
M. Kazemi
Department of Horticulture Sciences, Faculty of Agriculture and Natural Resources, Islamic Azad University Karaj Branch, Young Researchers Club, Karaj, Iran
E. Hadavi
Department of Horticulture Sciences, Faculty of Agriculture and Natural Resources, Islamic Azad University Karaj Branch, Young Researchers Club, Karaj, Iran
J. Hekmati
Department of Horticulture Sciences, Faculty of Agriculture and Natural Resources, Islamic Azad University Karaj Branch, Young Researchers Club, Karaj, Iran
American Journal of Plant Physiology
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 106-112







ABSTRACT
The effects of Salicylic acid and sucrose on the ACC-oxidase (ACO) activity, bacteria populations in vase flower preservative solution, anthocyanin leakage, Membrane stability and Malondialdehyde (MDA) content of cut flowers of carnations (Dianthus caryophyllus L. cv.White) were investigated. In this study, cut flowers of carnation were treated with Factor Salicylic acid (0,1.5, 3 mM) and sucrose (0, 3%). The vase were placed in chambers at 25°C, relative humidity about 70% and 14 h photoperiod that was maintained using fluorescent lamps (light intensity of 15 μmol m-2 sec-1) at the top of the corolla. The experimental results showed that SA treatment cause decrease malondialdehyde content and ACC-oxidase activity while the membrane stability was improved. Results also, showed that the best treatment involved 1.5 mM SA+ sucrose 3%. The vase solution containing 1.5 mM salicylic acid + sucrose 3% significantly decrease malondialdehyde content, ACC-oxidase activity and bacteria populations in vase flower preservative solution while increase the vase life and Membrane stability of carnation cut flower compared to the control. Results suggest that salicylic acid and sucrose increases Membrane stability by decrease malondialdehyde content, ACC-oxidase activity and bacteria populations in vase flower preservative solution of the carnation cut flowers. This study shows that Salicylic acid and sucrose treatment did show significant effect on quality parameters and carnation flower longevity. Salicylic acid and sucrose proved more effective in delaying petal senescence and/or flower wilting. However, our result showed that Salicylic acid and sucrose treatments maintained the vase life of flowers for a longer period.
PDF Abstract XML References Citation
Received: January 10, 2011;
Accepted: April 11, 2011;
Published: June 03, 2011
How to cite this article
M. Kazemi, E. Hadavi and J. Hekmati, 2011. Role of Salicylic Acid in Decreases of Membrane Senescence in Cut Carnation Flowers. American Journal of Plant Physiology, 6: 106-112.
DOI: 10.3923/ajpp.2011.106.112
URL: https://scialert.net/abstract/?doi=ajpp.2011.106.112
DOI: 10.3923/ajpp.2011.106.112
URL: https://scialert.net/abstract/?doi=ajpp.2011.106.112
INTRODUCTION
Carnation is one of the world's most popular flowers to produce cut flowers. The flower of Carnation is highly ethylene sensitive and the longevity of the cut flower is very short (Brandt and Woodson, 1992). Short postharvest vase life is one of the most important problems on the cut flowers. In general, the senescence of ethylene-sensitive flowers, such as carnations, is associated with a loss of membrane integrity, climacteric rise of respiration and enchanced ethylene synthesis (Tang et al., 1994; Da Silva, 2003). Some workers have shown that reduced vase life is associated with increased concentration of microbes in the vase solution (Burdett, 1970; Dansercau and Vines, 1975) the media used to determine microbial concentrations in vase solutions may not have been suitable for growth of microbial taxa adversely affecting flower vase life, also Kazemi et al. (2010) found that some bacteria from vase water produced ethylene. In carnations, senescence of the petals is associated with a climacteric-like increase in ethylene production during the final stages (Brandt and Woodson, 1992). Kazemi et al. (2010) have noted a strong correlation between membrane leakiness and phospholipid breakdown in senescing flowers. Ethylene accelerates the onset of membrane leakiness and phospholipid deterioration in petals. Senescing carnation flowers exhibit a climacteric-like rise in ethylene production. Ethylene production increases sharply with senescence while exogenous application of ethylene enhanced flower senescence and wilting, increased permeability of petal cells and accelerated the decrease in cell membrane fluidity (Reezi et al., 2009) and increase production of ROS. Reactive Oxygen Species (ROS) cause chlorophyll degradation and membrane lipid per oxidation and increase Malondialdehyde (MDA) product. To scavenge ROS, plants posses specific