
M.M. Azooz
Department of Biological Sciences, College of Science, King Faisal University, Saudi Arabia
M.M. Youssef
Department of Chemistry, College of Science, King Faisal University, Saudi Arabia
M.A. Al-Omair
Department of Chemistry, College of Science, King Faisal University, Saudi Arabia
American Journal of Plant Physiology
Year: 2011 | Volume: 6 | Issue: 6 | Page No.: 269-282







ABSTRACT
Soil contamination with heavy metals has become a worldwide problem, leading to a reduction of plant growth and productivity. The objective of this study was carried out to compare the effect of different levels of zinc (Zn2+) or lead (Pb2+)and their interactions on growth and physiological changes of Hassawi okra seedlings. These seedlings were grown in a soil, to which different levels (0, 5, 10, 20, 40, 80 mM) of Zn2+ or Pb2+ were added with water (control), singly or in combinations. The results revealed that, okra seedlings growth were reduced gradually with increasing Zn2+ or/and Pb2+. These parameters were completely inhibited at 80 mM of Zn2+ or Pb2+, when they added in combination. While, 5 mM Zn2+ had a favorable effect on these growth parameters. The contents of chl. a and chl. b were significantly reduced as a result of exposure to Zn2+ or/and Pb2+. However, carotenoids content was increased at lower levels as compared with control. The increased of Zn2+ or/and Pb2+ levels inhibited sugars accumulation while the contents of protein, total free amino acids and proline were promoted. Pb2+ had more toxic effect than Zn2+. The toxicity caused by each metal alone was lower than that caused when they were added in combinations. Our data suggested that Zn2+ and Pb2+ exhibited synergistic effects on the growth and physiological responses of Hassawi okra seedlings, leading to injurious effects followed by death of these seedlings at 80 mM. Combined exposure to Zn2+ and Pb2+ enhanced or inhibited some of the effects, that were induced when only one metal was applied to growth medium.
PDF Abstract XML References Citation
Received: July 14, 2011;
Accepted: September 27, 2011;
Published: December 03, 2011
How to cite this article
M.M. Azooz, M.M. Youssef and M.A. Al-Omair, 2011. Comparative Evaluation of Zinc and Lead and their Synergistic Effects on Growth and Some Physiological Responses of Hassawi Okra (Hibiscus esculentus) Seedlings. American Journal of Plant Physiology, 6: 269-282.
DOI: 10.3923/ajpp.2011.269.282
URL: https://scialert.net/abstract/?doi=ajpp.2011.269.282
DOI: 10.3923/ajpp.2011.269.282
URL: https://scialert.net/abstract/?doi=ajpp.2011.269.282
INTRODUCTION
Heavy metal pollution in air, water and agricultural soils is one of major ecological concern due its impact on human health through the food chain and its high persistence in the environment (Valko et al., 2005; Jiang et al., 2010). Environmental pollution by toxic metals originate from many activities, largely industrial (Waisberg et al., 2003). Soil contamination with Heavy Metals (HM) is a worldwide problem, leading to agricultural losses and hazardous health effects as metals enter the food chain. HM can infect soils in town areas, between many of them, Zn2+ and Pb2+ deriving from galvanized metals, rubber production batteries, petrol, steel mill residues and paints (Garg and Aggarwal, 2011).
The phytotoxicity of the heavy metals due to industrial pollution has serious implications in soil degradation (Friedland, 1990). This may reduce both the quality and productivity of plants. Although, Zn is an essential element to plants, excess Zn inhibits plant growth and impairs some important cellular processes. Zinc concentrations in plant above than 300 mg kg-1 are considered toxic (Marschner, 1995). It is an essential trace element for the normal healthy growth and reproduction of some plants and present in enzymatic system. Genc et al. (2006) reported that zinc has vast members of functions in plant metabolism. Although, Zn2+ required in low concentrations in the plant organs (Parker et al., 1992). It is highly toxic at elevated concentrations and can slow down plant growth and disrupt various essential physiological processes (Clemens, 2006). Zn2+ was decreased respiratory rate and increased of membrane damage in sunflower and safflower plants and affected the accumulation of other nutrients in different plants (Ismail and Azooz, 2005). Zn2+ at high levels leads to chlorosis in young leaves, inhibits photosynthesis at various steps and decreases the growth and metabolic activity in various plant species (Panda et al., 2003; Vaillant et al., 2005). Excess zinc in the soil can alter certain physiological equilibrium because the similarities in ion radius of bivalent cations and their local competition at various sites (Monnet et al., 2001). It has been reported that phototoxic concentrations of Zn2+ induce lipid peroxidation, thereby enhancing antioxidative activity in plants (Weckx and Clijsters, 1997).
Lead (Pb2+) is a heavy metal, with characteristic toxic action and main sources of environmental pollution. It inhibits some of metabolic activities in the plant such as the biosynthesis of nitrogenous compounds, carbohydrate metabolism and water absorption (Sharma and Dubey, 2005; John et al., 2008; Hamid et al., 2010). Plants exposed to Pb2+ showed a considerable decrease in their dry weight and a decline in the total chlorophyll and those photosynthetic efficiency (Kosobrukhov et al., 2004; Kambhampati et al., 2005). The plant processes are adversely affected by increasing Pb2+ levels in soils and even at very low concentration (Patra et al., 2004).
