
A.T. Nasseri
Department of Pharmaceutical Biotechnology and Pharmaceutical,Sciences Research Center, Faculty of Pharmacy,Shiraz University of Medical Sciences, P.O. Box 71345-1583, Shiraz, Iran
S. Rasoul-Amini
Department of Pharmaceutical Biotechnology and Pharmaceutical,Sciences Research Center, Faculty of Pharmacy,Shiraz University of Medical Sciences, P.O. Box 71345-1583, Shiraz, Iran
M.H. Morowvat
Department of Pharmaceutical Biotechnology and Pharmaceutical,Sciences Research Center, Faculty of Pharmacy,Shiraz University of Medical Sciences, P.O. Box 71345-1583, Shiraz, Iran
LiveDNA: 98.36188
Y. Ghasemi
Department of Pharmaceutical Biotechnology and Pharmaceutical,Sciences Research Center, Faculty of Pharmacy,Shiraz University of Medical Sciences, P.O. Box 71345-1583, Shiraz, Iran
American Journal of Food Technology
Year: 2011 | Volume: 6 | Issue: 2 | Page No.: 103-116







ABSTRACT
Single-cell proteins are the dried cells of microorganism, which are used as protein supplement in human foods or animal feeds. Microorganisms like algae, fungi, yeast and bacteria, utilize inexpensive feedstock and wastes as sources of carbon and energy for growth to produce biomass, protein concentrate or amino acids. Since protein accounts for the quantitatively important part of the microbial cells, these microorganisms, also called single cell protein as natural protein concentrate. With increase in population and worldwide protein shortage the use of microbial biomass as food and feed is more highlighted. Although single cell protein has high nutritive value due to higher protein, vitamin, essential amino acids and lipid content, there is a doubt to be replaced to the conventional protein sources due to their high nucleic acid content and slower in digestibility. They also may be considered as foreign material by body, which may subsequently results in allergic reactions.
PDF Abstract XML References Citation
Received: April 12, 2010;
Accepted: April 28, 2010;
Published: July 06, 2010
How to cite this article
A.T. Nasseri, S. Rasoul-Amini, M.H. Morowvat and Y. Ghasemi, 2011. Single Cell Protein: Production and Process. American Journal of Food Technology, 6: 103-116.
DOI: 10.3923/ajft.2011.103.116
URL: https://scialert.net/abstract/?doi=ajft.2011.103.116
DOI: 10.3923/ajft.2011.103.116
URL: https://scialert.net/abstract/?doi=ajft.2011.103.116
INTRODUCTION
The increasing world deficiency of protein is becoming a main problem of humankind. Since the early fifties, intense efforts have been made to explore new, alternate and unconventional protein. For this reason, in 1996, new sources mainly yeast, fungi, bacteria and algae named Single Cell Protein (SCP) as coined to describe the protein production from biomass, originating from different microbial sources. Microbial biomass has been considered an alternative to conventional sources of food or feed. Large-scale processes for SCP production show interesting features, including:
| • | The wide variety of methodologies, raw materials and microorganisms that can be used for this purpose |
| • | High efficiency in substrate conversion |
| • | High productivity, derived from the fast growth rate of microorganisms |
| • | Independence of seasonal factors (Roth, 1980; Parajo et al., 1995) |
Yeast was the first microorganism whose importance as animal feed supplement was recognized almost a century ago. During World War I, Germany replaced half of imported protein sources by yeast. Pruteen was the first commercial single cell protein used as animal feed additive. From a nutritional viewpoint, Nucleic Acids (NA) content in SCP is one of the main factors hindering its utilization as food. Excessive intakes of NA lead to uric acid precipitation, causing health disorders, such as gout or kidney stone formation.
| Table 1: | Average different compositions of the main groups of microorganisms (% dry weight) |
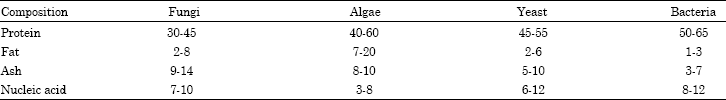 | |
| Miller and Litsky (1976) | |
So, in human their NA contents must be reduced below 2%. Several technologies have been reported to reduce the NA content of microbial cells, including both chemical and enzymatic procedures. Chemical and enzymatic methods have disadvantages (Parajo et al., 1995). Various microorganisms used for the production of SCP are bacteria (Cellulomonas, Alcaligenes, etc.), algae (Spirulina, Chlorella, etc.), molds (Trichoderma, Fusarium, Rhizopus, etc.) and yeast (Candida, Saccharomyces, etc.). Microorganisms can utilize a variety of substrate like agricultural wastes and effluents, industrial wastes, natural gas like methane, etc. that also help in decomposing pollutants (Huang and Kinsella, 1986).
SINGLE CELL PROTEIN DEFINITION
A variety of microorganisms and substrate are used to produce single cell proteins. Yeast is suitable for single cell protein production because of its superior nutritional quality (Table 1). The supplementation cereals with single cell proteins, especially yeast, make them as good as animal proteins (Huang and Kinsella, 1986). The necessary factor considered for use of SCP is the demonstration of the absence of toxic and carcinogenic compounds originated from the substrates, biosynthesized by the microorganisms or formed during processing. High nucleic acid content and low cell wall digestibility are two of the most important factors limiting nutritional and toxicological value of yeast for animal or human consumption (Alvarez and Enriquez, 1988). As constituents of nucleic acid, purine compounds in human diet mostly metabolized to yield uric acid whose high concentration may lead to gout or renal stones. However, nucleic acid is not a toxic component and it causes only physiological effects at higher levels like any other essential dietary ingredients taken in larger amounts. It has been calculated that 100 lbs of yeast will produce 250 tons of proteins in 24 h. Algae grown in ponds can produce 20 tons (dry weight) of protein, per acre, per year. Bacteria are usually high in protein (50 to 80%) and have a rapid growth rate. The principal disadvantages are as follows:
| • | Bacterial cells have small size and low density, which makes harvesting from the fermented medium difficult and costly |
| • | Bacterial cells have high nucleic acid content relative to yeast and fungi. To decrease the nucleic acid level additional processing step has to be introduced and this increases the cost |
| • | The general public thinking is that all bacteria are harmful and produce disease. An extensive education program is required to remove this misconception and to make the public accept bacterial protein |
Yeasts have advantages such as their larger size (easier to harvest), lower nucleic add content, high lysine content and ability to grow at acidic pH. However, the most important advantage is familiarity and acceptability because of the long history of its use in traditional fermentations. Disadvantages include lower growth rates, lower protein content (45 to 65%) and lower methionine content than in bacteria. Filamentous fungi have advantages in ease of harvesting, but have their limitations in lower growth rates, lower protein content and acceptability. Algae have disadvantages of having cellulosic cell walls which are not digested by human beings. Secondly, they also concentrate heavy metals. In the case of algae it has to be stressed that, due to technical and economical reasons, it is not the general intention to isolate and utilize the sole protein, but to propagate the whole algal biomass. So, the term SCP is not quite correct, because the micro-algal material is definitely more than just protein. To date, world-wide various sophisticated technologies are employed for mass production and processing of photoautotrophic microalgae. The annual world production of all microalgae species is estimated to about 10,000 tons year-1 (Becker, 1994, 2007; Richmond, 2004). The algal biomass as sun dried or in compressed form as pastilles is the predominant product in microalgal biotechnology (Radmer, 1996). More than 75% of the annual microalgal biomass production is used for the manufacture of powders, tablets, capsules, or pastilles. This biomass is harvested from natural waters or artificial ponds or photobioreactors (PBR) and subsequently separated from the growth media followed by drying (Renaud et al., 1994). The two major species cultivated for this purpose are the unicellular green alga, Chlorella and more recently, filamentous blue-green alga (Cyanobacterium), Spirulina (Raja et al., 2008). The production of SCP from various microbes, particularly from fungi and bacteria has received considerable attention, in contrast, only a few studies have dealt with the feasibility of using SCP from microalgae (Mahasneh, 1997). Algal proteins are of high quality and comparable to conventional vegetable proteins. However, due to high production costs as well as technical difficulties, cultivation of algae as protein is still in evaluation (Rasoul-Amini et al., 2009). The celluloid cell wall, which represents about 10% of the algal dry matter, poses a serious problem in digesting/utilizing the algal biomass, since it is not digestible for humans and other non-ruminants. Hence, effective treatments are necessary to disrupt the cell wall to make the protein and other constituents accessible for digestive enzymes. Several authors have studied the effect of different post-harvesting treatments on the digestibility of various algal species (Becker, 2007). Different species of algae, fungi, yeasts and bacteria are used as single cell protein and produced at commercial scale (Table 2). These organisms are grown on different carbon sources. Although, microalgae with some other supplements have been used as an essential food for the larval stages of fish and shellfish. Yeast cells have been considered as a substitute because of their small particle size, high protein content as SCP and relatively low production costs (Kim et al., 1998). However, poor digestibility may be an important constraint in the use of this SCP as a food source in seed production of aquacultural organisms, since yeast has a complex and thick cell envelope. The external mannoprotein layer of the yeast cell wall is probably the major barrier to digestion (Rumsey et al., 2007; Kim and Chung, 2001). Several methods have been developed to improve the digestibility of SCP products: mechanical disruption, autolysis and enzymatic treatment (Curran et al., 1990). For maintenance of a stock collection of microorganisms preservation by drying or freeze-drying for long periods of time was reported to be good for yeast or algae (Cordero and Voltolina, 1997; Kim and Chung, 2001). It has been agreed that the criteria used to evaluate SCP production are growth yield, total protein (39-73%) and nucleic acid contents (1-11%). Marine yeasts are better candidates for marifeed production according to their easy cultivation in the fermenter, high cell density and high content of essential amino acids. The substrates which have been used for SCP production by yeasts so far include sorghum hydrolysate, sulfate waste liquor, pawn-shell wastes, dairy wastes, methanol, molasses, starch and plant origin liquid waste. Several fungi like Fusarium oxyporum var., lini and Chetomium cellulolyticum, algae like Chlorella and Spirulina, yeast like Candida lipolytica and Saccharomyces sereviciae and phototrophic bacteria like Rodospirillum sp., had been explored for SCP.
| Table 2: | Microorganism and substrates used for single cell protein production |
 | |
| Bhalla et al. (2007) | |
It is also said that Scytalidium acidophilum compares favorably with the values for the well-known single-cell protein producer Candida utilis. Kluyveromyces marxianus produces enzyme that can be used to modify whey for the production of single cell protein (Chandrani-Wijeyaratne and Tayathilake, 2000). Spirulina maxima is grown on a commercial scale in Lake Texcoco in Mexico. The quality of SCP is an important factor for commercial production. An accurate method to evaluate the quality of proteins is the determination of the Protein Efficiency Ratio (PER), expressed in terms of weight gain per unit of protein consumed by the test animal in short-term feeding trials. Estimation of the Biological Value (BV) is a measure of nitrogen retained for growth or maintenance. Another parameter, which reflects the quality of a protein, is the Digestibility Coefficient (DC). Finally, the Net Protein Utilization (NPU) -equivalent to the calculation BVxDC-is a measure of the digestibility of the protein and the biological value of the amino acids absorbed from the food. The SCP generally values in terms of Kjeldhal nitrogenx6.25 (standard factor relating amino nitrogen to protein content). However, about 10-15% of the total nitrogen in fungi and yeasts is in the form of nucleic acids (Riviere, 1977).
SCP PRODUCTION
The worldwide, large-scale development of SCP processes has contributed greatly to the advancement of present day biotechnology. Research and development of SCP processes has involved work in the fields of microbiology, biochemistry, genetics, chemical and process engineering, food technology, agriculture, animal nutrition, ecology, toxicology, medicine and veterinary science and economics. In developing SCP processes new technical solutions for other related technologies in waste water treatment, production of alcohol, enzyme technology and nutritional science also improves. The future of SCP will be heavily dependent on reducing production costs and improving quality by fermentation, downstream processing and improvement in the producer organisms as a result of conventional applied genetics together with recombinant DNA technologies (Omar and Sabry, 1991). Single cell proteins have application in animal nutrition as: fattening calves, poultry, pigs and fish breading in the foodstuffs area as: aroma carriers, vitamin carrier, emulsifying aids and to improve the nutritive value of baked products, in soups, in ready-to-serve meals, in diet recipes and in the technical field as: paper processing, leather processing and as foam stabilizers. The production of single cell protein takes place in a fermentation process. This is done by selected strains of microorganisms which are multiplied on suitable raw materials in technical cultivation process directed to the growth of the culture and the cell mass followed by separation processes. Process development begins with microbial screening, in which suitable production strains are obtained from samples of soil, water, air or from swabs of inorganic or biological materials and are subsequently optimized by selection, mutation, or other genetic methods. Then the technical conditions of cultivation for the optimized strains are done and all metabolic pathways and cell structures will be determined. Besides, process engineering and apparatus technology adapt the technical performance of the process in order to make the production ready for use on the large technical scale. Here is where the economic factors (energy, cost) come into play. Safety demands and environmental protection is also considered in the production of SCP in relation both to the process and to the product. Finally, safety and the protection of innovation throw up legal and controlled aspects, namely operating licenses, product authorizations for particular applications and the legal protection of new process and strains of microorganisms (Steinkraus, 1986).
The classical raw materials are substances containing mono and disaccharides, since almost all microorgansims can digest glucose, other hexose and pentose sugars and disaccharides. These materials also are utilized in other branches of industry with a high price level, which puts the economic aspect of the production of microbial biomass in doubt (Oura, 1983). The choice of substrates that are normally abundant has determined the design and strategy of SCP processes. The most widespread and commonly used substrates for SCP production have been those where the carbon and energy source is derived from. Many companies producing SCP including BP (UK), Kanegafuichi (Japan) and Liquichimica (Italy) appeared on the scene. In the United States less than 15% of the plants making SCP were said to rely on hydrocarbons as the source of carbons and energy for the micro-organisms. Other potential substrates for SCP include bagasse, citrus wastes, sulphite waste liquor, molasses, animal manure, whey, starch, sewage, etc.
SCP derived from high energy sources: Materials with high commercial value as energy sources or derivatives like gas oil, methane, methanol and n-alkanes are of interest in SCP production. The microbes involved are mostly bacteria and yeast and several processes are now in operation. The wisdom of using such high-energy potential compounds for food production has been questioned by many scientists. British Petroleum uses two yeasts, Candida lipolytica and C. tropicalis and C12-C20 alkanes as substrate which is of the wax fraction of gas oils for treating. Some crude oils contain up to 15% in wax. The product produced was called TOPRINA. For 12 years TOPRINA was tested for toxicity and carcinogenecity and was marketed as a replacement for fish meal in high protein feeds and as a replacement for skimmed milk powder in milk replacers. After a while the main opposition came from Japan. In 1972 a specialised committee decided that SCP was only for animal feeding but later, Japan was the first country to ban petrochemical protein. In 1977 Italy stopped the SCP production from alkanes due to the increase in oil prices. The price of soya was more competitive. Now there is no factory which produces any petrochemical protein. Methane as a SCP source has been extensively researched. The species which has been extensively studied is Methylomonas methanica and nitrates or ammonium salts can serve as N-source. The technology used for this purpose faced too many technical difficulties to optimize the production. In contrast, methanol offers great economic SCP interest. A large-scale fermentation plant for producing the methanol-utilizing bacterium Methylophilus methtlotrophus was constructed by ICI (Imperial Chemical Industries Company), UK. The ICI SCP protein was used exclusively for animal feeding. Methanol as a carbon source for SCP has many inherent advantages over n-paraffins, methane gas and even carbohydrates composition is independent of seasonal fluctuations. There are no possible sources of toxicity in methanol and it dissolves easily in the aqueous phase in all concentrations. The ICI Pruteen (72% protein) plant (by Pseudomonas methylotrophus) was the only process of its kind in the Western world but could not operate economically at present methanol prices and cannot compete with soya and fish meal so has ceased production. Methanol represents approximately 50% of the costs of the product. In the USA the cost of SCP derived from methanol is two-to five-folds the cost of fishmeal. In the Middle East the low cost of methanol and higher costs of fishmeal coupled with a need to produce more animal products could make SCP an attractive proposition. Ethanol is a particular suitable source if the SCP is intended for human consumption. The process comes from the Amoco Company in the US utilizing food grade yeast: Torula. The product is sold by the name "Torutein" even in Canada and Sweden. The yeast is about 52% protein. Torutein is being marketed as a flavor enhancer of high nutritional value and a replacement for meat, milk and egg protein. However, it is not very successful in the United States since soya which is plentiful and cheap can serve as an alternative or substitute to meat and egg diets. In the foreseeable future the comparative status of ethanol SCP will depend on local factors: over-capacity in ethylene crackers, agricultural carbohydrates surpluses and political decisions about regional economic independence and foreign trade balances (Trinci, 1992). The use of n-alkanes as substrate for SCP has been extensively studied many countries and represents a very complex biotechnological process. However, most of these processes have now ceased operation because of suspected health hazards resulting from the presence of carcinogens in the SCP. The massive technology developed in this field in Japan and other Eastern countries has been turned over to the study of alcohol- based SCP and SCP form organic wastes. Many genera of yeasts and molds utilize aliphatic hydrocarbons (alkanes and alkenes) for growth and for SCP production. These microorganisms secrete emulsifying substances during fermentation which increase the solubility of alkanes and alkenes. These substrates are transported by passive diffusion into the cell. The higher lipid content in these microorganisms facilitates the hydrocarbon transport. For large-scale production of SCP, gas oil and C10-C17 alkanes were used as substrate. Gas oil fermentation has been carried out in airlift fermenter using Candida tropicalis and C. lipolytica (Riviere, 1977).
SCP from wastes: The amount of agricultural and some industrial wastes used for SCP production can be locally very high and may contribute to a significant level of pollution in watercourses. Thus, the utilization of such materials in SCP processes serves two functions as reduction in pollution and creation of edible protein. Cellulose from agriculture and forestry sources constitutes the most abundant renewable resource in the planet as potential substrates for SCP production. Cellulose has emerged as an attractive substrate for SCP production but in nature it is usually found with lignin, hemicellulose, starch, etc., in a complex form. Therefore, if cellulose is to be used as substrate it must be pretreated chemically (acid hydrolysis) or enzymatically (cellulases) to remove cellulose as fermentable sugars (Callihan and Clemmer, 1979). For the utilization of lignocellulose, a pre-treatment is usually necessary. Many pre-treatment methods have been reported which vary from alkali or acid treatment, steam explosion or even x-ray radiation. To the present time the only economical utilization of lignocellulosic wastes is in mushroom production. Besides our well known cultivated mushroom Agaricus bisporus there are many important ones which contain lignocellulolytic enzymes and are cultivated for food mainly in Asia and Africa. Some are of great economic significance and are cultivated on an industrial scale. Examples of important ones include Volvariella sp., Lentinus edodes and Pleurotus sp. In the manufacture of industrial cellulose for paper and tissue production the cost of these steps has prevented the generalized production of SCP from cellulose (Callihan and Clemmer, 1979). Wood can be also cooked in a medium containing calcium sulfite with excess free sulfur dioxide. Lignin is thus converted to lignosulfonates and hemicellulose is hydrolysed to monosaccharides and may be further broken down to furfurols. The amount of free sugars in the spent liquor is variable with the type of procedure chosen, as various cellulose fibers may be obtained with different degrees of degradation. Spent sulfite liquor has been used as a substrate for fermentations since 1909 in Sweden and later in many other parts of the world. The first organism to be used was Saccharomyces cerevisiae, although this organism is unable to metabolise pentoses which are found in considerable amounts in this waste product. Later, other organisms better suited for the assimilation of all the sugar monomers were chosen, namely Candida tropicalis and Candida utilis. Yeast produced from sulfite liquor has been used for feeding at war periods, but lost favour in peace time. However, experiences of baker�s yeast produced from sulfite liquor exist in Finland by Peliko process. The protein content of the fungus Paecilomyces variotii exceeds 55% (w/w) and has been officially approved as a food in Finland. In 1983, the projected biomass production of the process was estimated to be around 7000 tons per year (Oura, 1983). Nowadays, extracellular cellulases are commercially used in cellulose separating process. Cellulase is a complex of three enzymes (endocellulase, cellobiohydrolase, cellobiase). The cost of production of these extracellulases is about $ 0.011 L-1 of crude enzyme and $ 0.11 L-1 of purified enzyme. The cost of extraction of fermentable sugar using cellulase is about $ 0.50 kg-1 sugar. A number of efficient cellulase producers have been reported but Trichoderma viride continued to be well known high cellulase-producing organism. Chaetomium cellulolyticum is another cellulolytic fungus which grows faster and forms 80% more biomass-protein than Trichoderma. This means that C. cellulolyticum is suitable for SCP production while T. viride is a hyper producer of extracellular cellulases. The amino acid composition of C. cellulolyticum is generally better than that of T. viride and similar to alfalfa and soya meal protein (Bhalla et al., 2007). A cheaper, more amenable SCP substrate of carbohydrate origin is starch. This very abundant carbohydrate may be obtained from rice, maize and cereals. In tropical countries, cassava has been proposed as a good source of starch for SCP processes. The Symba process developed in Sweden utilized starchy wastes combining two yeasts in sequential mixed culture: the amylase producing Endomycopsis fibuligira and the fast growing Candida utilis. The process consists of three phases: The incoming starch waste is fed through heat exchangers and sterilized. The medium is then fed to a first bioreactor where the starch hydrolyzing yeast grows and hydrolyses starch. The hydrolyzed solution is then fed to a second reactor where culture conditions favor the proliferation of C. utilis. Whey traditionally originates from the curding process in cheese production, but can now be obtained after ultra filtration procedures for the production of spreading cheeses, where the protein fraction corresponding to lactalbumins and lactoglobulins is incorporated to the casein fraction and all the proteins are in native form with the principal component as lactose (4-6%) (w/v). Although other nutrients can be find in significant amounts (Moebus and Teuber, 1983). Whey has been presented as an extremely suitable substrate for the production of SCP. In 1956 The French dairy company Fromageries Bel pioneered a project to produce yeast from whey, using lactose assimilating Kluyveromyces marxianus (formerly K. fragilis) (Oura, 1983; Moulin et al., 1983). Molasses is a by-product of the sugar manufacturing process. Biomass production from molasses requires supplementation with a suitable nitrogen source, as well as phosphorus (Callihan and Clemmer, 1979). The Ranks Hovis McDougall mycoprotein (except edible mushrooms) has been cleared for human consumption. It uses a Fusarium graminearum growing in molasse, or glucose. The medium contains NH3 for nitrogen source and pH control. The product is heat treated for RNA reduction. The mycelium is separated by vacuum filtration and can be technologically treated to match food texture. In the UK it is marketed as pies and is considered a success since having less fat than meat. Coffee-pressing wastes contain soluble carbohydrates and have a high chemical oxygen demand (COD) and soluble solid contents. In Guatemala, Trichoderma sp. is used to produce SCP on this substrate. Cellulomonas grows on bagasse and Thermoactinomyces on fermented livestock wastes (Humphrey, 1975).
Fermentation process: The fermentation process requires a pure culture of the chosen organism that is in the correct physiological state, sterilization of the growth medium which is used for the organism, a production fermenter which is the equipment used for drawing the culture medium in the steady state, cell separation, collection of cell free supernatant, product purification and effluent treatment. A fermenter is the instrument, which is set up to carry out the process of fermentation mainly the mass culture of plant or animal cells. Fermenters can vary in size from laboratory experimental models of one or two litres capacity, to industrial models of several hundred litres capacity. A bioreactor is different from a fermenter as it used for the mass culture of microorganisms. The chemical compounds synthesised by these cultured cells such as therapeutic agents can be extracted easily from the cell biomass. The design engineering and operational parameters of both fermenters and bioreactors are identical. Fermenters and bioreactors are also equipped with an aerator, which supplies oxygen to aerobic processes also a stirrer is used to keep the concentration of the medium the same. A thermostat is used to regulate temperature and a pH detector and some other control devices, which keep all the different parameters needed for growth constant (Ferrianti and Fiechter, 1983; Sinclair and Cantero, 1990). For the producing and harvesting of microbial proteins cost is a major problem. Such a production even in high rate causes dilute solutions usually less than 5% solids. There are many methods available for concentrating the solutions like filtration, precipitation, centrifugation and the use of semi-permeable membranes. The equipment used for these methods of de-watering is expensive and so would not be suitable for small scale productions and operations. The removal of the amount of water that is necessary to make the material stable for mass storage is not economically viable. Single cell proteins need to be dried to 10% moisture or they can be condensed and denatured to prevent spoilage (Sinclair and Cantero, 1990). The physiological features of microbial organisms recommend the control of the carbon source concentrations, as a limiting substrate, as well as an adequate supply of oxygen for the maintenance of balanced growth under an oxidative metabolic pattern. However, since microbial growth is a time dependent process, it exerts continuous modifications on all process parameters which influence physiology, but most dramatically, over substrate concentration. Therefore, an adequate technology which maintains appropriate growth conditions for a prolonged period of time must be implemented specifically for the purpose of obtaining high yield and productivity values. Batch fermentations are clearly inadequate for the purpose of biomass production, since the conditions in the reaction medium change with time (Oura, 1983). Fed-batch fermentations are better suited for the purpose of biomass production, since they involve the control of the carbon source supply through feeding rates. However, as the biomass concentration increases, the oxygen demand of the culture reaches a level which cannot be met in engineering or economic terms. Fed-batch culture is still in use for bakers yeast production using well established and proven models (Steinkraus, 1986). However, they have not been favored for the production of SCP at a large industrial scale. Prolonging a microbial culture by continuous addition of fresh medium with the simultaneous harvesting of product has been implemented successfully in industrial fermentations destined to biomass production. The most commonly used principle has been the chemostat: a perfectly mixed suspension of biomass into which medium is fed at a constant rate and the culture is harvested at the same rate so that the culture volume remains constant. The technical implications of chemostat culture are various and extremely relevant (Cooney, 1986). Production periods as long as six weeks have been implemented in many fungal and yeast (Forage and Righelato, 1979). A common problem of industrial fermentations is the profuse appearance of foam on the head space of the reactor, causing reactor pressurization, spillages and contamination hazard. Among the various designs which have been put to effect, the deep-jet fermenter and the air-lift fermenter have been the most successfully applied (Stanbury et al., 2000). Air-lift has enjoyed the greatest success as the configuration of choice for continuous SCP production. This is presently used in the production of myco-protein which is the basis for Quorn™ products. The control of key process variables is a critical element of SCP production, from oxygen transfer, substrate and product concentration, to the appearance of minimal amounts of toxic compounds through undesired metabolic processes, which may compromise the quality of the final product. The biomass from yeast fermentation processes is harvested normally by continuous centrifugation. Filamentous fungi are harvested by filtration (Solomons, 1985). The biomass is then treated for RNA reduction and dried in steam drums of spray driers. Drying is expensive, but results in stabilized product with shelf lives of years. Solid State Fermentation (SSF) is growth of microorganisms on predominantly insoluble substrate where there is no free liquid. Generally, under combined conditions of low water activity and presence of intractable solid substrate, fungi show luxuriant growth. Hence, proper growth of fungi in SSF gives much higher concentration of the biomass and higher yield when compared to submerged fermentation. The advantage in SSF process is the unique possibility of efficient utilization of waste as the substrate to produce commercially viable products. The process does not need elaborate prearrangements for media preparation. The process of SSF initially concentrated on enzyme production. But presently, there is worldwide interest for (SCP) production due to the dwindling conventional food resources (Zadrazil and Puniya, 1995).
ECONOMIC ASPECTS
For SCP production large-scale fermenters are required. So with high biomass production, high oxygen transfer rates and high respiration rates which in turn increase metabolic heat production and the need of an efficient cooling system ensued. In such a continuous operation for SCP production the economics of this production must be strongly taken into account. The Economics factors that should be taken into account during this fermentation period are: Investment, Energy, Operating costs, Waste, Safety and the Global market.
Substrate costs: The substrate costs are the largest single cost factor. Simplifying the manufacture and purification of raw material can save costs. Moreover the manufacture of raw materials is more economical in larger plants. Factors involved in the raw materials costs are site, raw material production, process capacity of the plant and substrate yield.
Utilities: The energy for compressing air, cooling, sterilizing and drying forms the next most important cost factor. Sites with cheaply available thermal, electrical, fossil or process derived energy are to be preferred.
Capital load: The capital dependent costs are determined, by the cost of the apparatus for the process, the capacity of a plant and the capacity conditions. The main variable here is the size of the plant. Small plants can be profitable only if they include simplifications of processes and material to a considerable degree. The greater expenditure on apparatus in processes with cheap, simple and unpurified raw materials usually does not pay in comparison with more expensive pure substrates with simpler technology. High productivities in fermentation are compensated by the greater expenditure on energy to achieve these productivities, so that optimum can be determined.
Product-specific variables: The process costs arising are covered only by the product produced. The absolute value of the product is governed by the amount of product referred to the costs involved and by the quality of the product. The upgrading of the product may consist of purification and separation of the microbial biomass.
ACCEPTABILITY AND TOXICOLOGY OF SCP
The name of the raw materials used in SCP processes represents the main safety hazard. Toxicology testing of the final product must include short-term acute toxicity testing with several different laboratory animal species, followed by extensive and detailed long term studies. It represents a major scientific and financial investment. The acceptability of SCP when presented as a human food does not depend only on its safety and nutritional value. In addition to the general reluctance of people to consume material derived from microbes, the eating of food has many subtle psychological, sociological and religious implications.