mechanisms which include activation of antioxidant enzymes (Jaleel et al., 2006) and non-enzymatic antioxidants such as, carotenoids , ascorbic acid and Phenolic compounds (Mittler, 2002). The effects of ethylene and ROS can be reduced by inhibitors of ethylene biosynthesis and increase enzyme antioxidant activity. SA is a well known phenol that can prevent ACC-oxidase activity that is the direct precursor of ethylene and decrease ROS with increase enzyme antioxidant activity (Ansari and Misra, 2007; Mba et al., 2007; Mahdavian et al., 2007; Canakci, 2008), also Salicylic acid seems to act by germicide the decrease of bacteria which block the xylem vessels in the cut region and interfere with the normal flux of water through the stem (Nowak and Rudniki, 1990). Fan et al. (2008) showed that SA can extending the vase life of cut flowers with decrease ROS and ethylene. Salicylic Acid (SA) acid with increases the enzyme antioxidant activity cause delay the onset of hydrolysis of structural cell components, decrease ROS production, ACC-oxidase activity and sensitivity. SA acid decreased the permeability of plasma membrane of floret cells and improved the structure of chloroplasts which were badly damaged by ethylene. Other important factor in the deterioration of cut flowers involves the diminishing of respiration substrates, the speed of these changes depend, at least in part, on the amount of reserves that are present in the flower when they are cut (Han, 2003). Therefore, an exogenous carbohydrate supplementation would be enough to delay the senescence, considering that the main effect would be to maintain the structure and activity of the mitochondria. The postharvest life of flowers is strongly dependent on the carbohydrate status and the acceptable amount of metabolic sugars are factors that affect the rate of senescence. Keeping the flower in vase solutions containing sucrose has been shown to extend their vase-life (Kazemi et al., 2010). This research was designed to investigate the role of SA in alleviating membrane lipid per oxidation and MDA content in Cut flowers of Dianthus caryophyllus.
MATERIALS AND METHODS
Plant material: Carnations (Dianthus caryophyllus cv. White) were grown in the greenhouse standard production methods (pakdasht, Tehran, Iran). The experiment was started on February 15, 2010 and chlorophyll content, Membrane stability, MDA content and ACC oxidase activity were measured at 11th day of vase life. The experimental site was in horticulture laboratory of agriculture faculty of University Azad Karaj, Tehran, Iran. Flowers were weighed initially immediately after harvest and used for setting treatments. The factors were three levels of salicylic acid (0, 1.5, 3 mM) and two levels of sucrose (0.3%). The flowers were individually placed in bottles containing 250 mL of preservative solution and were held at ambient temperature (19±5°C).
Determination of anthocyanin leakage and ACC oxidase activity: Anthocyanin leakage was used to assess membrane permeability and measured using spectrophotometer. The procedure used was based on the method of Poovaiah (1979). ACC oxidase activity was determined according to the method described by Moya-Leon and John (1994).
Assays of MDA content (Lipid per oxidation): Oxidative damage to lipids was estimated by measuring the content of MDA in floret segment homogenates, prepared in 10% trichloroacetic acid containing 0.65% 2-Thiobarbituric Acid (TBA) and heated at 95°C for 25 min, as described by Heath and Packer (1968). MDA content was calculated by correcting for compounds other than MDA which absorb at 532 nm by subtracting the absorbance at 532 nm of a solution containing plant extract incubated without TBA from an identical solution containing TBA.
Chlorophyll (a + b) content measurement: Chlorophyll total (a + b) content was measured by Chlorophyll meter SPAD-502, Minolta Co. Japan which represented by SPAD value. The petal was inserted into the meter and measured SPAD value 3 times from different spot of a single leaves.
Microbe population: Test Microbe population were isolated from vase solutions of carnations. When the flowers had senesced (about 11 days), aliquots of the vase solutions were diluted 100-times and 25 μL aliquots of the diluted solution were spread on sterile nutrient agar, in sterile petri plates. The plates were allowed to incubate for 48 h at room temperature and individual colonies of microorganisms, representing the most common colony morphology types, then were picked off the agar media with a sterile loop and streaked on EMB medium for purification. Purified microbe population were maintained axenically on EMB medium and transferred daily to fresh medium.
