
Ikuo Nishigaki
NPO International Laboratory of Biochemistry, 1-166 Uchide, Nakagawa-Ku, Nagoya 454-0926, Japan
Balasubramanian Rajkapoor
NPO International Laboratory of Biochemistry, 1-166 Uchide, Nakagawa-Ku, Nagoya 454-0926, Japan
Ramachandran Venugopal
NPO International Laboratory of Biochemistry, 1-166 Uchide, Nakagawa-Ku, Nagoya 454-0926, Japan
Palaniyandi Senthilnathan
NPO International Laboratory of Biochemistry, 1-166 Uchide, Nakagawa-Ku, Nagoya 454-0926, Japan
Hao Qian Zhang
Department of Biochemistry, Graduate School of Medicine, Nagoya University, Tsurumai-cho, Showa-ku, Nagoya 466 8550, Japan
Dhanapal Sakthisekaran
Department of Medical Biochemistry, University of Madras, Taramani Campus, Chennai 600-113, India
Asian Journal of Cell Biology
Year: 2007 | Volume: 2 | Issue: 2 | Page No.: 34-42







ABSTRACT
Protective effect of fresh apple skin extract on Human Umbilical Vein Endothelial Cells (HUVEC) against glycated protein (GFBS)-iron chelate (FeCl3) induced toxicity was studied. HUVEC incubated in glycated protein either alone or combined with iron chelate showed a significant (p<0.001) elevation of lipid peroxidation (LPO) accompanied by depletion of superoxide dismutase (SOD), catalase, Glutathione Peroxidase (GPx) and Glutathione Reductase (GR). Treatment of HUVEC with apple extractat a concentration of 50 mg showed a significantly decreased level of LPO with concominant increase of antioxidant levels to near normal. Morphological changes of HUVEC were compared with respective control. Our finding indicate that apple skin extract has protective effect against glycated protein-iron chelate induced toxicity, which suggests that apple extractcould exert a beneficial effect on preventing diabetic angiopathies.
PDF Abstract XML References Citation
How to cite this article
Ikuo Nishigaki, Balasubramanian Rajkapoor, Ramachandran Venugopal, Palaniyandi Senthilnathan, Hao Qian Zhang and Dhanapal Sakthisekaran, 2007. Protective Effect of Fresh Apple Skin Extract on Human Umbilical Vein Endothelial Cells Against Glycated Protein-iron Chelate Induced Toxicity. Asian Journal of Cell Biology, 2: 34-42.
DOI: 10.3923/ajcb.2007.34.42
URL: https://scialert.net/abstract/?doi=ajcb.2007.34.42
DOI: 10.3923/ajcb.2007.34.42
URL: https://scialert.net/abstract/?doi=ajcb.2007.34.42
INTRODUCTION
Vascular endothelial cells and vascular muscle cells are the two major components of the vessel wall. Since endothelial cells are situated within the intimal layer of the vessel wall, abnormal levels of glucose in blood may trigger initial damage leading to functional and structural alterations, which are potentially responsible for diabetic complications (Xu et al., 2004). High glucose concentration has been reported to be toxic in vitro for endothelial cells, as represented by retarded cell proliferation (Curcio and Ceriello, 1992) disturbed cell cycle (Lorenzi et al., 1985) increased DNA damage (Lorenzi et al., 1986) and slightly accelerated cell death (Lorenzi et al., 1985). Such pathological changes may play important role in causing microvascular lesions that are common and prominent in diabetes. Thus prevention of hyperglycemia-triggered endothelial cell lesions may prevent diabetes associated microvascular complications. The Maillard or Browning reaction between reducing sugars and protein is proposed to be involved in the pathophysiological procedure of aging and diabetic complications (Brownlee et al., 1985).
The scavenging function of antioxidantenzymes is reduced by glycation of their free amino terminalsto produce Advanced Glycated End-product (AGE) variants ofthese enzymes. High blood glucose level causes binding of glucose covalently and non enzymatically to various proteins in vivoto generate AGEs (Szalecsky et al., 1999). Glycation can alter the structure as well asthe function of physiological proteins. Therefore, hyperglycemiamay modify the properties of the enzymatic antioxidant defenseproteins and increase the susceptibility of diabetic patient to vascular damage due to excessive production of free-radicalspecies.