Plants have developed defense strategies against heavy metals. They have developed two basic strategies to resist the toxicity of heavy metals: avoidance and accumulation (Tang et al., 2009). Most heavy metal-tolerant species have the capability of preventing heavy metal accumulation in their shoots and therefore are called excluders while others can take up heavy metals, translocate them into the shoots and requisition them in non-metabolic-active tissues and organs in less harmful forms and these plants, called hyper-accumulators (Kupper et al., 2007).
Okra Hassawi plant (Hibiscus esculentus L., member of Malvaceae) is cultivated widely in Al-Hassa region (eastern region) in Saudi Arabia. The plant organs, such as flowers and green tissues, may be consumed as foods while the seed oil may be used for industrial or food purposes (Andras et al., 2005).
The knowledge about the toxic effects of Zn2+ and Pb2+ on the physiological processes in Hassawi okra is potentially useful to establish criteria for selection of plants to be used as bio-indicators. Therefore, the purpose of this investigation was to evaluate the influence of an additional supply of Zn2+ and Pb2+ (in nitrate form) on: (1) growth parameters and (2) some physiological activities in Hassawi okra seedlings. This may be allowed to find differential characteristics suitable to be used as bio-indicators of toxic levels of Zn2+ and Pb2+. In addition, this study will provide better understanding to the tolerance mechanisms of Hassawi okra to the toxicity of Zn2+ and Pb2+. The treatments were added to the soil in solution with water only (control), one metal and with a mixture of both. The mixture of metals was included in order to determine the occurrence of any antagonistic or synergistic interactions between Pb2+ and Zn2+.
MATERIALS AND METHODS
Plant growth and treatments: The experiments were carried out within the period of September 2010-February 2011, in the Faculty of Science, King Faisal University. Homogenous Hassawi okra seeds were chosen and surface sterilized with 0.1% sodium hypochlorite for 5 min and washed three times with sterile water. Sterile seeds were germinated in dark on plastic trays covered with wet filter papers and were kept wet by spraying with sterile water. After complete germination, ten uniformly germinated seeds were sown in the prepared soil at a depth of 1 cm in each pot. Seedlings of okra (Hibiscus esculentus cv. Hassawi) were grown in plastic pots filled with soil composed of air dried clay and sand (1:1 by volume) in growth chamber under constant conditions. The Hassawi okra germinated seeds were divided into 4 groups. The first group was supplied with water (control). The second and the third groups were supplied with 5, 10, 20, 40 and 80 mM Zn (NO3)2 and Pb (NO3)2, respectively. The forth group was supplied with 5, 10, 20, 40 and 80 mM of both Zn (NO3)2 and Pb (NO3)2. The soil was irrigated with the applied concentrations of Zn2+ and Pb2+ or both. Three replicates for each treatment were prepared. The plants were watered with normal water to field capacity by weighing the pots every two days and left to grow until the end of experiment (30 Days after Sowing (DAS)).
Determination of growth parameters: At the end of experiment, Hassawi okra seedlings were harvested and washed with distilled water three times. Leaf area, root and shoot length of seedlings were determined. To determine fresh and dry weight of seedlings, the freshly seedlings are rinsed with deionized water and blotted on paper towels before being weighed (fresh weight). To determine the dry matter yields, the freshly tissues were dried in an aerated oven at 80°C to constant weight.
Photosynthetic pigments content (chl a and chl b and total carotenoids) were estimated in 80% acetone extracts using the spectrophotometric method according to Lichtenthaler and Wellburn (1983).
Membrane stability index (MSI): Leaf Membrane Stability Index (MSI) was determined according to the method of Premchandra et al. (1990) as modified by Sairam (1994).
Determination of some metabolic activities: Sugars were extracted from the plant tissues and quantified by the anthrone sulphuric acid method described by Fales (1951). The soluble and insoluble proteins were determined according to Bradford (1976). Free amino acids were extracted from plant tissues and determined according to the method of Lee and Takahashi (1966). Proline was determined according to Bates et al. (1973).
Statistical analysis: Statistical analysis of data was conducted using one-way Analysis of Variance (ANOVA) using SPSS 12.0 software. Values in the figures indicate the mean values±SD based on independent three determinations (n = 3). Least Significant Difference (LSD) test was used to assess the differences between control and different treatments; p≤0.05 was considered statistically significant.
RESULTS AND DISCUSSION
Growth parameters: The growth parameters were used as a useful bio-indicators of zinc or/and lead toxicity in okra seedlings. These parameters are expressed as leaf area, root and shoot length and fresh and dry weight (Fig. 1a-e). Increasing of Zn2+ or/and Pb2+ levels exhibited gradual reduction in these parameters. While, they were exhibited injurious effects followed by the death of the okra seedlings, when they added in combination at the highest (80 mM) level (So, data do not appear in the all figures at this level). It is worthy to note that, Zn2+ at low level (5 mM) promoted the growth parameters in comparing to their respective control. Inhibition of growth and reduction of biomass production are general responses of some higher plants to Zn2+ or/and Pb2+ toxicity and are often a reliable indication of plant's sensitivity to their stresses (Kosobrukhov et al., 2004; Vaillant et al., 2005; Sharma et al., 2009). The reduction in growth could be due to the suppression of the elongation growth of cells as reported by John et al. (2008). The inhibition of root growth could lead to the slowing down of the water absorption and mineral elements and consequently affects the growth of okra seedlings.
