
Abdoulaye Coulibaly
Jiangnan University, School of Food Science and Technology, Wuxi, China
Brou Kouakou
Universite d`Abobo-Adjame, UFR-STA, Abidjan, Cote d`Ivoire
Jie Chen
Jiangnan University, School of Food Science and Technology, Wuxi, China
American Journal of Plant Nutrition and Fertilization Technology
Year: 2011 | Volume: 1 | Issue: 1 | Page No.: 1-22







ABSTRACT
Cereals are a major source of micronutrients, the bioavailability of minerals such as iron, zinc, calcium, magnesium, is low because they are present as insoluble complex with food components such as phytic acid. After several readings it should be remembered that the phytic acid in whole is seen as an anti-nutritional factor than therapeutic element. The elimination of phytic acid can be done in several ways and should not be total if we want to minimize losses of dry matter and minerals.
PDF Abstract XML References Citation
Received: March 07, 2011;
Accepted: April 22, 2011;
Published: June 03, 2011
How to cite this article
Abdoulaye Coulibaly, Brou Kouakou and Jie Chen, 2011. Phytic Acid in Cereal Grains: Structure, Healthy or Harmful Ways to Reduce Phytic Acid in Cereal Grains and Their Effects on Nutritional Quality. American Journal of Plant Nutrition and Fertilization Technology, 1: 1-22.
DOI: 10.3923/ajpnft.2011.1.22
URL: https://scialert.net/abstract/?doi=ajpnft.2011.1.22
DOI: 10.3923/ajpnft.2011.1.22
URL: https://scialert.net/abstract/?doi=ajpnft.2011.1.22
INTRODUCTION
Phytic acid (C6H18O24P6) also known as inositol hexaphosphate (IP6) or phytate as a salt, is the storage form of Phosphorous in all grains and oil seeds (Jacela et al., 2010). It accounts for 50-80% of the total phosphorus in different cereals. The amount of phytate in grains, nuts, legumes and seeds is highly variable; the levels that researchers find when they analyze a specific food probably depends on growing conditions, harvesting techniques, processing methods, testing methods and even the age of the food being tested. Phytic acid will be much higher in foods grown using modern high-phosphate fertilizers than those grown in natural compost (Srivastava et al., 1955). The accumulation site of phytic acid in monocotyledonous seeds (wheat, millet, barley, rice, etc.) is the aleurone layer, particularly the aleurone grain. Corn differs from other cereals as more than 80% of phytic acid is concentrated in germ. Phytic acid content of cereals varies from 0.5 to 2.0%. Phytate is most known as a substance known to decrease mineral absorption however, it has also been looked at as a possible beneficial vitamin-like substance (Okazaki and Katayama, 2005). Phytic acid has a strong ability to chelate multivalent metal ions, specially zinc, calcium, iron and as with protein residue. The binding can result in very insoluble salts with poor bioavailability of minerals (Zhou and Erdman, 1995). The association of phytate with proteins begins in seeds during ripening, when phytate accumulates in the protein-rich aleurone layer of cereals. However, phytate-phosphorus is less nutritionally available since the phytate is not quantitatively hydrolyzable in human gut (Sandberg and Andersson, 1998). Many methods of phytic acid determination have been developed. Sandberg (1995) had reported the precipitation and ion-exchange method's are not specific as they do not separate inositol hexaphosphate from lower inositol phosphates and thus overestimate the phytate content in processed foods. The HPLC method determines the inositols in processed foods. The phytic acid in unprocessed products mainly appears as inositol hexaphosphate (IP6); since the precipitation methods are useful to measure the phytic acid content in unprocessed products.
Generally cereal grains contain lots of carbohydrates, about 70 to 80%, as starch, protein (until 15%); lipids in small proportions (less than 5%), minerals and vitamins. The Grains of great interest due to their good acceptability on the organoleptic properties and low cost makes them accessible to the most underprivileged populations. However, their nutritional quality about it remains insufficient. In addition, micronutrients found there are not only insufficient to cover the nutritional needs but also their bioavailability is low because of anti-nutritional factors (as phytic acid). Several pre-treatments exist to improve the quality of cereal. Nout wrote that it is the simple traditional household technologies have been used to process the cereal in order to improve the nutritional quality (Nout, 1993).
The goal of this study was to know how phytic acid is formed in the seed, its relationship with our health, by what ways can we reduce its quantity in cereals before consumption and the effects of those ways on nutritional quality.
SOURCE, STRUCTURE AND CONNECTIONS
Phytic acid is present in beans, seeds, nuts, grains-especially in the bran or outer hull; phytates are also found in tubers and trace amounts occur in certain fruits and vegetables like berries and green beans. Normally the salts of phytic acid are found in plant seeds, animals and soils but the acid originates from natural mineral sources containing phosphorus or from fertilizers (Marchner, 1997). As P containing fertilizers are applied to the soils, plant roots pick up the P at a physiological pH mainly as PO-34 which remains as inorganic Phosphorus (P) and is esterified through the I hydroxyl group to the carbon chain (C-O-P) as a single phosphate ester or attached to another phosphate by an energy rich pyrophosphate bond. The rate of exchange between P and organic P in the ester and the pyrophosphate bond is very high and this leads to plant roots incorporating P into the organic P within a few 1 min but this is released almost immediately again into the xylem. The amount of phytate present in plant seeds and grains ranges from 0.5-5% (Loewus, 2002). It is mainly present as a salt of mono-valent and divalent cations such as K+, Mg2+ and Ca2+. It is accumulated into the seed during the ripening period. In dormant seeds, phytate represents 60-90% of total phosphate. Phytate is a naturally occurring compound formed during maturation of plant seeds and grains and therefore a common constituent of plant derived food. The alkali metal (Na, K) salts of phytic acid are usually soluble but those formed with alkali earth and transition metals are practically insoluble. During the development of seeds especially cereals the cells become filled with starch grains and protein bodies known as aleouron which contain phytic acid that accumulates as the seed develops. During germination there is marked increase in the activity of various phosphate uses including phytase. The activity of phytate in cereals is highest in scutulum and aleouron layers which results in the liberation of considerable amount of organic phosphate, calcium, magnesium and potassium for general metabolic processes of the plant. However, it has been stated that leafy vegetables and fruits do not contain any phytic acid; Seeds and bran are the highest sources of phytates, containing as much as two to five times more phytate than even some varieties of soybeans which we know are highly indigestible.
The phytic acid being the principal storage form of phosphorus in many seeds is named myo-inositol hexaphosphoric acid, IP6. Its molecular formula is C6H18O24P6 and its molecular weight is 660.03. The structure of phytic acid is shown in Fig. 1 and then Fig. 2 shows the structure of phytic acid with the different possibilities to interact with both metal cations (minerals) as with protein residues.
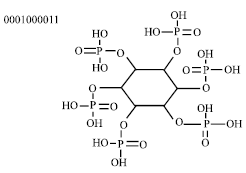 |
| Fig. 1: | Structure of phytic acid |
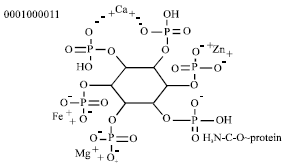 |
| Fig. 2: | Structure of phytic acid with the different possibilities to interact with both metal cations (minerals) as with protein residues |
It contains the mineral phosphorus tightly bound in a snowflake-like molecule. In humans and animals with one stomach, the phosphorus is not readily bioavailable. In addition to blocking phosphorus availability, the arms of the phytic acid molecule readily bind with other minerals, such as calcium, magnesium, iron and zinc, making them unavailable as well. Phytic acid molecule contains 12 dissociable hydrogens. Lasztity (1986) and Lasztity and Lasztity (1988) found that, depending on the pH of the solution different phytic acid anions may be formed having different degree of protonation. Except at the pH corresponding to the isoelectric point, protein molecules are also charged. The terminal amino groups like lysyl, histidyl and arginyl can be positively charged at a low pH below the isoelectric point of proteins; any of these groups can directly form a complex with a negatively charged phytate anion. One phytate anion can interact with two charged groups of protein in the normal steric condition. According to the number of positively charged groups and conformational conditions, the protein molecule can bind more phytate anions at the same time. Also at intermediate pH values, the lysyl and arginyl groups are only positively charged, so in this case a slight possibility of electrostatic interactions exists between these groups and phytate anions. The interaction between phytic acid and protein is reduced when the pH is very high. It should also be noted that the ternary complexes can be formed when the polyvalent cations are present. In this case the cation forms a bond between the phytate anion and a negatively charged group of the protein. Tompson (1986) showed that ternary complexes of protein, phytic acid and carbohydrate might form; the digestion rate of starch is affected in this case. In addition to being an element chelator of important minerals, phytic acid also inhibits the enzymes that we need to digest our food, namely pepsin, amylase and trypsin which are, respectively necessary for protein degradation in stomach, the starch into sugars and proteins degradation in the small intestine.
Phytate
Healthy or harmful: Phytic acid is considered as an anti-nutritient, it forms insoluble complexes with minerals such as zinc, calcium, magnesium and iron. During the pre-weaning period, the diet of an infant is based upon cereal flours as well as upon baby milk formulas. Therefore, the negative properties of phytates, particularly on the bioavalability of minerals, may have an effect on health during this first period. All kinds of cereal grains and other products, such as roots (cassava) and seeds (peanuts, soy) are used to prepare the cereals for children. The infant, after 4 months of age, is at risk of developing an iron deficit as a result of a decrease of his organic deposits and the increase of his needs, determined by the higher growth velocity. This shortage may also be influenced by the presence of phytates in these cereal derivatives. Cereal porridges are common complementary foods during the weaning period and often provide much of the dietary iron intake because the iron contribution from human milk is low. Both cereal grains and legume seeds are rich in phytic acid. Iron absorption of native iron and fortification iron may be very low because of the high phytate content of cereal porridges; this was reported by Cook et al. (1997) and Hurrell et al. (1992). Also Navert et al. (1985) reported that we will absorb approximately 20% more zinc and 60% magnesium from our food when phytate is absent. In the same vein (Hurrell, 2003) reported the iron absorption rate in the wheat porridge increased to 12% when the phytic acid was removed. Another set of experiments examined depression-fighting zinc and magnesium. Researchers provided two groups of people with bread. One group with bread with phytates and one with a control bread without phytates. Researchers then studied participants’ mineral absorption via stool samples. Without phytic acid, participants absorbed about 30% of magnesium and zinc. With phytic acid, participants absorbed only 13% of their magnesium and 23% of their zinc (Egli et al., 2002; Bohn et al., 2004). The zinc-and iron-blocking effects of phytic acid can be just as serious as the calcium-blocking effects (Hallberg et al., 1989) showed that a wheat roll containing 2 mg phytic acid inhibited zinc absorption by 18%; 25 mg phytic acid in the roll inhibited zinc absorption by 64% and 250 mg inhibited zinc absorption by 82%. The growing children run into severe problems in a phytate-rich diet, their bodies will suffer from the lack of calcium and phosphorus with poor bone growth, short stature, rickets, narrow jaws and tooth decay and for the lack of zinc and iron with anemia and mental retardation. Hurrell et al. (1998) showed that iron absorption in infants is inhibited by phytic acid in a way similar to iron absorption in adults.
Although there are many adverse effects of high intake of phytate in humans, many beneficial effects have also been found. Phytate exerts beneficial effects in the gastrointestinal tract and other target tissues through its chelating ability. Wise and Gilbert (1981) showed that phytate as a calcium salt can protect against dietary Pb2+ in experimental animals and human volunteers, hence phytate has the ability to counteract acute oral Pb2+ toxicity. Phytic acid has a structure similar to that of myo-inositol which has been demonstrated to reduce hepatic lipid levels. Onomi et al. (2004) found that phytic acid at a level of 0.035% may protect against a fatty liver resulting from elevated hepatic lipogenesis and that the anti-nutrient effect of phytic acid on mineral absorption will only occur at 10 fold higher levels. As chelator phytates bind to extra iron or toxic minerals and remove them from the body. As with all anti-nutrients, phytates may play a therapeutic role in certain cases. Heart disease is a leading cause of death in the Western countries but it is low in Japan and developing countries. Elevated plasma cholesterol or elevated LDL-cholesterol concentrations have been shown to be one of the risk factors. It has been suggested that dietary fibre or more specifically phytate which is a component of fibre can influence the aetiology of heart disease (Potter, 1995). Jariwalla et al. (1990) showed that dietary phytate supplementation resulted in the lowering of serum cholesterol and triglyceride levels. This effect accompanied the decrease in serum Zn level and Zn-Cu ratio. This is because coronary heart disease appears to be caused by an imbalance of Zn-Cu metabolism. Incidence of cancer especially intestinal cancer has been associated with dietary fat intake which is inversely related to fibre intake. The rate of colonic cancer can arise from many fibre-rich foods which may contain high phytate. Lima-Filho et al. (2004) through their research have shown that an enzymatic method based on the ability of urease to be inhibited by very low concentrations of copper ion was utilized as technique to study the protective effect of phytic acid against copper ions. They found that copper (0.79 and 1.57 μM) promoted a 43.82±2.72 and 71.84±6.29% inhibition of the enzyme activity, respectively; enzyme inhibition by the same copper concentrations in the presence of phytic acid (10 mM) were slightly lower. That would mean, phytic acid has a copper chelating capacity that could be useful in preventing copper interaction with biomolecules and, accordingly, in lowering the generation of ROS. Conversely high phytate intake can be a factor in reducing breast and prostate cancer in man (Vucenik and Shamsuddin, 2003). Cebrian et al. (2007) demonstrated that phytic acid is one of few chelating therapies used for uranium removal.
The use of phytic acid to eliminate the minerals excess may deprive other cells in the body that need it. For example, the removal of iron by phytates has an effect on red blood cells. Phytic acid is chelating effect may serve to prevent, inhibit or even cure some cancers by depriving those cells of the minerals (iron) they need to reproduce.
Phytic acid threshold: It appears that once the phytate level has been reduced, such that there is more available phosphorus than phytate in the grain, the food becomes more beneficial than harmful. Phytates should be lowered as much as possible for best health, ideally to 25 mg or less per 100 g or to about 0.035% of the phytate containing food eaten (Onomi et al., 2004). At this level, micronutrient losses are minimized. The Table 1 and 2 shows the phytate content of common foods as a percentage of dry weight (Reddy and Sathe, 2001).
| Table 1: | Phytate content of common foods as a percentage of dry weight |
 | |
| Table 2: | Bread phytate as percentage of weight |
 | |
An excess of 800 mg phytic acid per day is probably not a good idea, it should be noted that the Recommended Daily Intake (RDI) of phytic acid varies from one country to another. The average phytate intake in the United State and the United Kingdom ranges between 631 and 746 mg day-1; the average in Finland is 370 mg in Italy it is 219 mg and in Sweden a mere 180 mg per day.
Depending on age, physiological state and needs of each individual, the amount of phytic acid to consume differs. For example, children under six, pregnant women or people with a certain disease should eat a diet low in phytic acid.
WAYS TO REDUCE OR NEUTRALIZE PHYTIC ACID IN CEREAL GRAINS
Above we reported that, Phytic acid in grains represents a serious problem in our diets. Phytic acid blocks absorption of not only of phosphorus but also other minerals such as calcium, magnesium, iron and zinc. It also negatively affects the absorption of lipids and protein because phytic acid inhibits enzymes that we need to digest our food such as pepsin (which helps break down protein), amylases (convert starch into sugar for digestion) and trypsin (also used in protein digestion). But the question is, how do we effectively reduce phytic acid?
Germination or sprouting: The first step of the process of germination is soaking and is usually done at room temperature in the tropics. The soaking helps to bring the grain moisture at a value favourable to respiratory and metabolic activities and the mobilization of primary and secondary metabolites, thereby allowing germination. It also provides additional cleaning. Cereal grains are sometimes treated before and/or after soaking with chemicals such as formaldehyde 0.2% (Elmaki et al., 1999), sodium hypochlorite 1% available chlorine (Ogbonna et al., 2004), ethanol 70% (Mbofung and Fombang, 2003) to prevent mould growth. This product reduces germinative power of seeds, except formaldehyde but poses a health problem for the user. We must do this treatment before soaking and washing should be especially grains several times until complete elimination of the odour of the chemical to avoid the health problems for the user. One can also use lemon extract to 0.1% as described by Malleshi and Desikachar (1986).
Environmental factors (water, oxygen, light and temperature) play an important role in the germination. Water hydrates envelopes the embryo, a good aeration of the medium is a prerequisite for germination. The temperature changes the percentage and speed germination. Many seeds germinate only after exposure to light. The seed hydrated at a suitable temperature swells. The seed coat bursts and the embryo breathes actively develops. The germination of millet was not different from that of other cereals. The process for obtaining seeds germinated millet is described by Kouakou et al. (2008) in Fig. 3.
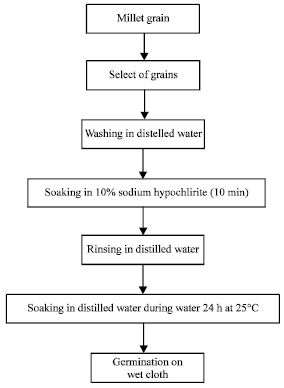 |
| Fig. 3: | Flow chart illustrating preparation of germinated millet |
Significant amounts of phytic acid will be eliminated in most of sprouted grain products. Many studies have shown, malting millet resulted in a decrease of 23.9% phytic acid after 72 h and 45.3% after 96 h (Makokha et al., 2002). With increased germination periods at 30°C, phytic acid removal progressed from 4 to 60% (Liang et al., 2008). Poiana et al. (2009) found also that the greatest reduction in phytate phosphorus was seen in germinated rye seeds while the smallest decrease was found for maize seeds. Konietzny et al. (1994) and Greiner et al. (2000) observed maximum phytase activity in espelt and barley on the 5th and 4th days of germination, respectively. It is the same for Agostini et al. (2010) who found that phytase activity peaked on the fourth day of germination and in contrast, the lowest amount of phytic acid was reached approximately on the same day of germination, indicating the hydrolysis of phytic acid by this enzyme. Other studies have proposed that the activity of endogenous phytase was the main factor lessading to a reduction of phytic acid during soaking (Lestienne et al., 2005a, c, d). Sprouting oats for five days at 52°F and then soaking for 17 h at 120°F removes 98% of phytates, adding malted rye further enhances oat phytate reduction (Larsson and Sandberg, 1992). Four pearl millets were germinated over a six day period and tested to see what the effects were on mineral levels and phytic acid levels. They found that germination significantly reduced phytic acid and that different minerals were lowered or raised according to the type of millet germinated. Overall, the conclusion was that there was a good correlation between antinutirtional factors reduced and minerals being more available through germination (Abdelrahaman et al., 2007). Other study used rye and barley, basically they took rye and barley and soaked it, then sprouted it and studied the effects on phytase and phytic acid. The soaking alone had some effect on phytic levels in the case of rye but the sprouting increased phytase and degraded phytic acid dramatically 84% phytic acid degradation for rye and 58% for barley (Centeno et al., 2001). Germination of cereals helps to make their minerals more available to our body. Consuming grains regularly that are only germinated will lead to excess intake of phytic acid.
Fermentation: Fermentation can be defined as the metabolic process by which carbohydrates are oxidized, releasing energy in the absence of external electron acceptor. The recent electron acceptors are organic compounds produced directly by the decomposition of carbohydrates.
According to Cheftel and Cheftel (1976), it can be defined as a desirable process of biochemical changes caused by microorganisms and their enzymes on the products of primary processing. In addition, fermentation provides a natural way to reduce the volume of the material to be transported, to destroy undesirable components, to enhance the nutritive value and appearance of the food, to reduce the energy required for cooking and to make a safer product (FAO, 1995).
The natural fermentation is a widespread practice among the world's technology transformation of cereals, vegetables and nuts. All are from the fermentation pyruvate hub of metabolism. According to Simango (1997), the conversion of monosaccharides (glucose, fructose) and disaccharides (lactose, maltose) pyruvic acid mainly follows the path of glycolysis still being called Embden-Meyerhof-Parnas named their descriptors.
A large number of microorganisms involved in fermentation process. Some of them produce amylases that could act on starch flour used for the preparation of boiled and thus reduce their ability to bind water.
However, the route then followed by pyruvic acid depends on the type of microorganism involved in the process of fermentation and metabolism:the lactic acid bacteria convert pyruvic acid into lactic acid by a hydrogenation reaction; on the other hand, yeasts convert it into ethanol by the reaction of decarboxylation. On the pasta of cereal, there are two types of lactic fermentation. The homolactic fermentation where glucose is converted primarily into lactic acid and the heterolactic fermentation where, in addition to lactic acid, there is the production of other compounds that are:CO2, acetic acid, ethanol and glycerol. The products formed during fermentation are the results of activities of enzymes secreted by the carbohydrate substrates, the homo and heterofermentative bacteria involved in fermentation and microflora present in cereal grains during their production.
Natural fermentation also provides optimum pH conditions for enzymatic degradation of phytic acid which is present in cereals in the form of complexes with polyvalent cations such as iron, zinc, calcium, magnesium and proteins. Such a reduction in phytate may increase the amount of soluble iron, zinc and calcium several folds (Haard et al., 1989). Mohamed et al. (2007) have shown through their result that fermentation of millet grains for 12 and 24 h could reduce the antinutritional factors (phytic acid and tannins). The increase in HCI extractable minerals may be attributed to reduction in phytate and presence of enhancers such as organic acids and ascorbic acid (Indumadhavi and Agte, 1992).
Knowing the microorganisms involved in fermentation, some authors have used direct on the raw material (cereal flours) to know its effect on phytic acid.
Reale et al. (2004) looked at the effect of Lactobacillus plantarum, Lactobacillus brevis, Lactobacillus curvatus and Saccharomyces cerevisiae strains on its ability to hydrolyze IP6 of Italian sourdough technique and they conclude that sourdough technology using lactic acid bacteria is now suggested to reduce phytic acid content. In a further study Reale et al. (2007) found that phytate degradation during cereal dough fermentation was positively correlated with endogenous plant phytase activity and also that when heat was applied to the endogenous cereal phytastes after lactic acid fermentation it resulted in the inactivity of phytase. The optimum phytate degradation was shown to be around pH 5.5. However, they concluded that lactic acid bacteria do not directly participate in phytate degradation but help to bring about a condition in which phytate degradation can be activated by lowering the pH value.
In an in vitro study Leenhardt et al. (2005) found that by acidifying phytic-acid-rich whole wheat dough via sourdough fermentation or by adding lactic acid to the dough, it showed a large phytate breakdown (around 70% compared to 40% control). They concluded that a slight drop in pH (meaning higher acid) is sufficient to reduce phytate content of whole meal flour and magnesium bioavailability was improved.
In a study by Lopez et al. (2000) where five strains of lactic acid bacteria were isolated from sourdough, they were examined to see their ability to degrade phytic acid. The bacteria were placed in both a white flour and whole wheat medium. In the white flour medium, in which phytic sources was the only source of phosphorus, phytate disappeared after only two hours of incubation and an increase of inorganic phosphate was observed. This indicates phytate breakdown, according to the study. In the whole wheat medium where the lactic acid bacterial strain, Leuconostoc mesenteroides strain 38 was placed for 9 h to ferment. Due to the production of lactic acid increase calcium and magnesium solubility were noted compared to the control medium.
Sanz-Penella et al. (2009) observed that breads with fermented Bifidobacterium bacterial strains had lower levels of IP6 and lesser amounts of IP3. They suggested bifidobacterium as a safe means to reduce phytic acid content in rich fibre products intended for human consumption, because of the bacteria phytate-degrading enzyme production.
Qazi et al. (2003) have shown through their studies that fermentation of whole-wheat flour for 45 min can reduce significantly the amount of phytic acid. In their studies, Ghaznavi and Fakhr-e-Sarhad cultivars of wheat were analyzed for phytic acid composition in whole-wheat flour and bread. They were found that the phytic acid content of Ghaznavi for control bread (unleavened) is 752.3 mg 100 g-1 and for bread leavened for 15, 30 and 45 min it was 662.1, 512.1 and 280.3 mg 100 g-1; for Fakhr-e-Sarhad, phytic acid content is 751.7 mg 100 g-1 for control (unleavened) bread, 661.1, 510.8 and 280.4 mg 100 g-1 for fermented breads (leavened) for 15, 30 and 45 min, respectively. Bread preparation from such treated flours dough will have high nutritional impact.
Fermentation is one of the most economic and effective measures for reducing the content of anti-nutritional factors. Studies have shown that both spontaneous fermentations as well as fermentations with starter cultures significantly reduced the content of phytic acid in millet (Sharma and Kapoor, 1996; Elyas et al., 2002; Murali and Kapoor, 2003). One study found starter culture fermentations were to be more effective than spontaneous fermentations (Murali and Kapoor, 2003). Similarly, as a result of lactic acid fermentation, the protein digestibility can be elevated (Antony and Chandra, 1998; Taylor and Taylor, 2002; Ali et al., 2003; Onyango et al., 2004) and the tannin content may be reduced in some cereals, leading to the increased absorption of iron (Khetarpaul and Chauhan, 1989, 1990; Motarjemi and Nout, 1996; Antony and Chandra, 1998; Sanni et al., 1999; Elyas et al., 2002; Onyango et al., 2005).
Soaking: Soaking is widely applied at both household and industrial scale. It is the most important operation in the germination or fermentation process of cereals. During cereals soaking for food making, a natural fermentative acidification takes place and this is regarded as important for food quality. Previously, it was reported that soaking of other cereals such as pearl millet with endogenous or exogenous phytase enzymes at optimum conditions increased the in vitro solubility of iron and zinc by 2-23% (Lestienne et al., 2005a, b). Soaking of sorghum flour (80% extraction) at room temperature for 24 h reduced phytic acid levels by 16-21% (Mahgoub and Elhag, 1998). Soaking of pounded maize for 1 h at room temperature already led to a reduction of phytic acid by 51% (Hotz et al., 2001).
When time is limited, the combination of soaking and cooking has been shown to be much more effective than only soaking for a short duration (Vidal-Valverde et al., 1994; Khokhar and Chauhan, 1986). However, because cooking has also been shown to be less effective at reducing phytic acid (Marfo et al., 1990) soaking is still important. For grains, light cooking is generally a good idea anyway since it further improves digestibility and deactivates natural plant toxins that may still exist after soaking. Although mostly focused on grains and beans, research has shown soaking to be quite effective for the reduction of phytic acid in as little as 12 h as well as the subsequent increase in mineral availability (Mbithi-Mwikya et al., 2000; Duhan et al., 1989; Perlas and Gibson, 2002; Ologhobo and Babatunde, 1984). The seeds being soaked can easily absorb chemicals from the water or container that they’re soaking in and in some cases, such chemicals may be even more of a concern than the phytic acid and protease inhibitors that you’re going out of your way to avoid. As such, it’s important to use a glass container and filtered water. Because phytase enzymes tend to function optimally at a slightly acidic pH at warmer temperatures (Sung et al., 2005; Greiner et al., 2000), it may also be beneficial to use warm water and increase its acidity with something like lemon juice.
TREATMENT EFFECTS ON NUTRITIONAL QUALITY
Cereal grains are considered to be one of the most important sources of dietary proteins, carbohydrates, vitamins, minerals and fibre for people all over the world. Cereals provide the bulk (45%) of food calories of humanity (FAO, 2003). However, the nutritional quality of cereals and the sensorial properties of their products are sometimes inferior or poor in comparison with milk and milk products. The reasons behind this are the lower protein content, the deficiency of certain essential amino acids (lysine), the low starch availability and the presence of antinutrients (phytic acid, tannins and polyphenols) and the coarse nature of the grains. Several methods have been employed with the aim of ameliorate the nutritional qualities of cereals. These include genetic improvement and amino acid supplementation with protein concentrates or other protein-rich sources such as grain legumes or defatted oil seed meals of cereals. Additionally, several pre-treatments which include roasting, fermentation, soaking and germination, have been put into practise to improve the nutritional properties of cereals. The application of these technological processes provokes biochemical and physico-chemical changes of the cereals components. They improve the nutritional quality of cereals products if they are not excessive. To illustrate the treatment effects on cereal nutritional quality, we will focus on a grain, millet.
Effect of roasting: Roasting can improve protein digestibility but has little or no effect during later preparation (Nout, 1993). Roasting is an important unit operation in processing of grain for making sattu due to its significant effect on the odour in the final product sattu which is the most desired quality of sattu (Mridula et al., 2008). In the same study, hardness of pearl millet grain increased with increase in roasting temperature and time. The reason for the increase in hardness at increased temperature and time may be attributed to decrease in moisture content of the grain during the roasting process.
According to Sade (2009), the proteins content of millet decrease during roasting. That is due to the destruction of amino acid by heat. The same thing has noted by Mauron (1982). According to the Fig. 4 from roasting, affect the colour of pearl millet grain. When the roasted pearl millet was milled into flour; the colour change is due to the colour of the endosperm.
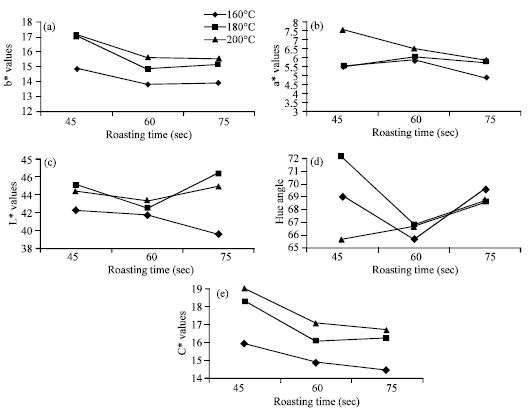 |
| Fig. 4: | (a-e) Effect of roasting on colour (L*, a* and b* values, h0 and C* (chroma) of pearl millet roasted at 160, 180 and 200°C temperature and 45, 60 and 75 sec time |
In the seed form, colour basically represents the colour of the seed coat while in ground form; it is the mixed effect of the colour of all component of the grain. Heat can kill or inactivate potentially harmful organisms including bacteria and viruses. Roasting reduces the amount of aflatoxins produced by fungi such as Aspergillus flavus and Aspergillus parasiticus. This was shown by Conway and Anderson (1978). Griffith and Castell-Perez (1998) reported the millet seed temperatures exceeded 80°C within 10 min which corresponded to aroma development, a pleasant roasted flavor developed when seeds approached 140°C.