The levels of AGE increased in the serum of patients with diabetes. AGEs have been linked with retinopathy,nephropathy and neuropathy, as well with largevessel disease (Nessar, 2005; Sara et al., 2003; Thomas et al., 2003). The mechanisms leading to micro-and macro-angiopathyare thought to occur by an interaction of AGE with a receptor(known as RAGE) expressed on the cell surface membrane of vascularendothelial cells (Sensi et al., 1995).It is therefore interesting to speculate that an inhibition of AGE synthesis, or its antagonism at the level of the RAGE receptor,could play a role in preventing the genesis of vascular diabeticcomplications.
Oxidative stress has emerged as a strong pathogenic cofactor in the development of longterm complications of type II diabetes, such as atherosclerosis, nephropathy and retinopathy (Jain and Ajay, 2004). The H2O2 is the most stable of the Reactive Oxygen Species (ROS) and is produced from the dismutation of superoxide and form the divalent reduction of oxygen by various oxidases. H2O2 is detoxified by catalase and by glutathione peroxidases. In general, the toxicity caused by superoxide and by H2O2 in a biological systems is believed to be due to the conversion of these ROS to more potent oxidants, such as the hydroxyl radical or ferryl or perferryl species, with the oxidizing power of the hydroxyl radical. This conversion is either catalyzed or actually by the presence of transition metals, such as iron or copper and it is believed that the toxicity of iron in biologic systems is due to iron-catalyzed production of the above-mentioned potent oxidants by Fenton or Haber-Weiss types of reactions (Jagetia et al., 2004; Halliwell and Gutteridge, 1983). Results from numerous studies have shown that iron potentiates the toxicity of drugs, redoxcycling agents and prooxidants, whereas removal of iron protects the system against toxicity. Although identification of the low molecular weight, nonheme iron pool in cells has proved elusive, cellular sources of iron clearly potentiate oxidative stress.
The level of glycated protein is known to increase in diabetic patients and the Fructosamine Value (FV) of serum is regarded as an indicator of blood glucose level. Yagi et al. (1985)have reported that chinoform-ferric iron chelate causes lipid peroxidation in cultured neural retinal cells and Nishigaki et al. (1998) have shown that glycated protein-iron chelate provokes lipid peroxidation in endothelial cells, these facts explain at least in part the mechanism of atherogenesis in diabetic patients.
Epidemiological studies have shown that a diet rich in fruits and vegetables has a beneficial preventive effect against cardiovascular diseases and cancer by mechanisms that have not yet been elucidated. These phenolic compounds are widely found in many berries, red grapes, purple sweet potatoes, red cabbages and other pigmented foods, plants and vegetables (Harborne, 1988). While the strong antioxidant potential of these molecules is well characterized (Hou, 2003).
It is generally assumed that the active dietary constituents contributing to these protective effects are antioxidant nutrients such as α-tocopherol and β-carotene. However, recent investigations highlight an additional role of polyphenolic components of higher plants that may act as antioxidants or via other mechanisms contributing to the anticarcinogenic or cardioprotective actions (Kanner et al., 1994; Tedesco et al., 2001). In particular, some beverages, such as wine and tea, have been shown to elicit antioxidant properties in both in vitro and in vivo systems (Huang and Ferrano, 1992; Newmark, 1992). In this research we have demonstrated that fresh apple skin extract produce a significant protective effect against glycated protein-iron chelate induced toxicity in human umbilical vein endothelial cells in vitro.
MATERIALS AND METHODS
Cells and Chemicals
Fresh apples were obtained from a localfarm. HUVEC and endothelial medium (EG2) were purchased from Kurabo (Japan). Catalase assay kit was obtained from Cayman Chemical Company (USA). Glutathione peroxidase and glutathione reductase assay kits were purchased from Oxi Research (USA) and Trevigen Inc (USA), respectively. Superoxide dismutase was obtained from Dojindo Laboratories (Japan). Protein assay kit was from Bio-RAD, USA. Cell culture flask was from Nunc, Denmark. All other chemicals used were of analytical grade and water was purified through Millipore Milli Q system (Millipore Co., USA).