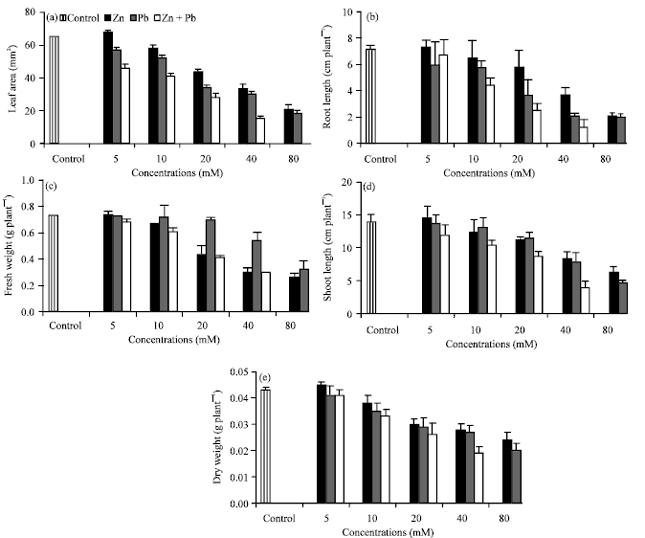 | |
| Fig. 1(a-e): | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on (a) leaf area (mm2), length (cm plant-1) of (b) root and (c) shoot and (d) fresh and (e) dry weight (g plant-1) of Hassawi okra seedlings. Vertical bars represent ±SD |
The inhibitory effects on growth parameters caused by Pb2+ was more phytotoxic than that of Zn2+, suggesting that the okra seedlings were more sensitive to the toxicity of Pb2+ than Zn2+ metal. In addition, Zn2+ is an essential element for plants and may be absorbed and become involved in metabolic pathways more readily than Pb2+ (Hardiman et al., 1984). The effects of Zn2+ and Pb2+ in combination were much severe than that of Zn2+ or Pb2+ alone, leading to the injurious effects followed by the death of the okra seedlings at the highest levels which may suggest a synergistic effect between Zn2+ and Pb2+ on okra seedlings growth. Similar inhibitory effects in co-presence of metals have also been observed in other plant species (Ruley et al., 2006; January et al., 2008; Israr et al., 2011). Guo et al. (2007) showed that the synergistic effects of heavy metal burdens are significantly more toxic than the individual heavy metal exposure. The enhancement effect of Zn2+ at the lower concentration on growth parameters was recommended by Ismail and Azooz (2005) and Sharma et al. (2009). This may be attributed to the beneficial vital role of Zn2+ in plant growth and development. Wierzbicka and Obidziska (1998) reported that Zn2+ is a micronutrient and indispensable for plant growth. Bagci et al. (2007) suggested that adequate applied of Zn2+ improved the water use efficiency of wheat plant. However, Zeid (2001) concluded that, at critical level of Zn2+ could behave as toxic metal like other heavy metals. These reports are in a good agreement and supported our results in respect to the biphasic effects of Zn2+ on the growth parameters of Hassawi okra seedlings.
Photosynthetic pigments: The contents of photosynthetic pigments such as chl. a, chl. b, total chlorophyll and carotenoids of Hassawi okra leaves, grown under different levels of Zn2+ or/and Pb2+ (Fig. 2a-e) indicated that, there were no significant changes on the contents of chl. a, chl. b up to the level of 10 mM Zn2+ or Pb2+, thereafter, a significant decrease were obtained. On the other side, the content of chl. a (at lower concentration of Zn) and carotenoids (at the most concentrations of Zn2+ or Pb2+) was increased as compared with control. The reduction in the content of chl. b was higher (43.3 and 60.9%) than chl. a (34.5 and 50.6%) below the control, when Hassawi okra seedlings were exposed to the highest Zn2+ and Pb2+ levels respectively, resulting in a higher chl. a/b ratio than in control. The reductions in photosynthetic pigments were more obvious in case of Pb2+ than Zn2+and in binary than in individually. Reduction in chlorophyll contents by excess Zn2+ and Pb2+ and increased of carotenoid content have been documented by other investigators (Ismail and Azooz, 2005; Sharma et al., 2009; Hamid et al., 2010). The biosynthesis of photosynthetic pigments is believed to be due to the adequate supply of metal ions. Thus, the excess supplies of Zn2+ and/or Pb2+ may be prevent the incorporation of Fe in phytoporphyrin molecule resulting in the reduction of chlorophyll pigments (Nyitrai et al., 2002; Jaleel et al., 2009). Sharma and Dubey (2005) reported that plants exposed to Pb2+ showed a decline in the photosynthetic rate as a result of restrained synthesis of chlorophyll. This may be as a result of increased in chlorophyllase activity (Drazkiewicz, 1994). Increasing of carotenoids content in the Hassawi okra seedlings at the most levels of Zn2+ or Pb2+ could be enhanced their capacity to minimize the toxic effect caused by these metals. Singh et al. (2006) reported that the enhancement in carotenoid content in heavy metal-treated plants, is probably a part of strategy adopted by the plant to alleviate the toxic effect of free radicals generated under heavy metal toxicity. Hu and Sparks (1991) reported that Zn2+ plays a key role in photosynthesis affecting chlorophyll synthesis. The observed increased in chlorophyll content at lower level of Zn2+ may be attributed to the better growth of okra seedlings. Sharma et al. (2009) suggested that increased of chlorophyll content at lower concentration of Zn2+ in Cicer artietinun might be due to eagerly gathering of Zn2+ in the leaf which significantly affects metabolic processes in the chloroplast.
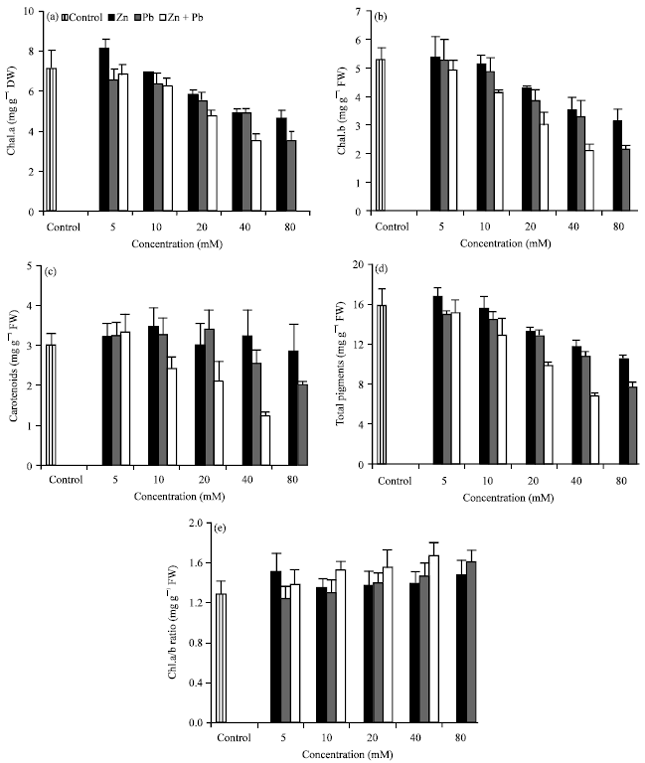 | |
| Fig. 2(a-e): | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on the leaf contents (mg g-1 fresh weight) of (a) chl. a (b) chl. b (c) carotenoids, (d) total chlorophyll and (e) chl. a/chl. b ratio in leaves of Hassawi okra seedlings. Vertical bars represent ±SD |
Sugar contents: The effect of Zn2+ or Pb2+ individually or in combination in Hassawi okra seedlings on reducing, non-reducing and total sugars (Fig. 3a-c) showed that, Zn2+ exhibited positive effect on the reducing sugars at most levels while a decreasing trend in the content of other sugar fractions was observed as a result of increasing in Zn2+ or/and Pb2+ levels when compared with control.
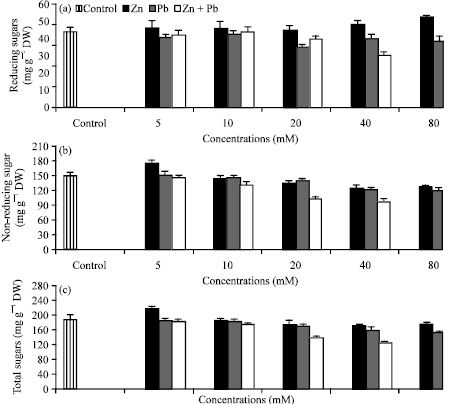 | |
| Fig. 3(a-c): | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on the contents (mg g-1 dry weight) of (a) soluble (b) insoluble and (c) total sugars of Hassawi okra seedlings. Vertical bars represent ±SD |
This decrease was more obvious when Zn2+ and Pb2+ were applied binary than individually; similarly with early other reports (Farshian et al., 2007; Jaleel et al., 2009; Hamid et al., 2010). The increases of reducing sugar at most levels of Zn2+ could be improve the tolerance of Hassawi okra seedlings to zinc toxicity. Verma and Dubey (2001) suggested that higher concentration of soluble sugar could possibly provide an adaptive mechanism in maintaining favorable osmotic potential under toxicity of heavy metals. This was in a good coincidence with our results and explains the slighter toxicity caused by Zn2+ in comparing to Pb2+ on the growth parameters of Hassawi okra seedlings. The reduction in sugar fractions may be due to the decrease observed in chlorophyll content and the abnormal structure of chloroplast which led to decrease photosynthesis efficiency (Farshian et al., 2007). A synergistic effect of both metals was detected in the seedlings exposed to the combination effect of both Zn2+ and Pb2+ leading to a higher decrease of sugar fractions as compared to the seedlings exposed to Zn2+ or Pb2+ alone.
Protein contents: Application of Zn2+ individually or in combination with Pb2+ increased protein content especially under the higher levels. While in most cases, excess supply of Pb2+ did not affect significantly insoluble and total protein while a decreased in soluble protein content was observed (Fig. 4a-c). The stimulation effect was more pronounced in seedlings exposed to Zn2+ alone than in combination with Pb2+. Heavy metal stress has been shown to induce a variety of proteins resulting in an overall increase in protein content (Shah and Dubey, 1997).
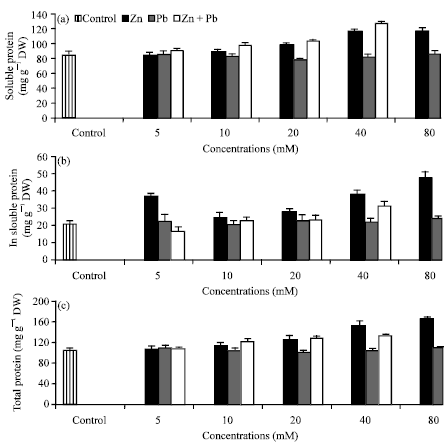 | |
| Fig. 4(a-c): | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on the contents (mg g -1 dry weight) of (a) soluble (b) insoluble and (c) total proteins of Hassawi okra seedlings. Vertical bars represent ±SD |
Our results coincide with the results obtained by John et al. (2009) in Brassica juncea L. and Hamid et al. (2010) in Phaseolus vulgari, who also reported a decrease in a soluble protein content in response to Pb2+ stress, suggesting that these heavy metals may have induced lipid peroxidation and fragmentation of proteins due to toxic effects of reactive oxygen species which led to reduced protein content. The decrease in protein content in Hassawi okra seedlings grown under Pb2+ may be due to the enhanced of protein degradation process as a result of increased protease activity (Palma et al., 2002) which is found to increase under stress conditions. While, the increased in protein content in okra seedlings exposed to Zn2+ or Pb2+alone and in combination may be due to the increase in nitrate resource in soil produced by these metals, because they were applied in nitrate form, leading to release of N in the soil and consequently increased the biosynthesis of protein, or may be due to de novo synthesis of stress proteins (Verma and Dubey, 2003; Mishra et al., 2006).
Total free amino acids and proline: The contents of total free amino acids and proline (Fig. 5a, b) were significantly increased as Zn2+ or/and Pb2+ increased while proline content was reduced at the highest (80 mM) level of Zn2+ or Pb2+. The stimulatory effect of Zn2+ individually or in combination with Pb2+ was more than Pb2+. The considerable accumulation of proline and total free amino acids as a result of heavy metals stress has been reported by Dhir et al. (2004), John et al. (2008) and Pant et al. (2011). Total free amino acids are regarded to play a significant role in metal chelation, by which heavy metal detoxification and tolerance in plants take place (Hall, 2002). Accordingly, the increase of total free amino acids in our research may be a detoxification response of Hassawi okra seedlings to Zn or/and Pb treatments.
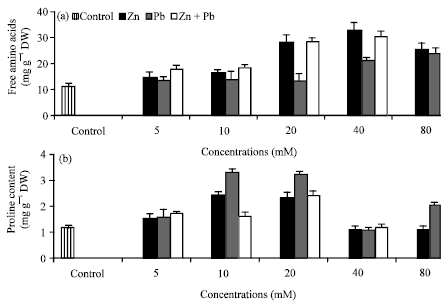 | |
| Fig. 5(a-b): | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on the contents (mg g-1 dry weight) of (a) total free amino acids and (b) proline of Hassawi okra seedlings. Vertical bars represent ±SD |
Proline was found to accumulate in Helianthus annuus due to Pb2+ and Zn2+ (Kastori et al., 1992). It suggests that physiological dryness might be an initial consequence of Zn2+ or Pb2+ stress as a result of inhibition of root growth which leads to the slowing down of the water absorption. The reduction of proline at the highest level of Zn2+ or Pb2+ may be attributed to that, these levels may be lead to either stimulation of proline degradation or inhibited its synthesis (Qureshi et al., 2005). Interestingly, our results showed that increasing of Zn2+ or Pb2+ up to 20 mM, did not alter the chlorophyll and carotenoids content. Therefore, it could be concluded that the increase in proline content explains the stability of photosynthetic pigments in Hassawi okra leaves under these levels. This results are in agreement with Backor et al. (2004) who found that proline treatment increased the content of chlorophyll and carotenoids of Trebouxia erici exposed to heavy metal stress. Further, proline contributes to stabilizing sub-cellular structures, scavenging free radicals under stress conditions (Ashraf and Foolad, 2007). This is in accordance with the alleviating effect of proline under excess of heavy metals (Mehta and Gaur, 1999). Increasing of proline content may be either due to de novo synthesis or decreased degradation or both (Kasai et al., 1998). Proline accumulation may serve as a means of osmotic adjustment and storing carbon and nitrogen when stress leads to slower growth. Thus, it is worthy to note, that the pattern of changes of total sugars was opposite to that obtained for changes in total protein, free amino acids and proline. This gives a considerable reason to believe that, heavy metals tolerance seems to be linked with an equilibrium and intereconversion between total sugars and nitrogen metabolism in Hassawi okra seedlings exposed to Zn2+ or/and Pb2+ toxicity. So, it could be concluded that, proline increases the stress tolerance of plants through such mechanisms as osmoregulation, protection of enzymes against denaturation and stabilization of protein synthesis (Schat et al., 1997).
Membrane stability index (MSI): Membrane Stability Index (MSI) which was estimated as electrolyte leakage has been used as indicator to assess tolerance of various plant species exposed to stress Sudhakar et al. (2001).
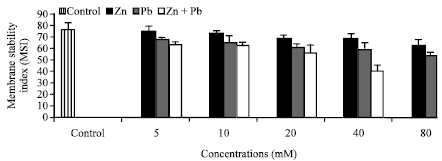 | |
| Fig. 6: | Effect of Zn2+ or/and Pb2+ treatment individually or in combination on Membrane Stability Index (MSI) of Hassawi okra seedlings. Vertical bars represent ±SD |
Linear decrease occurred in MSI (Fig. 6) with the increase of Zn2+ or/and Pb2+ in agreement with the result obtained by Zeng et al. (2011). This correlation was more notable in case of Pb2+ than Zn2+ and in binary than in individually. This is important sign of lower oxidative damage caused by ROS under Zn2+ toxicity leading to maintenance of the membrane integrity as well as reduction of membrane injury. The significant decrease in MSI at Pb2+ alone or in combination with Zn2+ may be due to plasmalemma injury caused by ROS. This supports our results which showed that, Hassawi okra seedlings exhibited higher Zn2+ tolerance than Pb2+, resulting in slighter toxicity caused by Zn2+ on their growth parameters in compared with Pb2+ when added alone or in combinations.
CONCLUSION
The results in the present work suggest that, level of 5 mM Zn2+ only had a favorable effect on growth parameters and some related physiological activities of Hassawi okra (Hibiscus esculentus L.) seedlings. While these parameters were significantly decreased as exposure to Zn2+ or Pb2+ separately or in combination. These were associated with a decreased of total sugars and increased in most cases of soluble sugar and nitrogenous compounds such as protein, total free amino acids and proline. The results obtained indicated that, Zn2+ was less toxic than Pb2+. The effect of toxicity caused by each metal alone was lower than that caused when they were used in combination. The combination effect of Zn2+ and Pb2+ produced an additive inhibitor effect on the growth parameters of Hassawi okra seedlings, suggesting synergistic or additive response between Zn2+ and Pb2+ leading to injurious effects followed by death of these seedlings, when they used in combination at 80 mM.
ACKNOWLEDGMENT
The authors are gratefully acknowledge the Deanship of the Scientific Research, King Faisal University, Saudi Arabia for funding this study (Grant No.110027).
REFERENCES
- Andras, C.D., B. Simandi, F. Orsi, C. Lambrou and D. Missopolinou-Tatala et al., 2005. Supercritical carbon dioxide extraction of okra (Hibiscus esculentus L) seeds. J. Sci. Food Agric., 85: 1415-1419.
CrossRef - Ashraf, M. and M.R. Foolad, 2007. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environ. Exp. Bot., 59: 206-216.
CrossRefDirect Link - Backor, M., D. Fahselt and C.T. Wu, 2004. Free proline content is positively correlated with copper tolerance of the lichen photobiont Trebouxia erici (Chlorophyta). Plant Sci., 167: 151-157.
CrossRefDirect Link - Bagci, S.A., H. Ekiz, A. Yilmaz and I. Cakmak, 2007. Effect of zinc deficiency and drought on grain yield of field-grown wheat cultivars in central anatolia. J. Agron. Crop Sci., 193: 198-206.
CrossRef - Bates, L.S., R.P. Waldren and I.D. Teare, 1973. Rapid determination of free proline for water-stress studies. Plant Soil, 39: 205-207.
CrossRefDirect Link - Bradford, M.M., 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72: 248-254.
CrossRefPubMedDirect Link - Clemens, S., 2006. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie, 88: 1707-1719.
CrossRefDirect Link - Dhir, B., P. Sharmila and P.P. Saradhi, 2004. Hydrophytes lack potential to exhibit cadmium stress induced enhancement in lipid peroxidation and accumulation of proline. Aquat. Toxicol., 66: 141-147.
CrossRefDirect Link - Fales, F.W., 1951. The assimilation and degradation of carbohydrate by yeast cells. J. Biol. Chem., 193: 113-124.
PubMed - Farshian, S., J. Khara and P. Malekzadeh, 2007. Effect of arbuscular mycorrhizal (G. etunicatum) fungus on antioxidant enzymes activity under zinc toxicity in lettuce plants. Pak. J. Biol. Sci., 10: 1865-1869.
CrossRefPubMedDirect Link - Garg, N. and N. Aggarwal, 2011. Effects of interactions between cadmium and lead on growth, nitrogen fixation, phytochelatin and glutathione production in mycorrhizal Cajanus cajan (L.) Millsp. J. Plant Growth Regul., 30: 286-300.
CrossRef - Genc, Y., McDonald, G.K. and R.D. Graham, 2006. Contribution of different mechanisms to zinc efficiency in bread wheat during early vegetative stage Plant Soil, 281: 353-367.
