
G. Kapoor
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, Shimla-171005, India
H.S. Banyal
Laboratory of Parasitology and Immunology, Department of Biosciences, Himachal Pradesh University, Shimla-171005, India
Asian Journal of Biological Sciences
Year: 2011 | Volume: 4 | Issue: 1 | Page No.: 96-100







ABSTRACT
The present study was undertaken to evaluate the humoral immune response directed against purified thioredoxin reductase (TrxR) protein during Plasmodium berghei infection. Thioredoxin reductase (TrxR; E.C. 1.8.1.9) was purified from rodent malaria parasite, Plasmodium berghei. TrxR induced humoral response in Balb/c mice immunized with parasite enzyme using saponin. Prechallenge sera gave ELISA titre of 1:512. IFA showed specificity of antimalarial antibody towards parasite. Immunization with TrxR provided partial protection in mice to P. berghei infection. Progression of P. berghei infection was delayed in TrxR immunized mice compared to normal controls.
PDF Abstract XML References Citation
How to cite this article
G. Kapoor and H.S. Banyal, 2011. Plasmodium berghei Thioredoxin Reductase Induces Humoral Response in Mice. Asian Journal of Biological Sciences, 4: 96-100.
URL: https://scialert.net/abstract/?doi=ajbs.2011.96.100
URL: https://scialert.net/abstract/?doi=ajbs.2011.96.100
INTRODUCTION
Malaria, a reemerging infectious disease is a global health hazard. The erythrocytic stages of parasite are responsible for the pathology of the disease and remain under enhanced oxidative stress due to Reactive Oxygen Species (ROS) produced by parasites metabolism and host immune system. Reactive Nitrogen Species (RNS) and ROS are accepted to kill intra-erythrocytic parasite mostly by production of INF-γ by Th-1 cells during acute Plasmodium infection, that activate the macrophages to secrete parasiticidal nitric oxide and ROS (Clark and Cowden, 2003; Stevenson and Riley, 2004). Oxidative stress is combated by the parasite with antioxidant enzymes like glutathione reductase (GR), thioredoxin reductase (TrxR) and superoxide dismutase (SOD). Plasmodium possesses a functional thioredoxin redox system comprising thioredoxin, thioredoxin reductase (TrxR) and thioredoxin dependent peroxidases representing an important line of antioxidant defense in malaria parasite (Rahlfs et al., 2002). TrxR, member of pyridine nucleotide-disulphide oxidoreductase family catalyses the transfer of electrons from NADPH to thioredoxin, which itself acts as a reductant to disulphide containing processes besides protecting cytosolic proteins from aggregation or inactivation via oxidative formation of intra-or inter-molecular disulphides. This enzyme has been found to be essential for the survival of P. falciparum erythrocytic stages (Krnajski et al., 2002) and disruption of this enzyme can be a feasible way to interfere with the development of the malaria parasite.
The present study evaluates the humoral immune response directed against TrxR protein during P. berghei infection.
MATERIALS AND METHODS
Parasite: P. berghei (NK-65) was maintained in 6-8 week old white Swiss mice, Mus musculus (Balb/c) housed in animal house of Himachal Pradesh University, Shimla under specific-pathogen free conditions by inoculating intraperitoneally (i.p.) 1x105 P. berghei-infected erythrocytes in citrate saline (2:1) from infected to naïve mice (Banyal et al., 1991). Course of infection was monitored daily by preparing Giemsa stained thin blood smears. All experiments were carried out by procedures authorized by the Institutional Animals Ethics Committee (IAEC) of H.P. University, Shimla vide letter number IAEC/Bio/19-2005.
Cell-free parasite: The citrated blood from P. berghei infected mice was collected, passed through CF-11 cellulose (Whatman) and the eluate centrifuged at 1,000 g for 10 min at 4°C. Pellet was suspended in equal volume of saponin (0.2% w/v) in phosphate buffer saline (PBS, 0.01M), pH 7.2 and incubated for 30 min at 4°C with intermittent mixing. The suspension was then centrifuged at 15,000 g for 20 min at 4°C. Haemolysate obtained was aspirated and erythrocyte membranes overlying cell-free parasite were separated and cell-free P. berghei was washed thrice with chilled 0.01 M PBS, pH 7.2.