Vase life: Vase life was considered to be terminated when wilting occurred.
Experimental design and statistical analysis: Experiment was arranged in a factorial test with complete randomized design with four replications. Analysis of variance was performed on the data collected using the General Linear Model (GLM) procedure of the SPSS software) Version 16, IBM Inc.). The mean separation was conducted by Tukey analysis in the same software (p = 0.05).
RESULTS AND DISCUSSION
Anthocyanin leakage and Acc-oxidase activity: Table 1 and 3 showed the effects of SA on the anthocyanin leakage of the petals of carnation under SA and SA+ SU treatment compared to the control. Two treatments (SA 1.5 mM and SA + SU (SA1.5 + SU3%) improved membrane permeability by the decrease of anthocyanin leakage and ACC-oxidase activity (Table 1, 3) while, two treatments (3 mM and 3 mM + SU 3%) impaired membrane permeability by increasing anthocyanin leakage and increase ACC-oxidase activity (Table 1, 3) (p≤0.05). Result showed that addition of 1.5 mM SA maintained membrane permeability, 1.5 mM SA with or without SU could alleviate or decrease cell wall damages (Table 3). It is evident from the data presented in Table 1 that the maximum anthocyanin leakage was recorded in 3 mM SA compare another treatments and control (p≤0.05). Present results showed that adding SA in vase solution can prevent anthocyanin leakage by maintained PH vase solution. Results also showed that adding SA was found to be positively correlated with anthocyanin leakage of the carnation cut flower (Table 2) (p≤0.01).
| Table 1: | Mean comparisons of chlorophyll content, vase life, MDA, bacteria populations in vase flower preservative solution, antocyanin leakage and ACC oxidase activity in SA treatment |
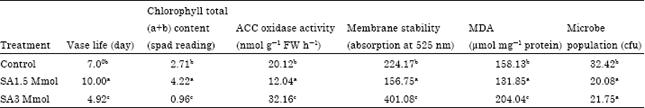 | |
| Means in each column followed by similar letters are not significantly different at 5% level using Duncan multiple rang test | |
| Table 2: | Simple correlation lines between the SA treatment with other variables |
 | |
| *, **: Significant different at 5 and 1% level, respectively | |
| Table 3: | Mean comparisons of chlorophyll content, Vase life, Antocyanin leakage, MDA content, bacteria populations in vase flower preservative solution and ACC Oxidase Activity in SA*SU treatment |
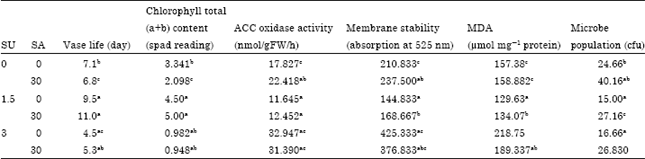 | |
| Means in each column followed by similar letters are not significantly different at 5% level using Duncan multiple rang test | |
The produce ethylene is in petal cut flower harmful to cut flowers and their consequent accelerate flower senescence, anthocyanin leakage and cell death. Adding a suitable inhibit produce ethylene in vase solution can prevent accelerate flower senescence, anthocyanin leakage and cell death. Present results are in agreement with previous scientists who have reported decreased accelerate flower senescence, anthocyanin leakage and cell death of cut flowers when placed in solutions of a suitable inhibit produce ethylene (Fan et al., 2008). The protective function of SA includes the regulation of ROS and antioxidant enzymes (Khan et al., 2003; El-Tayeb et al., 2006; Shi and Zhu, 2008; Joseph et al., 2010). Fan et al. (2008) showed that SA can extending the vase life of cut flowers with increase Membrane stability. The data reported here provide good evidence that SA had an additive effect in decreased accelerate flower senescence, anthocyanin leakage and cell death. It seems that in high concentrations of SA, pH decreased and affected vacuoles pH and resulted Anthocyanin leakage (Table 1). Table 1 and 3 demonstrate that Acc-oxidase activity stimulates anthocyanin leakage from petals. Results indicating that Acc-oxidase activity increases membrane permeability in an unspecific fashion. Therefore, since anthocyanins are localized within the vacuole of all cells, result our showed that the action of ethylene in enhancing the rate of leakage of this pigment can be interpreted as an effect of the gas on senescence and the membrane permeability of cut flower. SA in suitable concentration (1.5 mM) with sucrose can prevent negative effects low PH that cause Anthocyanin leakage. Unfortunately, High Salicylic acid concentrations rapidly increase Anthocyanin leakage, cell death and accelerate flower senescence.