Higher seed temperatures produced undesirable flavors and darkened colors due to heat-enhanced chemical reactions. The goal of roasting is to improve sensory qualities and achieve inactivation of destructive enzymes which improves the storage and nutritional quality of the product. Rackis et al. (1986) reported reduced trypsin inhibitor activity when seed temperatures reached 90-100°C. Chen and Whitaker (1986) reported also lipoxygenase activity was lost at temperatures of 75-80°C. Shinde et al. (1991) reported that during roasting total phenols and tannins decrease. Malik et al. (2002) observed the reduction in mineral contents during roasting; he said that might be due to the lost of nutrients while heating at high temperature. It should be noted that, the drying effect of roasting reduces the moisture content of the flour. Reduced moisture allows a larger concentration of solids by weight, resulting in an increased viscosity. According to Griffith and Castell-Perez (1998) roasting increased pearl millet viscosity by 27% over control flours.
Effect of fermentation: Natural fermentation of cereals leads to a decrease in the level of carbohydrates as well as some non-digestible poly and oligosaccharides. Certain amino acids may be synthesised and the availability of B group vitamins may be improved. Fermentation also provides optimum pH conditions for enzymatic degradation of phytic acid which is present in cereals in the form of complexes with polyvalent cations such as iron, zinc, calcium, magnesium and proteins. Reduction in phytate increase the amount of soluble iron, zinc and calcium several folds (Haard et al., 1989). HCI extractable minerals increase, due to reduction in phytate with presence of enhancers such as organic acids and ascorbic acid (Indumadhavi and Agte, 1992). The effect of fermentation on the protein and amino acids levels is a topic of controversy. The fermentation significantly improves the protein quality as well as the level of lysine in millet and other cereals (Hamad and Fields, 1979). In the same way, Nanson and Fields (1984) shown that, during the fermentation of corn meal the concentrations of available lysine, methionine and tryptophan increase. We must remember that the effect of fermentation on the nutritive value of foods is variable but the evidence for improvements is substantial. During millet fermentation several volatile compounds are formed which contribute to improve the shelf life, texture, taste and aroma of the final product. The presence of diacetyl acetic acid and butyric acid make fermented millet and other cereals-based products more appetizing. The acid-producing microorganisms, especially lactic acid, are of particular interest. The increase in pH below 4, would alone may decrease the ability of starches to fix the water during the hydrothermal treatment (Svanberg, 1995).
Cereals are particularly important substrates for fermented foods in all parts of the world and are staples in the Indian, in Asia and in Africa. Fermentation causes changes in food quality indices including texture, flavour, appearance, nutrition and safety. Indeed Mugula et al. (2003) observed a decrease in the concentration of starch and soluble sugars in the production of togwa, a Tanzanian fermented food. The author explains the phenomenon by the hydrolysis of starch and processing of fermentable sugars produced organic acids, the most important are:lactic acid, acetic acid, pyruvic acid, formic acid and the citric acid. In addition Odunfa and Adeyele (1985) showed that fermentation reduced the concentration of raffinose and stachyose, sugars causing flatulence. A significant increase in levels of vitamin B was also observed during the fermentation of sorghum through the action of yeast (Kazanas and Fields, 1981). These observations are consistent with the findings of Murdock and Fields (1984) who noted that three days of fermentation can increase the levels of Vitamin B12, riboflavin and folic acid in the fermented porridge. Furthermore Sripriya et al. (1997) reported an improvement of the bioavailability of copper, zinc, magnesium, calcium, phosphorus and iron during the fermentation of millet. According to Helland et al. (2004) ethanol produced by yeasts, organic acids produced by bacteria and anaerobic conditions induced by fermentation inhibit the development of pathogenic microorganisms. Note that the antimicrobial effect associated with the lactic fermentation is due to the action of a variety of metabolites synthesized during the fermentation process. Among these metabolites are organic acids, the diacetyl, CO2, antibiotics and bacteriocins.
Sripriya et al. (1997) observed an increase in the concentration of total phenols during the fermentation of millet and explains this by the hydrolytic activity of microorganisms that degrade tannins phenolic compounds of low molecular weight. However, further study on the fermentation of millet (El-Hag et al., 2002) reveal a decrease in polyphenol content. This reduction is due to the activity of polyphenoloxidase from the microbial fermentation.
The natural fermented foods prepared from most common types of cereals are well known in many parts of the world. The microbiology of many of these products is quite complex and not known. In most of these products the fermentation is natural and involves mixed cultures of yeasts, bacteria and fungi. Some microorganisms may participate in parallel, while others act in a sequential manner with a changing dominant flora during the course of the fermentation. The common fermenting bacteria are species of Leuconostoc, Lactobacillus, Streptococcus, Pediococcus, Micrococcus and Bacillus. The fungi in general Aspergillus, Paecilomyces, Cladosporium, Fusarium, Penicillium and Trichothecium are the most frequently found in certain products. The common fermenting yeasts are species of Saccharomyces which results in alcoholic fermentation.
Fermentation of food grains is known to be an effective method of improving the starch and protein digestibility and bioavailability of minerals (Boralkar and Reddy, 1985; Mahajan and Chauhan, 1987). Fermentation also brings down the level of anti-nutrients like phytic acid and polyphenols (Dhankher and Chauhan, 1987a). During fermentation the rapid drops in pH with corresponding increase in titratable acidity have been reported by Dhankher and Chauhan (1987b). According to Khetarpaul and Chauhan (1989) natural fermentation brought about a marginal non significant change in the protein content of the pear millet flour and single mixed with sequential pure culture fermentations either decreased or did not change the protein content of the pearl millet. After several readings it should be noted that fermentation of millet either decreased or did not alter the protein content. Increased protein catabolism by fermenting microorganisms may account for loss of protein by escaping ammonia. Khetarpaul and Chauhan (1989) shown that natural fermentation improved the fat content in the millet, pure culture fermentation decreased or did not alter the fat content, the mixed culture fermentation did not change the fat content. Some yeast strains are known to be fat producing (Pandey and Bhaskaram, 1987) and their likely participation in the uncontrolled fermentation may account for the increased amount of fat in the naturally fermented pearl millet.
Fermentation was found to cause a gradual reduction in a pH with time. Elyas et al. (2002) showed that the change in pH from zero to 36 h resulted in a pH drop from 5.9 to 3.6 for pearl millet; these results indicated that fermentation of pearl millet dough caused a reduction in pH and this was more pronounced after 8 h of fermentation. Giese (1994) obtained the same results; he reported that, the increased acidity and low pH enhances the keeping quality of millet foods, by inhibiting microbial growth and also contributing to the flavour of processed millet. Agte et al. (1997) reported that the levels of tannins in pearl millet were unaffected by fermentation. Inyang and Zakari (2008) observed during fermentation of pearl millet the increase of fat content. Other authors reported that, the total fat content of the millet and in other cereals was not affected by fermentation (Antony et al., 1996; Chavan et al., 1989).