Preparation of homogenate and purification of TrxR: Cell-free parasite was suspended in appropriate volume of 0.01 M PBS, pH 7.2 and homogenized in Potter-Elvehjem homogenizer (REMI, Bombay) at 4°C centrifuged at 1,000 g for 10 min at 4°C and the supernatant used as parasite homogenate (Kumar and Banyal, 1994). Protein was determined according to Lowry et al. (1951). The cell-free parasite homogenate was subjected to precipitation with ammonium sulphate (between 0 and 100%). The precipitates of each cut were dissolved in minimum volume of 10 mM Tris-HCl buffer, pH 7.5 and dialyzed at 4°C in same buffer containing 1 mM EDTA. The final dialyzed ammonium sulphate precipitates containing TrxR activity loaded onto pre equilibrated Sephadex G-200 (Sigma) column (1.5x20 cm) and elution of 1.0 mL volume were collected at 4°C. Each fraction was assayed for enzyme activity at 412 nm. The activity of TrxR (E.C. 1.8.1.9) was determined according to Holmgren and Bjorsnstedt (1995).
Immunization of mice: A group of 10 mice were immunized intraperitoneally with 50 μg of purified protein mixed with 30 μg of saponin in PBS (0.01 M, pH 7.2) as adjuvant. Control group of 5 mice received only the adjuvant in PBS. Two booster doses were given on day 14 and 28 postimmunization. Preimmune sera were collected before immunization. Blood samples were also collected on day 14 and 28 postimmunization and sera were analysed by enzyme-linked immunosorbent assay (ELISA).
Enzyme-Linked Immunosorbent Assay (ELISA): ELISA was carried out to evaluate the antimalarial antibody titre in the sera collected in 96-well plates according to Banyal and Inselburg (1986) using horseradish peroxidases goat antimouse immunoglobulin (Genei, Bangalore)]. Reference malaria positive and negative sera were used with each plate.
Immunofluorescence Assay (IFA): IFA was performed according to Collins and Skinner (1972) using fluorescein isothiocyanate conjugated goat anti-mouse IgG (FITC, Genei). The slides were examined under UV light and phase contrast using fluorescence microscope (Leica, Germany).
Challenge of immunized mice: The protective efficacy of TrxR was studied by parasite challenge to TrxR immunized mice with 1x105 P. berghei-infected erythrocytes on day 35-post immunization. Along with experimentals the control mice were also challenged with 1x105 P. berghei-infected erythrocytes. Course of parasitaemia was monitored by daily smear examination.
RESULTS
Thioredoxin reductase (TrxR) was purified from P. berghei by ammonium sulphate precipitation and gel filtration chromatography. SDS-PAGE analysis of P. berghei TrxR resulted in a single band of 22 kDa.
Sera of mice immunized with purified P. berghei TrxR showed presence of antibodies as determined by ELISA. Pooled pre-challenge sera of mice immunized with purified parasite TrxR gave antibody titre of 1:32 on day 14 which increased to 1:251 on day 28. On day 35 antibody titre of 1:512 was recorded before challenging the mice. No antibody titre was observed in the sera of the control mice. Post challenge serum of TrxR immunized mouse gave a high antibody titre of 1:1024. Reference malaria positive and negative serum used with experimental sera gave positive and no reaction, respectively.
Ten mice immunized with 50 μg of purified TrxR and 30 μg of saponin in PBS as adjuvant upon challenge with 1x105 P. berghei-infected erythrocytes on day 35 post-immunization showed parasites in their blood by day 4-post challenge. Three of the mice died by day 8 post challenge with maximum parasitaemia of 70%. Five mice showed migration of the parasite into reticulocytes of which 2 mice died by day 12 with 60 and 55.2% infection respectively. The other three mice died on day 14 with a maximum parasitaemia of 52.5%. Only two mice survived the parasite challenge. In these mice maximum parasitaemia observed was 3 and 7%, respectively and by day 8 the mice were cleared off the parasite. The immunized mice surviving challenge were re-challenged on day 67 post-immunization (32 days after first challenge). Upon re-challenge the mice died with parasite showing a patent period of 5 days (Fig. 1)
The placebo control mice (5) received only saponin in PBS, pH 7.2 during immunization. All mice upon challenge with parasite showed patency two days after post challenge. Three of the mice died by day 7 post challenge and showed a typical P. berghei infection pattern. The remaining 2 mice showed migration of the parasite to reticulocytes and hence, their patent period increased to 15 and 17 days respectively after which these animals also died (Fig. 1).