Lipid per oxidation, MDA content and chlorophyll content: The effects of SA on the data for Malondialdehyde (MDA) content are presented in Table 1 and 3 (p≤0.05). Under the effect of 3 mM SA treatment MDA content increased significantly in compared to control (Table 1) (p≤0.05). Salicylic acid 1.5 mM and salicylic acid 1.5 mM+sucrose 3% significantly reduced MDA content of cut flower in all treatments except than 3 mM SA and SA+SU (SA 3 mM + SU 3%) treatment compare to other treatment (Table 3) (p≤0.05), addition of SA at level 1.5 mM decrease produces this factor and could show better results compare to alone sucrose treatment and high SA concentration. Results showed adding SA was found to be positively correlated with Lipid per oxidation of the carnation cut flower (Table 2) (p≤0.01). This indicates that with SA concentration increased, the Lipid per oxidation was increased. Cut flower under senescence is in good correlation with increased lipid per oxidation levels and this result is in good correlation with the increase in MDA content under senescence (Shakirova, 2007). A lower lipid per oxidation resulting from elevated activities of antioxidants was also reported on of cut gerbera jamesonii flower (Yuping, 2009). The results of the present experiment were similar with the findings of Fan et al. (2008) in Gerbera which showed that added SA decreased the permeability of the plasma membrane of petal cells and decreased MDA level. It was reported that SA enhanced the stability of lipids in cell membranes of Gerbera when cut flower exposed to vase solution having SA. The evidence suggests that SA decreases the permeability of plasma membranes and membrane lipid per oxidation and maintains the membrane integrity. In this study, Two treatments (3 mM SA and 3 mM SA + SU 3%) decreased chlorophyll content in carnation cut flower but SA at 1.5 mM could prevent decrease chlorophyll content in carnation cut flower (Table 1). Canakci (2008) reported that treatment with salicylic acid significantly extends the vase life with increases total chlorophyll content.
Microbe population: The Microbe population vase solution of carnation cut flowers was decreased by the concentration of salicylic acid 1.5 and 3 mM used (Table 1). The Microbe population was lower in salicylic acid at 1.5 and 3 mM compared to salicylic combined with sucrose treatment and control (Table 3) (p≤0.05). The higher Microbe population was attained when sucrose was use comparison to control (Table 3). Means of Microbe population on vase solution of cut flowers in various salicylic acid + sucrose containing vase solutions was slightly significantly than control while means of microbe population on vase solution of cut flowers in various salicylic acid containing vase solutions was higher significantly than control. Results showed adding SA was found to be negatively correlated with microbe population vase solution of the carnation cut flower (Table 2) (p≤0.01). This indicates that with SA concentration increased, the microbe population vase solution was decreased.
Vase life: Table 1 showed that SA alone was capable of increasing longer vase life of carnation flowers than sucrose alone or together with SA (Table 1, 3) (p≤0.05). It is evident from the data presented in Table 3 that the maximum vase-life (10, 11 days ) were recorded in SA at 1.5 mM and Salicylic acid 1.5 mM+sucrose 3% respective compare other treatments and control. The minimum vase-life was noted in SA at 3mM compare other treatments and control (Table 3) (p≤0.05). Fan et al. (2008) showed that the treatment of salicylic acid extended the vase life and improved flower quality with reduced respiration rate delay senescence and decrease Lipid per oxidation, MDA content. Fan et al. (2008) showed that the treatment of salicylic acid extended the vase life and improved flower quality with reduced respiration rate delay senescence and decrease Lipid per oxidation, MDA content. Ichimura and Hiraya (1999) reported that treatment with sucrose extends the vase life of florets harvested at a bud stage. Keeping the flowers in vase solutions containing sucrose has been shown to extend their vase-life (Han, 2003; Yamane et al., 2005). Present results showed Adding SA was found to be positively correlated with vase life of the carnation cut flower (Table 2) (p≤0.01). The correlation was significant. This indicates that with SA concentration increased, the vase-life was decreased.