Preparation of Apple Extract
Fresh apple extract was preparedfrom Gala apples by adding 1 mL of alcohol per g ofskin and blended at high speed. The blendedhomogenate was strained, centrifuged and filter-sterilized.The extracts were then aliquoted and stored at -20°C.
Serum Glycation
For glycation of Fetal Bovine Serum (FBS), 20 mL of FBS was incubated with 50 mM glucose in 1 mL of 67 mM phosphate buffer (pH 7.4) at 37°C for 25 days under sterile conditions. After incubation, the mixtures was dialysed against the same buffer at 4°C. The dialysate was used as glycated FBS after concentration by using aquacide II (Calbiochem-Novabiochem Corp., USA). The extent of glycation of FBS was assayed with a commercial kit (Nippon Roche, Japan) and expressed as Fructosamine Value (FV). The FV of GFBS prepared were 12 mM and that of original FBS was 0.20 mM.
Cell Culture and Experiments
HUVEC were grown in endothelial medium-2 (EG2), supplemented with 10% (v/v) fetal bovine serum, gentamicin sulphate (50 mg mL-1)/amphotericine-B (50 μg mL-1) in addition to human recombinant fibroblast growth factor-B (hFGF-B; 5 μg mL-1), human recombinant epidermal growth factor (hEGF; 10 μg mL-1), hydrocortisone (1 mg mL-1) and heparin (10 mg mL-1). The cells were grown in 25 cm2 flasks. The cultures were maintained in a humidified atmosphere containing 5% CO2 at 37°C, until reaching 80% confluence and used for experiments between passage 3 and 4.
The experimental design was formed by 6 groups with 5 flasks in each group. Group I was treated with 15% of FBS. The group II, III and IV were treated with GFBS (70 μM), FBS+FeCl3 (20 μM) and GFBS+FeCl3, respectively. The groups V and VI were treated with FBS+FeCl3 (20 μM) + apple extract (50 mg) and GFBS+FeCl3 + apple extract (50 mg), respectively. After 24 h incubation, the cells were collected from all the experimental groups by detaching from the flasks using cell scraper-M (Sumitomo bakelite Co., Japan) with phosphate buffer and centrifuged at 1000 x g for 10 min at 4°C. The cell pellet were suspended with cold phosphate buffer and homogenized by using sonicator (Misonix incorporated, USA). The mixture was again centrifuged at 10,000 x g for 15 min at 4°C. A portion of the supernatant was used for the assay of lipid peroxidation and antioxidant enzymes activities.
Protein Assay
Protein content was quantified by means of the BIO-RAD reagent protein assay kit, following the method of Lowry et al. (1951).
Lipid Peroxidation Assay
Lipid peroxidation was determined by using the assay described in Kyowa kit (Japan) manual. In brief, the LPO assay is based on the reaction of a chromogenic reagent, N-methylcarbamoyl-3, 7-dimethyl amino-10H-phenothiazine with ascorbic acid oxidase and cumene hydroperoxide at 30°C. The stable chromophore with maximal absorbance at 675 nm and LPO activity were expressed as nmol MDA mL-1 mg-1 protein.
Catalase Activity
Catalase (CAT) activity was determined using Cayman chemical company kit. After addition of 10 mM H2O2, cell lysates were incubated for 20 min and mixed with chromogen substrate followed by the addition of stopping reagent. The colour developed during incubation was detected at 540 nm using microplate reader (ImmunoMini NJ). The rate of absorbance was converted to nmol/min/ml of enzyme activity, determined from the linear regression of standard curve using CAT.
Superoxide Dismutase Activity
SOD activity was determined by using the method described in Dojindo laboratories SOD kit manual. In brief, cell lysates were treated with Water Soluble Tetrazolium salt (WST) and SOD activity (% inhibition) was determined by measuring the absorbance at 450nm using microplate reader.
Glutathione Peroxidase Activity
Glutathione peroxidase activity was determined by using the method described in Oxi research GPx-340 kit. After the addition of 350 μL of NADPH to cell lysates, the reaction was initiated by the addition of tert-butyl hydroperoxide. The changes in absorbance were recorded at 340 nm using spectrophotometer (Jenway, UK).
Glutathione Reductase Activity
Glutathione reductase activity was determined using the assay described in Trevigen kit. In brief, cell lysates were treated with oxidized glutathione and changes were recorded at 340 nm after the addition of nicotinamide adenine dinucleotide phosphate (NADPH). The glutathione reductase activity was expressed in terms of the oxidation of NADPH mU mL-1.
Morphological Observation by Phase Contrast Microscopy
To examine morphological changes in endothelial cells by phase contrast microscopy, we observed the cells 24 h after incubation in the presence or absence of GFBS, FeCl3 and apple extract using a Carl Zeiss Axio vision electron microscope (Germany).
Statistical Analysis
The results were expressed as mean±SD and the statistical significance was analyzed by one way ANOVA followed by Tukey multiple comparison test (GraphPad InStat, USA). p-values<0.05 were considered significant.
RESULTS
Lipid Peroxidation
The lipid peroxide levels were measured as MDA concentration and expressed as nmol mL-1 mg-1 protein (Table 1). Treatment of HUVEC with GFBS-iron chelate elevated lipid peroxidation levels significantly when compared with FBS treatment group (p<0.001). The treatment of HUVEC with 50 mg of apple extractsignificantly (p<0.001) reduced the iron chelate induced lipid peroxidation.
Antioxidant Enzymes
GFBS-iron treatment significantly reduced the SOD and catalase activity in HUVEC when compared with the control group; however, when HUVEC was treated with apple extract elevated the cellular SOD and catalase levels when compared with FBS control and GEBS+FeCl3 groups, respectively (Table 1).
| Table 1: | Effect of apple skin extract on LPO and antioxidant levels in HUVEC against glycated protein-iron chelate induced toxicity |
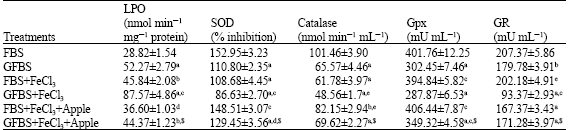 | |
| Average of 5 determinations; Values are expressed as mean±SEM. ap<0.001; bp<0.01 vs FBS; cp<0.001; dp<0.01; ep<0.05 vs GFBS. $p<0.001 vs GFBS+FeCl3. Data were analyzed by using one way ANOVA followed by Tukey multiple comparison test | |
 | |
| Fig. 1: | Phase contrast micrographs of HUVEC treated with GFBS-iron chelate and apple extract, (A) Control cells cultivated in the medium containing 15% FBS alone, (B) GFBS treated cells show injury of cells, degeneration and shrunken and individual cell necrosis, (C) FBS and FeCl3treated cells show degeneration and individual cell necrosis, (D) GFBS and FeCl3treated cells show massive necrosis and expanded intercellular space and shrinkage of cells, (E) FBS and FeCl3 +Apple extract treatment show reduced cell injury and recover from the iron toxicity and (F) GFBS and FeCl3+Apple extract treatment shows quenching of GFBS-iron induced toxicity and the recovery from massive cell necrosis |
The GFBS-iron chelate significantly reduced the GPx levels in HUVEC when compared with the control group (p<0.001). Treatment of HUVEC with 50 mg of apple extract significantly altered the GPx levels to near normal when compared with GFBS-iron treated group (Table 1).
The concentration of glutathione reductase has been expressed as mU mL-1 (Table 1). Treatment of HUVEC with GFBS-iron depleted GSH concentration significantly (p<0.001) when compared with the FBS treated group. Treatment of HUVEC with 50 mg of apple extract significantly elevated the cellular glutathione levels when compared to GFBS-iron treated group (p<0.001).
Morphological studies showed that GFBS and GFBS+FeCl3 produced injury to endothelial cells, massive necrosis with intercellular space was expanded due to shrinkage of cells. Treatment with apple extract reduced the cell injury and recovered the cells from the massive necrosis induced by GFBS-iron toxicity. All these results indicate the protective potential of the extract (Fig. 1).
DISCUSSION
Endothelial dysfunction has been proved to mediate the initiation and development of many kinds of diabetic angiopathies. Disorders of contraction and relaxation, the basic function of endothelium is caused by decreased NO release. Since endothelial cells were the intimal layer of vessel wall, it was proposed to be the prominent target of AGEs and other toxic factors in diabetes state. Hyperglycemia may exert toxic effect on endothelium through the formation of AGEs.
Well-established risk factors for the development of atherosclerosis such as hyperlipidemia, hyperglycemia, hypertension, hyperhomocysteinemia and local hemodynamic stress are known to mediate elevated levels of ROS in the vasculature (Ross, 1999). The molecular and cellular changes involved in linking these diverse risk factors to a common mechanism are unclear. Increase in ROS may affect four fundamental mechanisms that contribute to atherosclerosis: endothelial cell dysfunction, Vascular Smooth Muscle Cell (VSMC) growth, monocyte migration and oxidation of LDL (Alexander, 1995).
Lipid peroxidation has been used as an indirect measure of oxidative stress. The end products of stable aldehydes react with thiobarbituric acid (TBA) to form thiobarbituric acid-malondialdehyde adduct (Berliner and Heinecke, 1996).Treatment of HUVEC with apple extract significantly (p<0.001) reduced lipid peroxidation. Hence it is likely that apple extract has an antioxidant property.
Glutathione a tripeptide and an essential bio-factor synthesized in all living cells. It functions mainly as an effective intracellular reductant (Beckman and Ames, 1998).It protects cells from free radical-mediated damage caused by drugs and also by ionizing radiation. It forms an important substrate for GPx and several other enzymes, which are involved in free radical scavenging. Apple extract inhibits GFBS-iron chelate induced reduction in glutathione levels in HUVEC.
Cells are equipped with an impressive repertoire of antioxidant enzymes, such as superoxide dismutase, which hasten the dismutation of O−2 to H2O2 and catalase and glutathione peroxidase, which convert H2O2 to water (Fridovich, 1986). SOD brings the first line of defense against free radicals by dismutating toxic superoxide into a less toxic hydrogen peroxide. SOD works in conjugation with other H2O2 removing enzymes. Also SOD is required for the growth of aerobes without excessive DNA damage due to superoxide. Selenium containing GPx decomposes H2O2 and other peroxides and initiates free radical chain reaction. Catalase heme enzyme brings the decomposition of high amounts of H2O2 and other peroxides. SOD, GPx and catalase, by a concerted action, protect the oxidative attack of superoxide and hydrogen peroxide in the cells. Apple extract elevates cellular catalase and SOD levels accompanied by an arrest of iron-chelate induced depletion of SOD, GPx and catalase in HUVEC.
Diabetes-associatedhyperglycemia causes intracellular oxidative stress, which contributes to vascular dysfunction (Baynes, 1991). The effects of hyperglycemia on endothelial cell function can be mediated by several pathways: (a) production of ROS (b) accumulation of sorbitol (c) nonenzymatic glycoxidation of macromolecules and (d) direct activation of protein kinase C (Cines et al., 1998). Glycoxidation of proteins and lipids occur ubiquitously in patients with diabetes and it is irreversible. Its consequences are especially relevant to long-term vascular dysfunction. The initial glycoxidation of proteins results in the formation of early glycation products. The interaction of AGEs with cell surface receptors (RAGEs) has been shown to generate ROS and to decrease the levels of reduced gluthatione. GSH is a major endogenous antioxidant, the level of which declines during cardiovascular diseases (Sian, 1994).Several groups have shown that GSH depletion causes various biochemical and pathological changes.
Two mechanisms have been proposed to explain how hyperglycemia causes increased ROS formation (Wolff and Dean, 1987; Hunt et al., 1993). One mechanism involves the transition metal catalyzed autoxidation of protein-bound Amadori products which yields superoxide and hydroxyl radicals and highly reactive dicarbonyl compounds. The other mechanism involves the transition metal-catalyzed autoxidation of free sugars, which also yields dicarbonyl compounds and superoxide and hydroxyl radicals. Autoxidation of protein-bound Amadori products appears to be the most important source of free radicals under near physiologic conditions (Hunt et al., 1993).
The demonstration that an ROS-dependent process plays a central role in the generation of intracellular AGEs and that inhibition of oxidant pathways prevents intracellular AGE formation have important implications for pharmacologic attempts in preventing diabetic complications. Currently, AGE formation can be reduced both by lowering hyperglycemia (Hammes et al., 1995) and by blocking reactive AGE precursors with drugs such as aminoguanidine (Brownlee, 1995). Both the interventions are effective in preventing diabetic complications in animal models. Blocking intracellular AGE formation by antioxidants offers an additional strategy for the potential prevention of diabetic complications and this point deserves further exploration.
In vitro capacity of apple extract, to protect the endothelium against the insult induced by reactive oxygen species. The results show that the apple extract has antioxidant properties in a cellular environment, at a concentration of 50 mg, which is devoid of toxicity to the endothelial cells. Treatment with apple extract reduced cell injury and absence of necrosis is an indication of its protective efficiency.
In conclusion, these results suggest that the apple extract may have some beneficial effect to the diabetic patient, since the apple extract showed to be able to prevent oxidative damage to the endothelial cells and so it may prevent or slow-down the development of vascular complications. It would be necessary to perform more assays to assess the possible protective effect of the precursors of apple extract with specific determination of their metabolites and their clinical evaluation.
REFERENCES
- Alexander, R.W., 1995. Theodore Cooper memorial lecture. Hypertension and the pathogenesis of atherosclerosis: Oxidative stress and the mediation of arterial inflammatory response: A new perspective. Hypertension, 25: 155-161.
Direct Link - Baynes, J.W., 1991. Role of oxidative stress in development of complications in diabetes. Diabetes, 40: 405-412.
CrossRefPubMedDirect Link - Beckman, K.B. and B.N. Ames, 1998. The free radical theory of aging matures. Physiol. Rev., 78: 547-581.
PubMed - Berliner, J.A. and J.W. Heinecke, 1996. The role of oxidized lipoproteins in atherogenesis. Free Radic. Biol. Med., 20: 707-727.
CrossRefPubMedDirect Link - Brownlee, M., H. Vlassara and A. Cerami, 1985. Nonenzymatic glycosylation products on collagen covalently trap low-density lipoprotein. Diabetes, 34: 938-941.
PubMed - Brownlee, M., 1995. Advanced protein glycosylation in diabetes and aging. Annu. Rev. Med., 46: 223-234.
Direct Link - Cines, D.B., E.S. Pollak, C.A. Buck, J. Loscalzo and G.A. Zimmerman et al., 1998. Endothelial cells in physiology and in the pathophysiology of vascular disorders. Blood, 91: 3527-3561.
Direct Link - Curcio, F. and A. Ceriello, 1992. Decreased cultured endothelial cell proliferation in high glucose medium is reversed by antioxidants: New insights on the pathophysiological mechanisms of diabetic vascular complications. In vitro Cell. Dev. Biol., 28A: 787-790.
CrossRefDirect Link - Fridovich, I., 1986. Biological effects of the superoxide radical. Arch. Biochem. Biophys., 247: 1-11.
PubMed - Halliwell, B. and J.M. Gutteridge, 1984. Oxygen toxicity, oxygen radicals, transition metals and disease. Biochem. J., 219: 1-14.
PubMed - Hammes, H.P., D. Strodter, A. Weiss, R.G. Bretzel, K. Federlin and M. Brownlee, 1995. Secondary intervention with aminoguanidine retards the progression of diabetic retinopathy in the rat model. Diabetologia, 38: 656-660.
Direct Link - Hou, D.X., 2003. Potential mechanisms of cancer chemoprevention by anthocyanins. Curr. Mol. Med., 3: 149-159.
Direct Link - Hunt, J.V., M.A. Bottoms and M.J. Mitchinson, 1993. Oxidative alterations in the experimental glycation model of diabetes mellitus are due to protein glucose adduct oxidation: Some fundamental differences in proposed mechanism of glucose oxidation and oxidant production. Biochem. J., 291: 529-535.
Direct Link - Jagetia, G.C., T.K. Reddy, V.A. Venkatesha and R. Kedlaya, 2004. Influence of naringin on ferric iron induced oxidative damage in vitro. Clin. Chim. Acta, 347: 189-197.
CrossRef - Li, J.M. and A.M. Shah, 2004. Endothelial cell superoxide generation: Regulation and relevance for cardiovascular pathophysiology. Am. J. Physiol. Regul. Integr. Comp. Physiol., 287: R1014-R1030.
Direct Link - Kanner, J., E. Frankel, R. Granit, B. German and J. Kinsella, 1994. Natural antioxidants in grapes and wines. J. Agric. Food Chem., 42: 64-69.
Direct Link - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Lorenzi, M., E. Cagliero and S. Toledo, 1985. Glucose toxicity for human endothelial cells in culture. Delayed replication, disturbed cell cycle and accelerated death. Diabetes, 34: 621-627.
Direct Link - Lorenzi, M., D.F. Montisano, S. Toledo and A. Barrieux, 1986. High glucose induces DNA damage in cultured human endothelial cells. J. Clin. Invest., 77: 322-325.
PubMed - Ahmed, N., 2005. Advanced glycation endproducts-role in pathology of diabetic complications. Diabetes Res. Clin. Pract., 67: 3-21.
CrossRefDirect Link - Nishigaki, I., A. Tanimoto, S. Yasuyuki and K. Yagi, 1998. Glycated protein-iron chelate increases lipid peroxide level in cultured aortic endothelial and smooth muscle cells. Biochem. Mol. Biol. Int., 45: 519-526.
PubMed - Ross, R., 1999. Atherosclerosis: An inflammatory disease. N. Engl. J. Med., 340: 115-126.
CrossRefDirect Link - Sara, V., F. Peter and F. Hank, 2003. Therapeutic potential of breakers of advanced glycation end product-protein crosslinks. Arch. Biochem. Biophys., 419: 89-96.
Direct Link - Sensi, M., F. Pricci and G. Pugliese, 1995. Role of advanced glycation end-products (AGE) in late diabetic complications. Diabetes Res. Clin. Pract., 28: 9-17.
Direct Link - Sian, J., D.T. Dexter, A.J. Lees, S. Daniel and Y. Agid et al., 1994. Alteration in glutathione levels in Parkinson`s disease and other neurodegenerative disorders affecting basal ganglia. Ann. Neurol., 36: 348-355.
Direct Link - Szalecsky, E., J. Prechl, J. Feher and A. Somogyi, 1999. Alterations in enxymatic oxidant defense in diabetes mellitus-a rational approach. Postgrad. Med. J., 75: 13-17.
Direct Link - Tedesco, I., G.L. Russo, F. Nazzaro, M. Russo and R. Palumbo, 2001. Antioxidant effect of red wine anthocyanins in normal and catalase-inactive human erythrocytes. J. Nutr. Biochem., 12: 505-511.
CrossRefDirect Link - Thomas, O.M., L.A. Nathan, R.T. Suzanne and W.B. John, 2003. Pyridoxamine, an inhibitor of advanced glycation and lipoxidation reactions: A novel therapy for treatment of diabetic complications. Arch. Biochem. Biophys., 419: 41-49.
Direct Link - Wolff, S.P. and R.T. Dean, 1987. Glucose autoxidation and protein modification. The potential role of autoxidative glycosylation in diabetes. Biochem. J., 245: 243-250.
CrossRefPubMedDirect Link - Xu, H.Q., H.P. Hao, X. Zhang and Y. Pan, 2004. Morroniside protects cultured human umbilical vein endothelial cells from damage by high ambient glucose. Acta Pharmacol. Sinica, 25: 412-415.
Direct Link - Yagi, K., K. Ohtsuka and N. Ohishi, 1985. Lipid peroxidation caused by chinoform-ferric chelate in cultured neural retinal cells. Experientia, 41: 1561-1563.
CrossRef