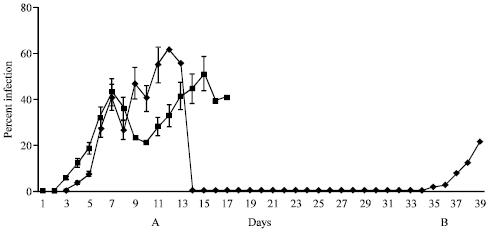 | |
| Fig. 1: | Course of parasitaemia in control mice and mice immunized with purified TrxR and challenged, (A) with P. berghei-infected erythrocytes and subsequently re-challenged, (B) with P. berghei-infected erythrocytes on day 32. Bar represents data of ten mice with SD immunized mice control mice |
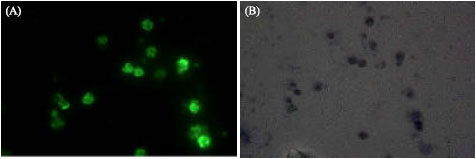 | |
| Fig. 2: | IFA reaction using serum of mice immunized with purified thioredoxin reductase. (A) As seen under UV light (x1000), (B) As seen under phase contrast (x1000) |
The specificity of the antigen-antibody reaction was analysed by IFA. Negative control using normal mouse serum did not show any fluorescence and only red counter stained reaction was seen under UV light whereas, reference positive serum showed bright fluorescence on whole of the parasite. TrxR immunized mouse sera showed specificity of parasite for antibody raised as fluorescing structure corresponded to parasite when seen under phase contrast microscope (Fig. 2A, B).
DISCUSSION
Immunity to malaria is complex and still incompletely understood. Present study was carried to evaluate the humoral immune response directed against P. berghei thioredoxin reductase using saponin as an adjuvant. Adjuvants play a major role in eliciting protective response in mice and the choice of adjuvant is important in potentiating the immune response besides determining which effector arm of immune system is stimulated. Many adjuvants like Freunds complete (FCA) and incomplete (FIA) adjuvants have been used in animals but they produce severe side effects and hence are not suitable for human use. Aluminium salts used in various vaccines are generally regarded as safe, however recent study revealed that aluminium adjuvants at levels comparable to those administered to Gulf war veterans can cause motor neuron death (Petrik et al., 2007). Saponin used as adjuvant exhibited significant humoral response and partially protected TrxR immunized animals when challenged with live parasites.
P. berghei TrxR immunized mice exhibited partial protection when challenged with 1x105 P. berghei infected erythrocytes. The sera obtained from immunized mice showed antibody titre of 1:512 in pre-challenge sera, which increased in the animals surviving the challenge. The placebo control sera did not show any demonstrable antiparasitic antibodies. A delay in the onset of infection was observed in immunized mice than the control mice and immunized mice died much later than the control mice. Perhaps the amount of antibody was not sufficient to destroy the parasite and parasitized erythrocytes in vivo following a challenge infection with P. berghei. This also indicates polyclonal B-cell activation as reported by Greenwood et al. (1970). Elevated levels of serum non-malaria specific immunoglobulins have been associated with malaria infection (Perlmann and Troye-Blomberg, 2002).
Present study shows TrxR is immunogenic and generates a humoral immune response. Other enzymes and cysteine proteases like falcipain-1 and aspartic proteases (plasmepsins) of malaria parasite have also been found to protect against infection (Kumar et al., 2007) suggesting that intracellular enzymes of Plasmodium may act as targets for antimalarial, chemotherapeutic or immunogenic. TrxR can be developed as a component of multistage, multisubunit vaccine.
ACKNOWLEDGMENT
Mr. Gaurav Kapur is thankful to ICMR New Delhi for providing JRF/SRF.
REFERENCES
- Banyal, H.S. and J. Inselburg, 1986. Plasmodium falciparum: Induction, selection and characterization of pyrimethamine-resistant mutants. Exp. Parasitol., 62: 61-70.
CrossRef - Clark, I.A. and W.B. Cowden, 2003. The pathophysiology of falciparum malaria. Pharmacol. Ther., 99: 221-260.
PubMed - Collins, W.E. and J.C. Skinner, 1972. The indirect fluorescent antibody test for malaria. Am. J. Trop. Med. Hyg., 21: 690-695.
Direct Link - Holmgren, A. and T.M. Bjorsnstedt, 1995. Thioredoxin and thioredoxin reductase. Methods Enzymol., 252: 199-208.
PubMed - Kumar, A., K. Kumar, R. Korde, S.K. Puri, P. Malhotra and V.S. Chauhan, 2007. Falcipain-1, a Plasmodium falciparum cysteine protease with vaccine potential. Infect. Immun., 75: 2026-2034.
CrossRef - Lowry, O.H., N.J. Rosebrough, A.L. Farr and R.J. Randall, 1951. Protein measurement with the folin phenol reagent. J. