
L. Abubakar
Department of Crop Science, Faculty of Agriculture, Usmanu Danfodiyo University, P.M.B. 2346, Sokoto, Nigeria
S.G. Ado
Department of Plant Science, Faculty of Agriculture, Ahmadu Bello University, Samaru-Zaria, Nigeria
Asian Journal of Crop Science
Year: 2009 | Volume: 1 | Issue: 1 | Page No.: 15-25







ABSTRACT
Five onion cultivars were crossed in a diallel and their progenies evaluated at Sokoto and Talata Mafara, during the 2004/2005 and 2005/2006 seasons. Thirty milliliter of 10-1 cfu of spore suspension of Alternaria porri was poured into each plot. Combined analysis indicated that location, recorded highly significant mean squares (p<0.01) for disease incidence, fresh and cured bulb yields. Genotype recorded highly significant (p<0.01) mean squares for all characters. Genotypexlocation interactions recorded highly significant (p<0.01) mean squares for disease incidence and severity and fresh and cured bulb yields.
PDF Abstract XML References Citation
How to cite this article
L. Abubakar and S.G. Ado, 2009. GenotypexEnvironment Interaction for Resistance to Purple Blotch (Alternaria porri L. (Ellis) Cif.) in Onion (Allium cepa L.) in Nigeria. Asian Journal of Crop Science, 1: 15-25.
DOI: 10.3923/ajcs.2009.15.25
URL: https://scialert.net/abstract/?doi=ajcs.2009.15.25
DOI: 10.3923/ajcs.2009.15.25
URL: https://scialert.net/abstract/?doi=ajcs.2009.15.25
INTRODUCTION
Onion (Allium cepa L.) belongs to the genus Allium (Messiaen, 1994). In comparison with other fresh vegetables, onions are relatively high in food value (Hussaini et al., 2000). They contain a phytochemical quercetin, which is effective in reducing the risk of cardiovascular disease, an anticancer and has promise as an antioxidant (Smith, 2003). On a worldwide basis, onion ranks as one of the five most important fresh market vegetable crops (Cramer, 2000). In Nigeria the crop is second only to tomatoes in importance among the vegetables and is mainly grown for its bulbs (Hussaini et al., 2000). Onion is produced world wide, in 2004, 56.80, 4.26 and 1.06 million metric tonnes were produced of dry bulb for the world, Africa and West Africa and 615,000 metric tonnes for Nigeria (FAOSTAT, 2004). Similarly, in 2004, 3.09 million hectares were cultivated with onion the world over. In Africa, West Africa and Nigeria 280,059 ha, 61,160 ha and 41,000 ha, respectively were cultivated with onions in 2004 (FAOSTAT, 2004). Global average yield of onion in 2004 has been estimated at 18.3 t ha-1, with 15.21 t ha-1 for Africa, 15.187 t ha-1 for West Africa and 15 t ha-1 for Nigeria (FAOSTAT, 2004).
According to Green (1969, 1972) low onion production in Nigeria is as a result of pests and disease infestation and of the traditional methods employed in onion production by the local farmers (Kadams, 1983). Wet season trials at the Institute for Agricultural Research (IAR), Samaru, Zaria, found that low yields were associated with attack by leaf pathogens especially Alternaria porri.
Purple blotch of onion caused by A. porri is an important disease of onion worldwide except in very cool production areas (Awad et al., 1978; Everts and Lacy, 1990a; Brar et al., 1990; Aveling et al., 1993, 1994; Chaput, 1995, Cramer, 2000; Schwartz et al., 2005). It is especially troublesome in warm and humid environments (Suheri and Price, 2000). The fungus attacks both leaves and flower stalks (Bock, 1964), reducing foliar production by 62-92% (Suheri and Price, 2001). The disease can cause a yield loss of 30% (Everts and Lacy, 1990b) and 100% of the seed crop when the weather favours it (Daljeet et al., 1992; Schwartz, 2004). Purple blotch disease of onion is so important as a disease complex that nutrition (Awad et al., 1978), cultural practices (Arboleya et al., 2003), environmental conditions (Everts and Lacy, 1990a; Suheri and Price, 2000, 2001) and prevalence of other disease factors (Brar et al., 1990) all contribute to resistance or susceptibility to the disease, thereby making it more difficult to control. The most reliable measure of control of the disease so far is through crop rotation and use of resistant varieties (Delahaut and Stevenson, 2004; Latin and Helms, 2001) or good cultural practices (Schwartz, 2004; Allen, 2005). The use of resistant varieties is not only suitable to low input farming, but also reduces the consequence of chemical sprays on non target organisms and on the environment (Thomas and Waage, 1996).
Genotypexenvironment interaction (GxE) is of notable importance in the development and evaluation of new varieties and an ideal variety is one that combines high yield with stability of performance (Ado and Ishiyaku, 1999). Russell and Stuber (1985) studied genotypexphotoperiod and genotypextemperature interactions in maize using a six parent diallel mating design. They recorded significant differences between the genotypes with respect to environmental variation. McCallum et al. (2001) in New Zealand reported that firm storage onion types produced from areas in higher latitudes (intermediate to long-day) are highly pungent and have high dry matter with soluble sugars, in contrast to onions selected from lower altitude (short-day varieties) which generally have lower pungency, lower dry matter and relatively more simple sugars that impart sweetness (fructose and sucrose). They also reported that both genotype and environment affected days to 50% tops down, days to maturity, pungency and soluble solids under New Zealand conditions. They also indicated that pyruvic acid content is strongly influenced by environmental variation and the method of extraction.
The objective of our study was to determine the genotypexenvironment interactions of resistance to purple blotch disease in onion under northern Nigerian conditions. Our aim was to obtain information, which will serve as a guide in the breeding of resistant cultivars for this region, where onion is an economically important crop. For this purpose we conducted a diallel cross between five shortday cultivars choosen for their diverse responses to purple blotch and then assessed their performance in field trials at two locations over two seasons.
MATERIALS AND METHODS
Seeds of five onion varieties Red Creole (H), Kaharda (I), Koumassa (A), Sokoto Local (G) and Ori (E) were crossed in a complete diallel mating to generate diversity for resistance to purple blotch disease (Table 1) during the 2003/2004 onion growing season (October 2003-May 2004). Sokoto local was also chosen because it is the local standard cultivar. Seeds of the varieties were raised in a nursery where the soil was thoroughly mixed with farmyard manure at the rate of 5.5 t ha-1 (NAERLS, 1993). The seedlings were allowed to grow for a period of forty- nine days and later transplanted into plastic pots of 1458 cm3. The seedlings were allowed to grow to form bulbs. Bulbs generated were then cut across to encourage flowering and planted into plastic pots of the same dimension for growth up to flowering. At flowering diallel cross was made among the five varieties in all possible combinations giving rise to twenty-five progenies, including the crosses, selfs and the reciprocals.
The twenty five progenies and their parents were evaluated over two onion growing seasons (2004/2005 and 2005/2006) at two locations: Sokoto (Kwalkwalwa village; latitude 13°06′ 28 N and longitude 05°12′ 46 E) in Sokoto State and T/Mafara (latitude 12° 13′ 18 N and longitude 06°05′ 05 E and altitude 1150 m) in Zamfara State of Nigeria. In each field trial the genotypes were laid out in a Randomised Complete Block Design (RCBD) and replicated three times at each location and growing season. The total farm area was 333 m2 per season, with a plot size of 1.8x1.5 m of sunken beds.
| Table 1: | Description of the five onion varieties used in the study |
 | |
The genotypes were:
1 = A, 2 = E, 3 = G, 4 = I, 5 = H, 6 = AxE, 7 = AxG, 8 = AxI, 9 = AxH, 10 = AxA, 11 = ExA, 12 = ExG, 13 = ExI, 14 = ExH, 15 = ExE, 16 = GxA, 17 = GxE, 18 = GxI, 19 = GxH, 20 = GxG, 21 = IxA, 22 = IxE, 23 = IxG, 24 = IxH, 25 = IxI, 26 = HxA, 27 = HxE, 28 = HxG, 29 = HxI and 30 = HxH
Seeds for all the experiments were sown on 15th of October of each year in a nursery. Farmyard manure at the rate of 5.5 t ha-1 (NAERLS, 1993) was thoroughly incorporated into the soil of the nursery beds, thereafter sunken beds of 2x1 m dimensions were made. The soil was made into fine tilth after removing large stones and stumps and watered then left for two days. Seeds of the genotypes for evaluation were separately broadcast into the sunken beds and mulched with millet stalks and irrigated. One week after germination the mulch materials were removed from the beds. The seedlings were thereafter watered (irrigated) in the evenings at two days interval initially and later at five days interval until the time of transplanting.
Transplanting of the seedlings was carried out forty nine days after sowing (7 WAS). The seedlings were removed from the seedbeds after watering to moisten the soil. At the field sites the seedlings were transplanted at a spacing of 30 cm between rows and 15 cm within rows. Each plot consisted of six rows of ten plants per row, the genotypes being evaluated were planted in the two middle rows of each plot and the other four rows were planted with a guard row onion variety (Aleiro).
No fertilizer was applied to the fields because according to Awad et al. (1978) application of nitrogen to onion plants increases susceptibility to purple blotch disease due to the production of succulent leaves, while addition of potassium and calcium super phosphate improves resistance to purple blotch.
Soil tests were conducted at both locations and for both seasons. The results of the soil analysis indicate that the soil at Sokoto is sandy loam while at Talata Mafara the soil is loamy sand.
The inoculation of the field was carried out at two weeks after transplanting allowing the seedlings to overcome the transplanting shock and also close enough to 10 WAS when the varieties used in the study were at the 5-7 leaf stage. This is in accordance with Arboleya et al. (2003) who reported that plants should be inoculated at 5-7 leaf stage. Thirty milliliters of 10-1 cfu of the spore suspension of Alternaria porri was poured in the centre of each plot immediately after irrigation.
The first three irrigations of the fields after transplanting were carried out every four days. Thereafter irrigations were maintained at 5 day interval up to harvest. At Sokoto irrigation was by flooding method using water pump, while at Talata Mafara irrigation was by flooding using the gravity method, using water from canals constructed by the Bakolori irrigation project.
No sprays of any kind were carried out since the genotypes were being evaluated for fungal infection. Three weeding regimes were carried out during each season at each location. The first weeding was carried at 10 WAS, the second at 14 WAS and the last at 19 WAS. Weeding was done manually by hoeing.
Harvesting was carried out when more than 50% of the tops were down for all the materials. The crops were carefully harvested using hoes to bring the bulbs to the surface of the soil, while the upper parts of the plants were cut with knives and sickles to separate the bulbs from the tops level with the neck. The harvested bulbs were spread on the floor in a ventilated room and allowed to dry for 10 days.
Data on fresh bulb yield (kg ha-1), cured bulb yield (kg ha-1), average bulb weight (g) were determined by weighing ten bulbs. Bulb diameter was determined using a vernier calliper for ten bulbs and days to maturity was recorded when 50% tops were down for each plot. Number of leaves/plant were counted at maturity. Disease incidence (%) and disease severity were assessed fortnightly. Disease incidence was determined according to Tarr (1981):
Diseased plants were plants that had sunken spots on leaves, which later enlarged to become purple with a yellow halo and elongated destroying the leaf tissue and eventually causing the bulb to rot. Disease severity was determined for each plot on the basis of standard procedures recommended by the International Plant Genetic Resource Institute, Rome, Italy. The rating was in the following order: 1 = Highly resistant, 2 = Resistant, 3 = Moderately resistant, 4 = Susceptible and 5 = Highly susceptible (IPGRI et al., 2001).
Data of the experiments were statistically analysed using the Statistical Analysis Systems (SAS, 1996) computer package. The statistical model used for the combined analysis over seasons and locations was a mixed model given by Obi (1986) as:
Yijkl = μ + Gi + Lj + Sk + RL + (GL)ij + (GS)ik + (LS)jk + (GLS)ijk + eijkl |
Where:
| Yijkl | = | The observation on ith genotype in jth environment in kth replication |
| μ | = | The general mean |
| Gi | = | The effect of genotypes |
| Lj | = | The effect of location |
| Sk | = | The effect of season |
| RL | = | The effect of replication within season and location |
| (GL)ij | = | The effect of genotypexlocation interaction |
| (GS)ik | = | The effect of genotypexseason interaction |
| (LS)jk | = | The effect of locationxseason interaction |
| (GLS)ijk | = | The effect of genotypexlocationxseason interaction |
| eijkl | = | The error effect associated with ijklth observation |
The components of variance were estimated from the mean squares for each character by using the observed mean squares, thus with reference to Table 2 and 3.
| Table 2: | Form of Analysis of variance (ANOVA) table for single experiment (one season one location) |
 | |
| r = No. of replications, g = No. of genotypes, δ2e = Error variance, δ2g = Total genetic variances among the genotypes MS subscript: The observed mean squares of the subscript effect | |
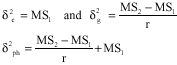 |
RESULTS AND DISCUSSION
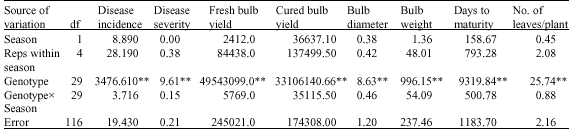
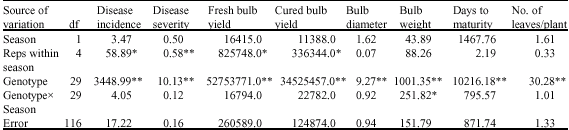
Combined analysis of variance for the two seasons at Sokoto indicated highly significant genotypic mean squares for all the characters (Table 4). The season and genotype x season interaction were not significant for all the characters. At Talata Mafara, the genotypic variances were highly significant for all the characters while the genotype x season interaction indicated significant mean squares for bulb weight only (Table 5).
Highly significant mean squares were observed for the genotypes for all the traits during the 2004/2005 season across the locations (Table 6). The analysis also indicated highly significant mean squares for replications within location for disease incidence and disease severity and significant mean squares for fresh bulb yield. Fresh and cured bulb yields recorded highly significant mean squares for location, while disease incidence recorded significant mean squares for location during the season.
During the 2005/2006 season, however, the combined analysis across locations indicated highly significant genotypic mean squares for all the traits. Location effect was highly significant for fresh and cured bulb yields, while disease incidence recorded significant mean squares (Table 7). The genotypexlocation interactions were not significant for all the traits measured.
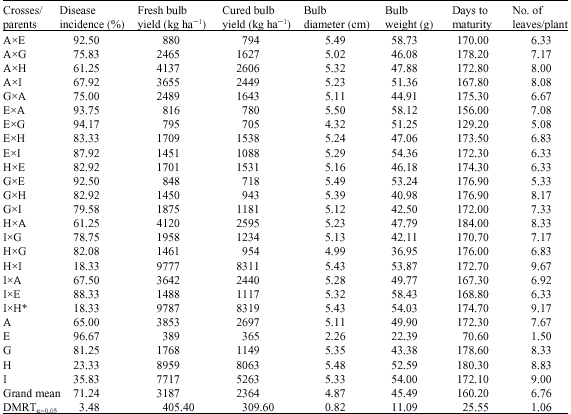
The performance of the genotypes and their parents across seasons and locations (Table 8), indicated cultivars Koumassa, Kaharda and Red Creole having lower mean disease incidence and higher mean fresh and cured bulb yields than their respective grand means. Crosses AxH, AxI, HxA, HxI, IxA and IxH recorded lower mean disease incidence than their grand mean, while crosses AxH, AxI, HxA, HxI, IxA and IxH recorded greater mean fresh and cured bulb yields than their grand mean.
Combined analysis across seasons and locations indicated highly significant (p = 0.01) mean squares for the genotypes for all the characters (Table 9). Highly significant mean squares were also recorded for the genotypexlocation for disease incidence and disease severity and fresh and cured bulb yields. Similarly, highly significant location effects were recorded for disease incidence and fresh and cured bulb yields. The genotypexseasonxlocation interactions were, however, not significant for all the traits. Similarly the seasonal effects, the season by location and the genotype by season interactions were not significant for all the traits.
According to Lamkey (2006), plant breeders evaluate germplasm in environments to identify genotypes that exhibit optimal adaptation to the needs of society, the demands of nature and the desires of the market place. The combined analysis of the experiments indicated significant variation among the parents and the crosses evaluated for all the traits under study (Table 9). Both superior and inferior genotypes exist in the population, with respect to the characters under consideration. This therefore suggests that selection for these traits during breeding programmes is possible.
The combined analysis of variance across seasons and locations also indicate that location, genotypes and genotypexlocation interaction were the sources of variation that accounted for the variability observed with respect to disease incidence (Table 9).
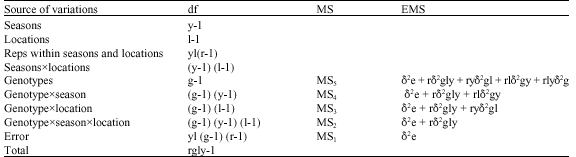
| Table 3: | Form of combined Analysis of variance (ANOVA) across seasons and locations |
 | |
| Table 4: | Combined analysis of variance for eight onion characters grown at Sokoto during the 2004/2005 and 2005/2006 seasons |
 | |
| *: Significant at 5%, **: Significant at 1% levels of significance | |
| Table 5: | Combined analysis of variance for eight onion characters grown at Talata Mafara during the 2004/2005 and 2005/2006 seasons |
 | |
| *: Significant at 5%, **: Significant at 1% levels of significance | |
| Table 6: | Combined analysis of variance for eight onion characters grown at Sokoto and Talata Mafara during 2004/2005 season |
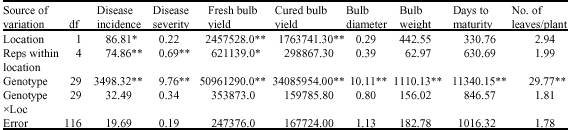 | |
| *: Significant at 5%, **: Significant at 1% levels of significance | |
| Table 7: | Combined analysis of variance for eight onion characters grown at Sokoto and Talata Mafara during 2005/2006 season |
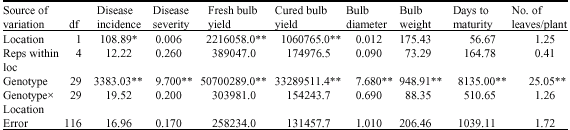 | |
| *: Significant at 5%, **: Significant at 1% levels of significance | |
| Table 8: | Mean performance of onion crosses and parents inoculated with A. porri grown at Sokoto and Talata Mafara during 2004/2005 and 2005/2006 seasons |
 | |
| Table 9: | Combined analysis of variance of onion genotypes inoculated with A. porri at Sokoto and Talata Mafara for two seasons |
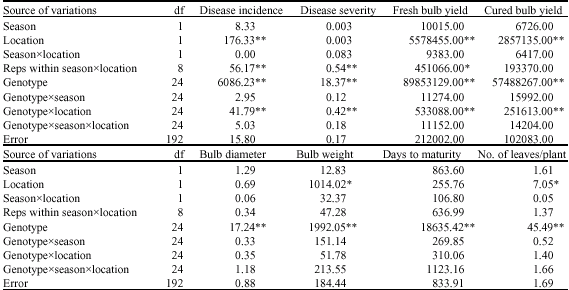 | |
| *: Significant at 5%, **: Significant at 1% levels of significance | |
This suggests that the two locations differed in disease incidence score for the genotypes. The differences between the locations in environmental conditions most especially during the harvest months of March and April (Table 10) are the likely reason for the location effect. The highly significant genotypexlocation interaction for disease incidence indicate disease incidence among the genotypes vary with location. In other words, the ranking of the genotypes for the character vary with location.
| Table 10: | Meteorological data for harvest period at Sokoto and Talata Mafara during 2004/2005 and 2005/2006 seasons |
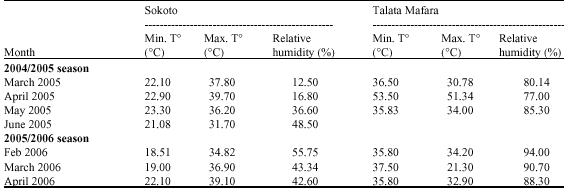 | |
| Source: Sokoto Energy Research Center, Sokoto and Institute for Agricultural Research meteorological station Talata Mafara | |
According to Everts and Lacy (1990b) the disease can cause a yield loss of 30%, while Daljeet et al. (1992) and Schwartz (2004) reported yield loss of 100% of the seed crop when the weather favours the disease. The severity of purple blotch disease is greatly influenced by location (environment), nutrition (Awad et al., 1978), cultural practices (Arboleya et al., 2003), environmental conditions (Everts and Lacy, 1990a; Suheri and Price, 2000, 2001) and prevalence of other disease factors (Brar et al., 1990), all of which contribute to resistance or susceptibility to the disease. The influence of the environment on the development of purple blotch has been reported by several authors. Green (1972), reported that wet season trials of onions at IAR Samaru, Nigeria were associated with low yields which was attributed to attack by leaf pathogens especially Alternaria porri (Ell). In our trials disease severity was significant for genotype and genotypexlocation interactions suggesting that the pattern of location effect on disease severity was similar to that of disease incidence as expected. For both traits, the magnitude of the genotypic variance is higher than the first order interaction variance. The second order interactions were not significant for any of the measured traits. Breeders and farmers have long known that the best variety in one season in a sample of ten similar locations might not be the best in another season or when averaged across several seasons at the same locations (Lamkey, 2006). Unless therefore GxE is dealt with effectively, the potential genetic gains of plant breeding programmes will not be realized and delivered to the market place. According to Van Eeuwijk (2006), while experimental error can complicate characterization of genotypic performance, it can be reduced by experimental design and/or analytical methodologies. Within - location replication is also useful, since it allows separation of GxE from experimental error, thereby enabling better characterization of GxE. Unfortunately, GxE interaction can not be reduced or mitigated by design or analysis methods because GxE is an inherent attribute of the given genotypes in the given environment.
The combined analysis also indicated that fresh and cured bulb yields were highly significantly influenced by location, genotypes and genotypexlocation interaction. Breeding for resistance to purple blotch and for fresh and cured bulb yields should take cognizance of these factors. Breeding for fresh and cured bulb yields in onions should therefore be location specific. Similar conclusions were reported by other workers (Jones and Mann, 1963; Purseglove, 1972; Bednarz and Olarewaju, 1986). According to Van Eeuwijk (2006), in the absence of GxE and with experimental error at reasonably low levels, average phenotypic performance across environments provides a good representation of genotypic performance. Consequently, relative performance of genotypes can be determined from differences in these phenotypic performances. However, in the presence of significant GxE interaction, relative genotypic performance can only be characterized for specific environments.
Location effects were also significant for bulb weight and number of leaves/plant while genotypic differences for the traits were highly significant. The result therefore suggests that location and genotypes are two important factors to consider when selecting for bulb weight and number of leaves/plant in onions. The results indicate a need for location specificity when breeding for fresh and cured bulb yields in onion. Bulb diameter and days to maturity did not show any significant differences for seasons, locations and their interactions suggesting that the determining factor when selecting for these traits is the genotype. McCallum et al. (2001), however, in New Zealand, working on locally- adapted longer-day onions, reported that both genotypes and environment affect days to 50% tops down (days to maturity), pungency and soluble solids. Differences between their findings and ours may be connected with the fact that all of our onion cultivars were short-day and relatively well-adapted to our locations. Also, the cultural practices for the two sets of experiments may not have been the same during the cultivation of the crop.
The variations observed between different research groups may be connected with environmental variations. GxE interaction can be used to describe differential genotypic response to various geographic locations in a given year. There is also a temporal component to GxE, since the same geographic location will have a different environment in different years and at different times of the year (e.g., effects of planting date and growing season). In order to deal explicitly with both the spatial and the temporal aspects of environment, the GxE is separated into genotypexlocation, genotypexseason and genotypexseasonxlocation (Van Eeuwijk, 2006).
CONCLUSIONS
From our research we conclude that genetic diversity in short-day onions with respect to resistance to purple blotch disease exists and that breeding for resistant cultivars is therefore possible. Location, genotype and genotypelocation interactions were shown to influence disease incidence, fresh and cured bulb yields all of which are characters important in resistance to the disease. In future West African breeding programmes for resistance to purple blotch disease therefore these factors should be considered.
ACKNOWLEDGMENTS
We wish to acknowledge the contributions of Dr. Lesley Currah, who assisted in obtaining the seeds of Red Creole from Seminis Vegetable Seeds and also for her scientific contribution to this work. We also wish to acknowledge the authorities of ICRISAT, Sadore, Niger Republic, INRAN and ONAHA all in Niger, Republic for the seeds of their cultivars.
REFERENCES
- Aveling, T.A.S., H.G. Snyman and F.H.J. Rijkenberg, 1994. Morphology of infection of onion leaves by Alternaria porri. Can. J. Bot., 72: 1164-1170.
CrossRefDirect Link - Cramer, C.S., 2000. Breeding and genetics of Fusarium basal rot resistance in onion. Euphytica, 115: 159-166.
CrossRefDirect Link - Daljeet, S., J.S. Dhiman, A.S. Sidhu and H. Singh, 1992. Current status of onions in India: Strategies for disease resistance breeding for sustained production. Onion Newslett. Trop., 4: 43-44.
Direct Link - Hussaini, M. and E.B. Amans, 2000. Yield, bulb size distribution and storability of onion (Allium cepa (L.) under different levels of N fertilization and irrigation regime. Trop. Agric., 77: 145-149.
Direct Link - McCallum, J.A., D.G. Grant, E.P. McCartney, J. Scheffer, M.L. Shaw and R.C. Butler, 2001. Genotypic and environmental variation in bulb composition of New Zealand adapted onion (Allium cepa) germplasm. N. Z. J. Crop Hortic. Sci., 29: 149-158.
CrossRef - Russell, W.K. and C.W. Stuber, 1985. Genotype photoperiod and Genotype Temperature interactions for maturity in maize. Crop Sci., 25: 152-158.
Direct Link - Suheri, H. and T.V. Price, 2000. Infection of onion leaves by Alternaria porri and Stemphylium vesicarium and disease development in controlled environments. Plant Pathol., 49: 375-382.
CrossRef - Suheri, H. and T.V. Price, 2001. The epidemiology of purple blotch on leeks in Victoria, Australia. Eur. J. Plant Pathol., 107: 503-510.
CrossRef