Effect of germination: Germination transforms the appearance of seeds but also their nutritional value. Vitamins synthesized during germination increased by 6 to 10 times compared to their levels in the non-germinated seeds. In germinated seeds, proteins, fats and starches are made more assimilated with the action of endogenous enzymes. In the same direction Martinez et al. (1980) said the germination of seeds improves the nutritional value of cereals while raising their content in lysine and in tryptophan and in vitamins such as the Vitamins C, B, A and E. Soaking helps to eliminate flatulence factors (stachyose, raffinose) and reduced phytate content (FAO, 1995). Germination begins, thanks to amylases and proteases synthesized by hydrolysis of starch and protein, providing sugars and amino acids directly assimilated by the body. During germination, enzymes convert the starch amylolytic simple sugars (glucose, maltose) and complex maltodextrins. Among these enzymes are distinguished phosphorylases and amylases. Phytases are responsible in plants from the release of inorganic phosphorus during the germination of seeds (reserve of phosphorus in seeds are in the form of phytate) (Hatzack et al., 2001) and the production of free myo-inositol (non phosphate) is an important growth factor. Coulibaly and Chen (2011) observed phytase activity during germination of Foxtail millet, they noted that, the level of phytase activity reached a maximum at 7 days of germination, this maximum value of phytase activity is 70.023 μg Pi/h/mg of proteins. On the other hand, the hydrolysis of phytate also releases cations could be bound to phytate chelating due to its negative charge. There was also a reduction in phytic acid content with increasing duration of germination (Mamudu et al., 2005). We may also note that germination is causing profound changes in the seed and derivatives, thus giving them a special flavour. On account of hydrolysis of starch, derived porridge from cereals sprouts has a low viscosity. Thus the porridge from cereal sprouts for children have a nutritional certain advantage:high energy density, intake of macronutrients and micro level (Kayode et al., 2005). Studies such as that Kouakou et al. (2008) showed that the seeds of cereals during their germination and fermentation develop a strong enzymatic activity (amylolytic) Fig. 5, the maximum value of amylolytic activity was 0.04 and 0.01 μg/min/mg of protein, respectively for germination and fermentation.
The α-amylase of cereals is soluble in water (Muralikrishna and Nirmala, 2005). In generally, α-amylase is completely inactivated at pH below 4 and temperatures above 70°C. Their optimum pH stand between 4,5 and 5,5. The isoenzymes present in a single grain may or may not have the same pH optimum it the same for optimum temperature. Thus, finger millet, a form α1, respectively for optimum temperature and pH 5.0 and 45°C and is completely inhibited at 75°C. The α2 and α3 have optimum pH and temperature, respectively 5.5 and 5.0, 50 and 40°C and completely inactivated at 55 and 70°C. The most active is the α3.
Carbohydrate contents in the cereals decrease during the germination because of the utilisation of some sugars during the growth metabolic activity. This has been observed by Sade (2009) and Nnam (2000). Similar observation was made by Sripriya et al. (1997) who reported that starch content decrease and total sugars increased during germination of finger millet Mamudu et al. (2005) also reported that sucrose, maltose, glucose and fructose of the grains increased in the course of germination Demuyakor and Ohta (1992) have studied the malt characteristic of sorghum vulgares varieties and reported that dextrin, glucose and maltose increased during germination.
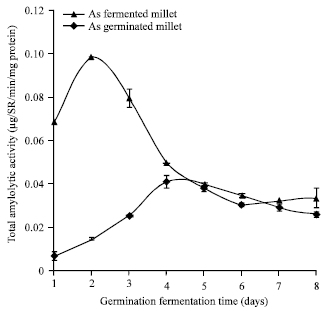 |
| Fig. 5: | Changes in amylase activity during natural fermentation and germination of millet from 1 to 8 days at 25°C |
Germination significantly increased the protein, dry matter and ash content while fat content and energy values of the flour samples showed a decrease (Ocheme and Chinma, 2008). Abdelrahaman et al. (2007) reported that germination of various pearl millet cultivars increased significantly the HCl-extractable parts of both major and trace minerals and also reduced significantly the phytic acid and polyphenol content of the cultivars. Many workers have also observed increase in proteins during germination of various cereals (Akpapunam et al., 1996). This increase could be attributed to a synthesis of enzymatic proteins by germinating seeds (WHO, 1998). Nzeribe and Nwasike (1995) reported increased activities of proteases during germination of acha. Marero et al. (1988) also reported that the increase in protein might be due to the fact some amino acids are produced in excess of the requirement during proteins synthesis and these tend to accumulate in free amino acid pool.
CONCLUSION
Phytic acid by some authors is anti-nutritional factor; others find that it could be used for therapeutic purposes. However, a great controversy exits as to the desirability of eliminating it from food. It is necessary to point out the relevance of an adequate combination of food in order to have a correct intake of all the nutrients that are essential to the organism, especially in those population groups where the impact of higher phytate concentrations can be more serious. This is the case with children, whose mineral requirements are particularly critical. During the weaning period, the diet of an infant is based upon cereal flours as well as upon baby milk formulas. Therefore, the negative properties of phytates, particularly on the bioavalability of minerals, may have an effect on health during this first period. For example, low iron is more likely a problem than high iron for young women, for women who are post-menopausal, high iron may be a problem but then again, low iron may still be a problem. Excess iron is implicated in disease as well and including phytates in our diet is an effective way to reduce our body’s iron levels.
Through this article, our goal is not to scare them with foods containing phytic acid, only to draw your attention using cereals in your diet. It is not necessary to completely eliminate the phytic acid in your food but to keep it to acceptable levels. According to individual needs for example, children under age six, pregnant women or those with serious illnesses, it is best to consume a diet as low in phytic acid as possible.
In short depending on our own circumstances and stage in the life cycle, our needs are going to be different.
REFERENCES
- Coulibaly, A. and J. Chen, 2011. Evolution of energetic compounds, antioxidant capacity, some vitamins and minerals, phytase and amylase activity during the germination of foxtail millet. Am. J. Food Technol., 6: 40-51.
CrossRefDirect Link - Abdelrahaman, S.M., H.B. Elmaki, W.H. Idris, A.B. Hassan, E.E. Babiker and A.H. El-Tinay, 2007. Antinutritional factor content and hydrochloric acid extractability of minerals in pearl millet cultivars as affected by germination. Int. J. Food Sci. Nutr., 58: 6-17.
CrossRef - Akpapunam, M.A., S.O. Igbedioh and I. Aremo, 1996. Effect of malting time on chemical composition and functional properties of soybean and bambara groundnut flours. Int. J. Food Sci. Nutr., 46: 27-33.
Direct Link - Ali, A.M., H.A. El-Tinay and H.A. Abdalla, 2003. Effect of fermentation on the in vitro digestibility of pear millet. Food Chem., 80: 51-54.
CrossRef - Antony, U. and T.S. Chandra, 1998. Antinutrient reduction and enhancement in protein, starch and mineral availability in fermented flour of finger millet (Eleusine coracana). J. Agric. Food Chem., 46: 2578-2582.
CrossRef - Demuyakor, B. and Y. Ohta, 1992. Malt characteristics of sorghum vulgares varietes from Ghana. J. Sci. Food Agric., 59: 457-462.
CrossRefDirect Link - Kayode, A.P.P., D.J. Hounhouigan, M.J.R. Nout and A. Niehof, 2005. Household production of sorghum beer in Benin: Technological and socio-economic aspects. Int. J. Consumer Stud., 31: 258-264.
CrossRef - Inyang, C.U. and U.M. Zakari, 2008. Effect of germination and fermentation of pearl millet on proximate, chemical and sensory properties of instant Fura: A Nigerian cereal food. Pak. J. Nutr., 7: 9-12.
CrossRefDirect Link - Centeno, C., A. Viveros, A. Brenes, R. Canales, A. Lozano and C. De La Cuadra, 2001. Effect of several germination conditions on total P, phytate P, phytase and acid phosphatase activities and inositol phosphate esters in rye and barley. J. Agric. Food Chem., 49: 3208-3215.
CrossRef - Chen, A.O. and J.R. Whitaker, 1986. Purification and characterization of a lipoxygenase from immature English peas. J. Agric. Food Chem., 34: 203-211.
CrossRef - Cook, J.D., M.B. Reddy, J. Burri, M.A. Juillerat and R.F. Hurrell, 1997. The influence of different cereal grains on iron absorption from infant cereal foods. Am. J. Clin. Nutr., 65: 964-969.
CrossRefDirect Link - Cebrian, D., A. Tapia, A. Real and M.A. Morcillo, 2007. Inositol hexaphosphate: A potential chelating agent for uranium. Radiat. Prot. Dosim., 127: 477-479.
CrossRef - Dhankher, N. and B.M. Chauhan, 1987. Effect of temperature and fermentation time on phytic acid and polyphenol content of rabadi: A fermented pearl millet food. J. Food Sci., 52: 828-829.
CrossRef - Dhankher, N. and B.M. Chauhan, 1987. Technical note: Preparation, acceptability and B vitamin content of rabadi: A fermented pearl millet food. Int. J. Food Sci. Technol., 22: 173-176.
CrossRef - Duhan, A., B.M. Chauhan, O. Punia and A.C. Kapoor, 1989. Phytic acid content of chickpea (Cicer arietinum) and black gram (Vigna mungo): Varietal differences and effect of domestic processing and cooking methods. J. Sci., Food Agric., 49: 449-455.
CrossRef - Egli, I., L. Davidsson, M.A. Tuillert, D. Barclay and R.F. Hurrell, 2002. The influence of soaking and germination on the phytase activity and phytic acid content of grains and seeds potentially useful for complementary feeding. J. Food Sci., 67: 3484-3488.
CrossRef - El-Hag, M.E., A.H. El-Tinay and N.E. Yousif, 2002. Effect of fermentation and dehulling on starch, total polyphenols, phytic acid content and in vitro protein digestibility of pearl millet. Food Chem., 77: 193-196.
CrossRef - Elmaki, H.B., E.E. Babiker and A.H. El-Tinay, 1999. Changes in chemical composition, grain malting, starch and tannin contents and protein digestibility during germination of sorghum cultivars. Food Chem., 64: 331-336.
CrossRefDirect Link - Mohamed, M.E., B.H. Amro, A.S. Mashier and E.B. Elfadil, 2007. Effect of processing followed by fermentation on antinutritional factors content of pearl millet (Pennisetum glaucum L.) cultivars. Pak. J. Nutr., 6: 463-467.
CrossRefDirect Link - Elyas, S.H.A. A.H. El-Tinay, N.E. Yousif and E.A.E. Elsheikh, 2002. Effect of natural fermentation on nutritive value and in vitro protein digestibility of pearl millet. Food Chem., 78: 75-79.
CrossRefDirect Link - Sade, F.O., 2009. Proximate, antinutritional factors and functional properties of processed pearl millet (Pennisetum glaucum). J. Food Technol., 7: 92-97.
Direct Link - Greiner, R., K.D. Jany and M.L. Alminger, 2000. Identification and properties of myo-inositol hexakisphosphate phosphohydrolases (phytases) from barley (Hordeum vulgare). J. Cereal Sci., 31: 127-139.
CrossRef - Hamad, A.M. and M.L. Fields, 1979. Evaluation of protein quality and available lysine of germinated and fermented cereal. J. Food Sci., 44: 456-459.
Direct Link - Hotz, C., R.S. Gibson and L. Temple, 2001. A home-based method to reduce phytate content and increase zinc bioavailability in maize-based complementary diets. Int. J. Food Sci. Nutr., 52: 133-142.
CrossRef - Hatzack, F., F. Hubel, W. Zhang, P.E. Hansen and S.K. Rasmussen, 2001. Inositol phosphates from barley low-phytate grain mutants analysed by metal-dye detection HPLC and NMR. Biochem. J., 354: 473-480.
Direct Link - Hurrell, R.F., 2003. Influence of vegetable protein sources on trace element and mineral bioavailability. J. Nutr., 133: 2973S-2977S.
PubMedDirect Link - Hurrell, R.F., L. Davidsson, M. Reddy, P. Kastenmayer and J.D. Cook, 1998. A comparison of iron absorption in adults and infants consuming identical infant formulas. Br. J. Nutr., 79: 31-36.
PubMed - Hurrell, R.F., M.A. Juillert, M.B. Reddy, S.R. Lynch, S. Dassenko and J.D. Cook, 1992. Soy protein, phytate and iron absorption in humans. Am. J. Clin. Nutr., 56: 573-578.
PubMed - Indumadhavi, M. and V. Agte, 1992. Effect of fermentation on ionizable iron in cereal-pulse combinations. Int. J. Food.Sci. Technol., 27: 221-228.
CrossRef - Liang, J., B.Z. Han, M.J.R. Nout and R.J. Hamer, 2008. Effects of soaking, germination and fermentation on phytic acid, total and in vitro soluble zinc in brown rice. Food Chem., 110: 821-828.
CrossRef - Lima-Filho, G.L., U.C. Araujo, G.M.T. Lima, L.C.M. Aleixo and S.R.F. Moreno et al., 2004. A new in vitro enzymatic method to evaluate the protective effect of phytic acid against copper ions. Pak. J. Nutr., 3: 118-121.
CrossRefDirect Link - Agostini, J.D.S., R.B. Nogueira and E.I. Ida, 2010. Lowering of phytic acid content by enhancement of phytase and acid phosphatase activities during sunflower germination. Braz. Arch. Biol. Technol., 53: 975-980.
Direct Link - Kazanas, N. and M.L. Fields, 1981. Nutritional improvement of sorghum by fermentation. J. Food Sci., 46: 819-821.
CrossRefDirect Link - Khetarpaul, N. and B.M. Chauhan, 1989. Effect of fermentation by pure cultures of yeasts and lactobacilli on phytic acid and polyphenol content of pearl millet. J. Food Sci., 54: 780-781.
CrossRef - Khetarpaul, N. and B.M. Chauhan, 1990. Fermentation of pearl millet flour with yeasts and lactobacilli: In vitro digestibility and utilization of fermented flour for weaning mixtures. Plant Foods Hum. Nutr., 40: 167-173.
CrossRef - Konietzny, U., R. Greiner and K.D. Jany, 1994. Purification and characterization of a phytase from spelt. J. Food Biochem., 18: 165-183.
CrossRefDirect Link - Griffith, L.D. and M.E. Castell-Perez, 1998. Effects of roasting and malting on physicochemical properties of select cereals and legumes. Cereal Chem., 75: 780-784.
Direct Link - Hallberg, L., M. Brune and L. Rossander, 1989. Iron absorption in man: Ascorbic acid and dose-dependent inhibition by phytate. Am. J. Clin. Nutr., 49: 140-144.
Direct Link - Lestienne, I., P. Besancon, B. Caporiccio, V. Lullien-Pellerin and S. Treche, 2005. Iron and zinc in vitro availability in pearl millet flours (Pennisetum glaucus) with varying phytate, tannin and fiber contents. J. Agric. Food Chem., 53: 3240-3247.
CrossRef - Lestienne, I., C. Mouquet-Rivier, C. Icard-Verniere, I. Rochette and S. Treche, 2005. The effects of soaking of whole, dehulled and ground millet and soybean seeds on phytate degradation and Phy/Fe and Phy/Zn molar ratios. Int. J. Food Sci. Technol., 40: 391-399.
CrossRefDirect Link - Larsson, M. and A.S. Sandberg, 1992. Phytate reduction in oats during malting. J. Food Sci., 57: 994-997.
CrossRefDirect Link - Mahajan, S. and B.M. Chauhan, 1987. Phytic acid and extractable phosphorus of pearl millet flour as affected by natural lactic acid fermentation. J. Sci. Food Agric., 41: 381-386.
CrossRef - Mahgoub, S.E.O. and S.A. Elhag, 1998. Effect of milling, soaking, malting, heat-treatment and fermentation on phytate level of four Sudanese sorghum cultivars. Food Chem., 61: 77-80.
CrossRef - Makokha, A.O., R.K. Oniang'o, S.M. Njoroge and O.K. Kamar, 2002. Effect of traditional fermentation and malting on phytic acid and mineral availability from sorghum (Sorghum bicolor) and finger millet (Eleusine coracana) grain varieties grown in Kenya. Food Nutr. Bull., 23: 241-245.
PubMedDirect Link - Malik, M., U. Singh and S. Dahiya, 2002. Nutrient composition of pearl millet as influenced by genotypes and cooking methods. J. Food Sci. Technol., 39: 463-468.
Direct Link - Marero, L.M., E.M. Payumo, E.C. Librando, W.N. Lainez, M.D. Gopez and D. Homma, 1988. Technology of weaning food formulations prepared from germinated cereals and legumes. J. Food Sci., 53: 1391-1395.
CrossRefDirect Link - Marfo, E.K., B.K. Simpson, J.S. Idowu and O.L. Oke, 1990. Effect of local food processing on phytate levels in cassava, cocoyam, yam, maize, sorghum, rice, cowpea and soybean. J. Agric. Food Chem., 38: 1580-1585.
CrossRef - Poiana, M.A., E. Alexa and M. Bragea, 2009. Studies concerning the phosphorus bioavailability improvement of some cereals used in nourishment. Roumanian Biotechnol. Lett., 14: 4467-4473.
Direct Link - Mbithi-Mwikya, S., J. van Camp, Y. Yiru and A. Huyghebaert, 2000. Nutrient and antinutrient changes in figer millet (Eleusine coracan) during sprouting. Lebensmittel Wissenschaft Technologie, 33: 9-14.
CrossRef - Motarjemi, Y. and M.J. Nout, 1996. Food fermentation: A safety and nutritional assessment. Joint FAO/WHO workshop on assessment of fermentation as a household technology for improving food safety. Bull. World Health Org., 74: 553-559.
PubMed - Mridula, D., R.K. Goyal and M.R. Manikantan, 2008. Effect of roasting on texture, colour and acceptability of pearl millet (Pennisetum glaucum) for making sattu. Int. J. Agric. Res., 3: 61-68.
CrossRefDirect Link - Murali, A. and R. Kapoor, 2003. Effect of natural and pure culture fermentaion of finger millet on zinc availability as predicted from HCl extractability and molar ratios. J. Food Sci. Technol., 40: 112-114.
Direct Link - Muralikrishna, G. and M. Nirmala, 2005. Cereal α-amylases: An overview. Carbohydr. Polym., 60: 163-173.
CrossRefDirect Link - Murdock, F.A. and M.L. Fields, 1984. B-vitamin content of natural lactic acid fermented cornmeal. J. Food Sci., 49: 373-375.
CrossRef - Nanson, N.J. and M.L. Fields, 1984. Influence of temperature of fermentation on the nutritive value of lactic acid fermented cornmeal. J. Food Sci., 49: 958-959.
CrossRef - Nout, M.J.R., 1993. Processed weaning foods for tropical climates. Int. J. Food Sci. Nutr., 43: 213-221.
CrossRef - Nzelibe, H.C. and C.C. Nwasike, 1995. The brewing potential of Acha (Digitaria exilis) malt compared with pearl millet (Pennisetum typhoides) malts and sorghum (Sorghum bicolor) malts. J. Inst. Brewing, 101: 345-350.
CrossRefDirect Link - Ocheme, O.B. and C.E. Chinma, 2008. Effects of soaking and germination on some physicochemical properties of millet flour for porridge production. J. Food Technol., 6: 185-188.
Direct Link - Odunfa, S.A. and S. Adeyele, 1985. Microbiological changes during the traditional production of Ogi-baba, a West African fermented sorghum gruel. J. Cereal Sci., 3: 173-180.
CrossRefDirect Link - Ogbonna, A.C., S.K.C. Obi and B.N. Okolo, 2004. Optimization of proteolytic activities in malting sorghum. Process Biochem., 39: 713-718.
CrossRef - Ologhobo, A.D. and L.F. Babatunde, 1984. The effect of processing on the trypsin inhibitor, hemagglutinin, tannic acid and phytic acid contents of seeds of ten cowpea varieties. J. Food Process Preser., 8: 31-40.
CrossRef - Onomi, S., Y. Okazaki and T. Katayama, 2004. Effect of dietary level of phytic acid on hepatic and serum lipid status in rats fed a high-sucrose diet. Biosci. Biotechnol. Biochem., 68: 1379-1381.
CrossRefPubMedDirect Link - Onyango, C., H. Noetzold, A. Ziems, T. Hofmann, T. Bley and T. Henle, 2005. Digestibility and antinutrient properties of acidified and extruded maize-finger millet blend in the production of uji. LWT-Food Sci. Technol., 38: 697-707.
CrossRef - Perlas, L.A. and R.S. Gibson, 2002. Use of soaking to enhance the bioavailability of iron and zinc from rice-based complementary foods used in the Philippines. J. Sci. Food Agric., 82: 1115-1121.
CrossRefDirect Link - Potter, S.M., 1995. Overview of proposed mechanisms for the hypocholesterolemic effect of soy. J. Nutr., 125: 606s-611s.
PubMed - Qazi, I.M., S. Wahab, A.A. Shad, A. Zeb and M. Ayuab, 2003. Effect of different fermentation time and baking on phytic acid content of whole-wheat flour bread. Asian J. Plant Sci., 2: 597-601.
CrossRefDirect Link - Zhou, R.J. and J.W. Jr. Erdman 1995. Phytic acid in health and disease. Crit. Rev. Food Sci. Nutr., 35: 495-508.
CrossRefDirect Link - Sandberg, A.S. and H. Andersson, 1988. Effect of dietary phytase on the digestion of phytate in the stomach and small intestines of humans. J. Nutr., 118: 469-473.
PubMed - Sanni, A.I., A.A. Onilude and O.T. Ibidapo, 1999. Physicochemical characteristics of weaning food formulated from different blends of cereal and soybean. Zeitschrift Lebensmitteluntersuchung Forschung A, 208: 221-224.
CrossRef - Sharma, A. and A.C. Kapoor, 1996. Levels of antinutritional factors in pearl millet as affected by processing treatments and various types of fermentation. Plant Food Hum. Nutr., 49: 241-252.
CrossRef - Simango, C., 1997. Potential use of traditional fermented foods for weaning in Zimbabwe. Soc. Sci. Med., 44: 1065-1068.
CrossRefDirect Link - Sripriya, G., U. Antony and T.S. Chandra, 1997. Changes in carbohydrate, free amino acids, organic acids, phytate and HCl extractability of minerals during germination and fermentation of finger millet (Eleusine coracana). Food Chem., 58: 345-350.
CrossRef - Taylor, J. and J.R.N. Taylor, 2002. Alleviation of the adverse effect of cooking on sorghum protein digestibility through fermentation in traditional African porridges. Int. J. Food Sci. Technol., 37: 129-137.
CrossRef - Bohn, T., L. Davidsson, T. Walczyk and R.F. Hurrell, 2004. Phytic acid added to white-wheat bread inhibits fractional apparent magnesium absorption in humans. Am. J. Clin. Nutr., 79: 418-423.
Direct Link - Antony, U., G. Sripriya and T.S. Chandra, 1996. The effect of fermentation on the primary nutrients in foxtail millet (Setaria italica). Food Chem., 56: 381-384.
CrossRef - Vidal-Valverde, C., J. Frias, I. Estrella, M.J. Gorospe, R. Ruiz and J. Bacon, 1994. Effect of processing on some antinutritional factors of lentils. J. Agric. Food Chem., 42: 2291-2295.
CrossRefDirect Link - Vucenik, I. and A.M. Shamsuddin, 2003. Cancer inhibition by inositol hexaphosphate (IP6) and inositol: From laboratory to clinic. J. Nutr., 33: 3778S-3784S.
PubMedDirect Link - Wise, A. and D.J. Gilbert, 1981. Binding of cadmium and lead to the calcium-phytate complex in vitro. Toxicol. Lett., 9: 45-50.
Direct Link