SCP PROCESSING FOR FOOD
The effective use of microbial protein for human food requires:
| • | Liberation of cell proteins by destruction of indigestible cell walls |
| • | Reduction of nucleic acid content |
Methods of cell wall destruction: The use of microorganism for refined SCP requires not only an adequate amount of specific organism but also an efficient means of disrupting the cell wall (Baldwin and Robinson, 1994). Mechanical integration of cell wall can be carried out either by crushing, crumbling, grinding, pressure homogenization or ultra sonification. Various enzymes or combination of enzymes can be used to digest and disrupt cell wall, either partially or completely. Enzymatic hydrolysis of cell wall is attractive in terms of its delicacy and specificity for only the cell wall structure. It may be used as an alternative to the mechanical disruption, especially for materials that can be inactivated during the mechanical process and it can be performed by endogenous or exogenous enzyme from other microorganisms. However, extensive enzymatic lysis of cells is a very slow process compared to mechanical disruptions. It is possible to use two or more methods for cell disruptions. Combined mechanical and enzymatic degradation of yeast cell wall was tested by Asenjo and Dunnill (1981). In case of yeast cells they first can be mechanically broken and then incubated with a lytic enzyme (Damodaran and Kinsella, 1983). This resulted in the release of a substantial amount of protein mostly from organelles and cell walls. Baldwin and Robinson (1994) reported enhanced disruption of Candida utilis by enzymatic pretreatment and high-pressure homogenization. Other methods employed for yeast cell breakage include: autolysis followed by enzymatic or alkali treatments (Benaiges et al., 1989), NaCl induced autolysis at different temperature, chemical disintegration using detergent such as sodium dodecyl sulfate or Triton X-100, acid or solvent (Lee, 1996). The digestibility of yeast and microalgae can be greatly increased by drying at high temperature under certain conditions. However, the heat treatment needed to increase the digestibility of the cells also affects the protein quality and other valuable cell components (Hedenskog and Morgen, 1973). Here are the methods for cell disruption:
Non-mechanical methods
| • | Chemical treatment: acid, base, solvent, detergent |
| • | Enzyme analysis: lytic enzymes, phage infection, autolysis |
| • | Physical treatment: freeze-thaw, osmotic shock, heating and drying |
Mechanical methods
| • | High pressure homogenization |
| • | Wet milling |
| • | Sonification |
| • | Pressure extrusion: french press, freeze pressing |
| • | Decompression (pressure chamber) |
| • | Treatment with grinding particles (Engler, 1985; Middelberg, 1995) |
Removal of nucleic acids: Several methods have been proposed to reduce nucleic acid levels in SCP. These methods involve chemical and enzymatic treatments. Each has disadvantages both in terms of cost and potential nutritional concern. In 1977, the extraction of nucleic acid by acidified alcohol, salt, acid and alkalis has been proposed. Alkaline extraction of microbial biomass at elevated temperature was also used in 1970. The process resulted in high protein yield with low nucleic acid. However, alkaline hydrolysis of nucleic acid at high temperature causes the formation of potentially toxic compounds such as lysinoalanine. It is an unusual amino acid involved in cross-linking of alkaline protein. Lysinoalanine has been shown to reduce digestion and induce kidneys changes in rats (Damodaran and Kinsella, 1983). It also implicated in skin allergy in some persons consuming treated protein (Scrimshaw and Dillen, 1977). Chemical modification of yeast nucleoproteins with anhydrides has been used to reduce the nucleic acid levels. Yeast contains considerable amounts of endogenous ribonulease activity that is used to hydrolyzed yeast RNA and that cause reduction of nucleic acid level in yeast protein. At the optimum conditions of ribonuclease activity, significant activation of endogenous protease also occurs. This result in proteolytic degradation of protein and thereby, decreases the yield of protein. Alternatively, nuclease has been added exogenously to reduce the nucleic acid content of SCP. Pancreatic ribonuclease (RNase A) and a fungal ribonuclease of Aspergillus candidus strain M16 has been used as the source of exogenous nuclease for the reduction of nucleic acid in the cells of yeast species allowing a substantial reduction of NA (Maul et al., 1970; Kunhi and Rao, 1995). Bacterial or pancreatic nucleases have been also studied for NA removal from yeast cells. Hydrolysis of NA has also been performed by using immobilized enzymes (Parajo et al., 1995).
CONCLUSION
Single celled protein (SCP) production, referring to the fact that most of the micro-organisms used as producers grow as single or filamentous individuals rather than as complex multi-cellular organism such as plants or animals. Use of microbes in the production of proteins gives many advantages over the conventional method methods. Microbes have shorter generation time, allow easy transformation, utilize many substrates, have no requirements in arable land or any particular season to grow and have the possibility of continuous production in any part of the world. The cell yield varies according to the substrate and type of microorganism.
REFERENCES
- Alvarez, R. and A. Enriquez, 1988. Nucleic acid reduction in yeast. Applied Microbiol. Biotechnol., 29: 208-210.
CrossRefDirect Link - Baldwin, C.V. and C.W. Robinson, 1994. Enhanced disruption of Candida utilis using enzimatic pretreatment and high pressure homogenization. Biotechnol. Bioeng., 43: 46-56.
PubMedDirect Link - Becker, E.W., 2007. Micro-algae as a source of protein. Biotechnol. Adv., 25: 207-210.
CrossRefDirect Link - Benaiges, M.D., J. Lopez-Santin and C. Sola, 1989. Partial purification of 5-phosphodiesterase activity from barley rootlets. Enzyme Microbiol. Technol., 11: 444-451.
CrossRef - Cordero, B. and D. Voltolina, 1997. Viability of mass algal cultures preserved by freezing and freeze-drying. Aquac. Eng., 16: 205-211.
CrossRef - Curran, D.A., B.J. Tepper and T.J. Montville, 1990. Use of bicarbonates for microbial control and improved water-binding capacity in cod fillets. J. Food Sci., 55: 1564-1566.
