152: 69-76.
CrossRefDirect Link - Pita, J.S., V.N. Fondong, A. Sangaré, R.N.N. Kokora and C.M. Fauquet, 2001. Genomic and biological diversity of the African cassava geminiviruses. Euphytica, 120: 115-125.
CrossRefDirect Link - Pita, J.S., V.N. Fondong, A. Sangare, G.W. Otim-Nape, S. Ogwal and C.M. Fauquet, 2001. Recombination, pseudorecombination and synergism of geminiviruses are determinant keys to the epidemic of severe cassava mosaic disease in Uganda. J. Gen. Virol., 82: 655-665.
CrossRefDirect Link - Ranomenjanahary, S., R. Rabindran and D.J. Robinson, 2002. Occurrence of three distinct begomoviruses in cassava in Madagascar. Ann. Applied Biol., 140: 315-318.
CrossRef - Sseruwagi, P., W.S. Sserubombwe, J.P. Legg, J. Ndunguru and J.M. Thresh, 2004. Methods of surveying the incidence and severity of cassava mosaic disease and whitefly vector populations on cassava in Africa: A review. Virus Res., 100: 129-142.
CrossRefPubMedDirect Link - Tamura, K., J. Dudley, M. Nei and S. Kumar, 2007. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol., 24: 1596-1599.
CrossRefPubMedDirect Link - Vanitharani, R., P. Chellappan, J.S. Pita and C.M. Fauquet, 2004. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of Posttranscriptional gene silencing. J. Virol., 78: 9487-9498.
CrossRefDirect Link - Vanitharani, R., P. Chellappan and C.M. Fauquet, 2005. Geminiviruses and RNA silencing. Trends Plant Sci., 10: 144-151.
PubMed - Xie, M., F.H. Wan, Y.H. Chen and G. Wu, 2011. Effects of temperature on the growth and reproduction characteristics of Bemisia tabaci B-biotype and Trialeurodes vaporariorum. J. Applied Entomol., 135: 252-257.
CrossRef - Zhou, X., Y. Liu, L. Calvert, C. Munoz, G.W. Otim-Nape, D.J. Robinson and B.D. Harrison, 1997. Evidence that DNA-A of a Geminivirus associated with severe cassava mosaic disease in Uganda has arisen by interspecific recombination. J. Gen. Virol., 78: 2101-2111.
CrossRefDirect Link - Zhou, X., D.J. Robinson and B.D. Harrison, 1998. Types of variation in DNA-A among isolates of East African cassava mosaic virus from Kenya, Malawi and Tanzania. J. Gen. Virol., 79: 2835-2840.
PubMed - Dellaporta, S.L., J. Wood and J.B. Hicks, 1983. A plant DNA minipreparation: Version II. Plant Mol. Biol. Rep., 1: 19-21.
CrossRefDirect Link - Saitou, N. and M. Nei, 1987. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol., 4: 406-425.
CrossRefPubMedDirect Link