CrossRef - Guo, T.R., G.P. Zhang and Y.H. Zhang, 2007. Physiological changes in barley plants under combined toxicity of aluminum, copper and cadmium. Colloids Surf. B., 57: 182-188.
CrossRef - Hall, J.L., 2002. Cellular mechanisms for heavy metal detoxification and tolerance. J. Exp. Bot., 53: 1-11.
CrossRefDirect Link - Hamid, N., N. Bukhari and F. jawaid, 2010. Physiological responses of Phaseolus vulgaris to different lead concentrations. Pak. J. Bot., 42: 239-246.
Direct Link - Hardiman, R.T., B. Jacoby and A. Banin, 1984. Factors affecting the distribution of cadmium, copper and lead and their effect upon yield and zinc content in bush bean (Phaseolus vulgaris L.). Plant Soil, 81: 17-27.
CrossRef - Hu, H. and D. Sparks, 1991. Zinc deficiency inhibits chlorophyll synthesis and gas exchange in Stuart pecan. Hort. Sci., 26: 267-268.
Direct Link - Israr, M., A. Jewell, D. Kumar and S.V. Sahi, 2011. Interactive effects of lead, copper, nickel and zinc on growth, metal uptake and antioxidative metabolism of Sesbania drummondii. J. Hazard. Mater., 186: 1520-1526.
CrossRef - Jaleel, C.A., K. Jayakumar, Z. Chang-Xing and M. Iqbal, 2009. Low concentration of cobalt increases growth, biochemical constituents, mineral status and yield in Zea Mays. J. Sci. Res., 1: 128-137.
Direct Link - January, M.C., T.J. Cutright, H. van Keulen and R. Wei, 2008. Hydroponic phytoremediation of Cd, Cr, Ni, As and Fe: Can helianthus annuus hyperaccumulate multiple heavy metals?. Chemosphere, 70: 531-537.
CrossRef - John, R., P. Ahmad, K. Gadgil and S. Sharma, 2008. Effect of cadmium and lead on growth, biochemical parameters and uptake in Lemna polyrrhiza L. Plant Soil Environ., 54: 262-270.
Direct Link - John, R., P. Ahmad, K. Gadgil and S. Sharma, 2009. Heavy metal toxicity: Effect on plant growth, biochemical parameters and metal accumulation by Brassica juncea L. Int. J. Plant Prod., 3: 65-76.
Direct Link - Kambhampati, M.S., G.B. Begonia, M.F.T. Begonia and V. Bufford, 2005. Morphological and physiological responses of morning glory (Ipomoea lacunosa L.) grown in a lead- and chelateamended soil. Int. J. Environ. Res. Public Health, 2: 299-303.
PubMed - Kastori, R., M. Petrovic and N. Petrovic, 1992. Effect of excess lead, cadmium, copper and zinc on water relations in sunflower. J. Plant Nutr., 15: 2427-2439.
CrossRefDirect Link - Kosobrukhov, A., I. Knyazeva and V. Mudrik, 2004. Plantago major plants responses to increase content of lead in soil: Growth and photosynthesis. Plant Growth Regul., 42: 145-151.
Direct Link - Kupper, H., A. Parameswaran, B. Leitenmaier, M. Trtilek and I. Setlik, 2007. Cadmium-induced inhibition of photosynthesis and long-term acclimation to cadmium stress in the hyperaccumulator Thlaspi caerulescens. New Phytol., 175: 655-674.
CrossRef - Lee, Y.P. and T. Takahashi, 1966. An improved colorimetric determination of amino acids with the use of ninhydrin. Anal. Biochem., 14: 71-77.
CrossRefDirect Link - Lichtenthaler, H.K. and A.R. Wellburn, 1983. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochem. Soc. Trans., 11: 591-592.
CrossRefDirect Link - Marschner, H., 1995. Mineral Nutrition of Higher Plants. 2nd Edn., Elsevier, Amsterdam, Netherlands, ISBN-13: 9780080571874, Pages: 889.
Direct Link - Mehta, S.K. and J.P. Gaur, 1999. Heavy-metal-induced proline accumulation and its role in ameliorating metal toxicity in Chlorella vulgaris. New Phytol., 143: 253-259.
Direct Link - Mishra, S., S. Srivastava, R.D. Tripathi, R. Kumar, C.S. Seth and D.K. Gupta, 2006. Lead detoxification by coontail (Ceratophyllum demersum L.) involves induction of phytochelatins and antioxidant system in response to its accumulation. Chemosphere, 65: 1027-1039.
CrossRef - Monnet, F., N. Vailant, P. Vernay, A. Coudret, H. Sallanon and A. Hitmi, 2001. Relationship between PSII activity, CO2 fixation and Zn, Mn and Mg contents of Lolium perenne under zinc stress. J. Plant Physiol., 158: 1137-1144.
CrossRefDirect Link - Nyitrai, M., A.G. Szent-Gyorgyi and M.A. Geeves, 2002. A kinetic model of the co-operative binding of calcium and ADP to scallop (Argopecten irradians) heavy meromyosin. Biochem. J., 365: 19-30.
CrossRef - Palma, J.M., L.M. Sandalio, C.F. Javier, M.C. Romero-Puertas, I. McCarthy and L.A. del Rio, 2002. Plant proteases, protein degradation and oxidative stress: Role of peroxisomes. Plant Physiol. Biochem., 40: 521-530.
CrossRef - Panda, S.K., I. Chaudhury and M.H. Khan, 2003. Heavy metals induce lipid peroxidation and affect antioxidants in wheat leaves. Biol. Plant., 46: 289-294.
CrossRefDirect Link - Pant, P.P., A.K. Tripathi and V. Dwivedi, 2011. Effect of heavy metals on some biochemical parameters of Sal (Shorea robusta) seedling at nursery level, Doon Valley, India. J. Agric. Sci., 2: 45-51.
Direct Link - Patra, M., N. Bhowmik, B. Bandopadhyay and A. Sharma, 2004. Comparison of mercury, lead and arsenic with respect to genotoxic effects on plant systems and the development of genetic tolerance. Environ. Exp. Bot., 52: 199-223.
CrossRefDirect Link - Premachandra, G.S., H. Saneoka and S. Ogata, 1990. Cell membrane stability, an indicator of drought tolerance, as affected by applied nitrogen in soyabean. J. Agric. Sci., 115: 63-66.
CrossRefDirect Link - Qureshi, M.I., M. Israr, M.Z. Abdin and M. Iqbal, 2005. Responses of Artemisia annua L. to lead and salt-induced oxidative stress. Environ. Exp. Bot., 53: 185-193.
CrossRef - Ruley, A.T., N.C. Sharma, S.V. Sahi, S.R. Singh and K.S. Sajwan, 2006. Effects of lead and chelators on growth, photosynthetic activity and Pb uptake in Sesbania drummondii grown in soil. Environ. Pollut., 144: 11-18.
CrossRef - Schat, H., S.S. Sharma and R. Vooijs, 1997. Heavy metal-induced accumulation of free proline in metal-tolerant and a non-tolerant ecotype of Silene vulgaris. Physiol. Planta., 101: 477-482.
CrossRef - Shah, K. and R.S. Dubey, 1997. Effect of cadmium on proline accumulation and ribonuclease activity in rice seedlings: Role of proline as a possible enzyme protectant. Biol. Plant., 40: 121-130.
CrossRef - Sharma, P. and R.S. Dubey, 2005. Lead toxicity in plants. Braz. J. Plant Physiol., 17: 35-52.
CrossRefDirect Link - Sharma, S., P. Sharma, S.P. Datta and V. Gupta, 2009. Morphological and biochemical response of Cicer arietinum var.-pusa-256 towards an excess zinc concentration. Afric. J. Basic Applied Sci., 1: 105-109.
Direct Link - Singh, S., S. Eapen and S.F. D'Souza, 2006. Cadmium accumulation and its influence on lipid peroxidation and antioxidative system in an aquatic plant, Bacopa monnieri L. Chemosphere, 62: 233-246.
CrossRef - Sudhakar, C., A. Lakshmi and S. Giridarakumar, 2001. Changes in the antioxidant enzyme efficacy in two high yielding genotypes of mulberry (Morus alba L.) under NaCl salinity. Plant Sci., 161: 613-619.
CrossRefDirect Link - Tang, Y.T., R.L. Qiu, X.W. Zeng, R.R. Ying, F.M. Yu and X.Y. Zhou, 2009. Lead, zinc, cadmium hyperaccumulation and growth stimulation in Arabis paniculata Franch. Environ. Exp. Bot., 66: 126-134.
CrossRef - Valko, M., H. Morris and M.T.D. Cronin, 2005. Metals, toxicity and oxidative stress. Curr. Med. Chem., 12: 1161-1208.
CrossRefPubMedDirect Link - Verma, S. and R.S. Dubey, 2001. Effect of cadmium on soluble sugars and enzymes of their metabolism in rice. Biologia Plantarum, 44: 117-123.
CrossRefDirect Link - Verma, S. and R.S. Dubey, 2003. Lead toxicity induced lipid peroxidation and alters the activities of antioxidant enzyme in growing rice plants. Plant Sci., 164: 645-655.
Direct Link - Waisberg, M., P. Joseph, B. Hale and D. Beyersmann, 2003. Molecular and cellular mechanisms of cadmium carcinogenesis. Toxicology, 192: 95-117.
CrossRefDirect Link - Wierzbicka, M. and J. Obidziska, 1998. The effect of lead on seed imbibition and germination in different plant species. Plant Sci., 137: 155-171.
Direct Link - Zeid, I.M., 2001. Responses of Phaseolus vulgaris chromium and cobalt treatments. Biol. Plant., 44: 111-115.
CrossRef - Zeng, X.W., L.Q. Ma, R.L. Qiu and Y.T. Tang, 2011. Effects of Zn on plant tolerance and non-protein thiol accumulation in Zn hyperaccumulator Arabis paniculata Franch. Environ. Exp. Bot., 70: 227-232.
CrossRef - Parker, D.R., J.J. Aguilera and D.N. Thomason, 1992. Zinc-Phosphorus interactions in two cultivars of tomato (Lycopersicon esculentum L.) grown in chelator-buffered nutrient solutions. Plant Soil, 143: 163-177.
CrossRefDirect Link