Direct Link - Damodaran, S. and J.E. Kinsella, 1983. The use of chaotropic salts for separation of ribonucleic acids and proteins from yeast nucleo-proteins. Biotechnol. Bioeng., 25: 761-770.
PubMedDirect Link - Ferrianti, M.P. and A. Fiechter, 1983. Production and Feeding of Single Cell Protein. Applied Science Publications, London, ISBN: 085334231, pp: 201.
Direct Link - Hedenskog, G. and H. Morgen, 1973. Some methods for processing of single cell protein. Biotechnol. Bioeng., 15: 129-142.
PubMedDirect Link - Huang, Y.T. and J.E. Kinsella, 1986. Functional properties of phosphorylated yeast protein: Solubility, water-holding capacity and viscosity. J. Agric. Food Chem., 344: 670-674.
CrossRefDirect Link - Kim, J.K. and H.Y. Chung, 2001. Preservation of manipulated yeast diet. Aquac. Int., 9: 171-181.
CrossRefDirect Link - Kim, J.K., K.T. Tak and J.H. Moon, 1998. A continuous fermentation of Kluyveromyces fragilis for the production of a highly nutritious protein diet. Aquac. Eng., 18: 41-49.
CrossRef - Kunhi, A.A.M. and M.R.R. Rao, 1995. The utility of a fungal ribonuclease for reducing the nucleic acid content of permeabilized yeast cells. Food Biotechnol., 9: 13-28.
Direct Link - Maul, S.B., A.J. Sinskey and S.R. Tannenbaum, 1970. New process for reducing the nucleic acid content of yeast. Nature, 228: 181-181.
CrossRefDirect Link - Middelberg, A.P.J., 1995. Process-scale disruption of microorganisms. Biotechnol. Adv., 13: 491-551.
CrossRef - Moulin, G., B. Malige and P. Galzy, 1983. Balanced flora of an industrial fermenter: Production of yeast from whey. J. Dairy Sci., 66: 21-28.
Direct Link - Omar, S. and S. Sabry, 1991. Microbial biomass and protein production from whey. J. Islamic World Acad. Sci., 4: 170-172.
Direct Link - Parajo, J.C., V. Santos, H. Dominguez and M. Vazquez, 1995. NH4OH-based pretreatment for improving the nutritional quality of single cell protein. Applied Biochem. Biotechnol., 55: 133-149.
CrossRefDirect Link - Radmer, R.J., 1996. Algal diversity and commercial algal products. Bioscience, 46: 263-270.
Direct Link - Raja, R., S. Hemaiswarya, N. A. Kumar, S. Sridhar and R. Rengasamy, 2008. A perspective on the biotechnological potential of microalgae. Cr. Rev. Microbiol., 34: 77-88.
Direct Link - Rasoul-Amini, S., Y. Ghasemi, M.H. Morowvat and A. Mohagheghzadeh, 2009. PCR amplification of 18S rRNA, single cell protein production and fatty acid evaluation of some naturally isolated microalgae. Food Chem., 116: 129-136.
CrossRef - Renaud, S.M., D.L. Parry and L.V. Thinh, 1994. Microalgae for use in tropical aquaculture I: Gross chemical and fatty acid composition of twelve species of microalgae from the Northern Territory, Australia. J. Applied Phycol., 6: 337-345.
CrossRefDirect Link - Rumsey, G.L., S.G. Hughes and J.L. Kinsella, 1990. Use of dietary yeast Saccharomyces cerevisiae nitrogen by lake trout. J. World Aquac. Soc., 21: 205-209.
CrossRefDirect Link - Trinci, A.P.J., 1992. Myco-protein: A twenty-year overnight success story. Mycol. Res., 96: 1-13.
CrossRef - Zadrazil, F. and A.K. Puniya, 1995. Studies on the effect of particle size on solid-state fermentation of sugarcane bagasse into animal feed using white-rot fungi. Bioresour. Technol., 54: 85-87.
CrossRefDirect Link
Md. Aabid Hussain Reply
Great !! Good job....very simple language and easy to understand...
Thank You.
Regards:
Aabid
Achmad Farits Fauzi Reply
Subhanalloh ...
Thanks sir ...
I hope this journal can use for next time ...
Dr.sushil mathur Reply
I would like a copy of this article.kindly mail thanks in anticipation,& hope a positive reply.Mathur
uduakobong raphael akpan Reply
It is a good work.pls can send me biotechnology aspect of scp production.thanks
shahab uddin Reply
comprehensive and easy to get.... shahab
Jane S. Fom Reply
I love the work and would like to have a copy.
Asma BEN TEKAYA Reply
awesome
Mayang Reply
The concept of producing protein from microorganisms like algae, fungi, yeast, and bacteria is an innovative approach to address the worldwide protein shortage. The use of inexpensive feedstock and wastes as sources of carbon and energy for growth to produce biomass or protein concentrate is an added advantage. Single cell proteins have high nutritive value with higher protein, vitamin, essential amino acids, and lipid content. However, the concern over their high nucleic acid content, slower digestibility, and potential for allergic reactions may raise questions about their widespread use as a replacement for conventional protein sources. Further research and development in this area could address these concerns and contribute to sustainable protein production.