CONCLUSION
This study shows that Salicylic acid and sucrose treatment did show significant effect on quality parameters and rose flower longevity. Salicylic acid and sucrose proved more effective in delaying petal senescence and/or flower wilting. However, our result showed that Salicylic acid and sucrose treatments maintained the vase life of flowers for a longer period. Further studies are required, especially to validate some antioxidant enzyme activity and antioxidant capacity.
REFERENCES
- Ansari, M.S. and N. Misra, 2007. Miraculous role of salicylic acid in plant and animal system. Am. J. Plant Physiol., 2: 51-58.
CrossRefDirect Link - Canakci, S., 2008. Effects of salicylic acid on fresh weight change, chlorophyll and protein amounts of radish (Raphanus sativus L.) seedlings. J. Biol. Sci., 8: 431-435.
CrossRefDirect Link - da Silva, J.A.T., 2003. The cut flower: Postharvest considerations. J. Biol. Sci., 3: 406-442.
CrossRefDirect Link - El-Tayeb, M.A., A.E. El-Enany and N.I. Ahmed, 2006. Salicylic acid-induced adaptive response to copper stress in sunflower (Helianthus annuus L.). Int. J. Bot., 2: 372-379.
Direct Link - Han, S.S., 2003. Role of sugar in the vase solution on postharvest flower and leaf quality of oriental lily stargazer. Hort. Sci., 38: 412-416.
Direct Link - Ichimura, K. and T. Hiraya, 1999. Effect of silver thiosulfate complex (STS) in combination with sucrose on the vase life of cut sweet pea flowers. Hort. Sci., 68: 23-27.
Direct Link - Joseph, B., D. Jini and S. Sujatha, 2010. Insight into the role of exogenous salicylic acid on plants grown under salt environment. Asian J. Crop Sci., 2: 226-235.
CrossRefDirect Link - Khan, W., B. Prithiviraj and D.L. Smith, 2003. Photosynthetic responses of corn and soybean to foliar application of salicylates. J. Plant Physiol., 160: 485-492.
CrossRefDirect Link - Mahdavian, K., K.M. Kalantari and M. Ghorbanli, 2007. The effect of different concentrations of salicylic acid on protective enzyme activities of pepper (Capsicum annuum L.) plants. Pak. J. Biol. Sci., 10: 3162-3165.
CrossRefPubMedDirect Link - Mba, F.O., X. Zhi-Ting and Q. Hai-Jie, 2007. Salicylic acid alleviates the cadmium toxicity in Chinese cabbages (Brassica chinensis). Pak. J. Biol. Sci., 10: 3065-3071.
CrossRefPubMedDirect Link - Mittler, R., 2002. Oxidative stress, antioxidants and stress tolerance. Trends Plant Sci., 7: 405-410.
CrossRefPubMedDirect Link - Reezi, S., M. Babalar and S. Kalantari, 2009. Silicon alleviates salt stress, decreases malondialdehyde content and affects petal color of salt stressed cut rose (Rosa xhybrida L.) Hot Lady. Afr. J. Biotechnol., 8: 1502-1508.
Direct Link - Shi, Q. and Z. Zhu, 2008. Effects of exogenous salicylic acid on manganese toxicity, element contents and antioxidative system in cucumber. Environ. Exp. Bot., 63: 317-326.
CrossRefDirect Link - Tang, X., A. Gome, A. Bhatia and W. William, 1994. Pistil-specific and ethylene-regulated ex-pression of 1-aminocyclopropane-1-carboxylate oxidase genes in petunia flowers. Plant Cell, 6: 1227-1239.
Direct Link - Yamane, K., T. Kawauchi, Y.T. Yamaki and N. Fujishige, 2005. Effects of treatment with trehalose and sucrose on sugar contents, ion leakage and senescence of florets in cut gladiolus spikes. Acta. Hort., 669: 351-358.
Direct Link - Brandt, A.S. and W.R. Woodson, 1992. Variation in flower senescence and ethylene biosynthesis among carnations. HortScience, 27: 1100-1102.
Direct Link - Jaleel, C.A., R. Gopi, G.M.A. Lakshmanan and R. Panneerselvam, 2006. Triadimefon induced changes in the antioxidant metabolism and ajmalicine production in Catharanthus roseus (L.) G. Don. Plant Sci., 171: 271-276.
CrossRefDirect Link - Heath, R.L. and L. Packer, 1968. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys., 125: 189-198.
CrossRefPubMedDirect Link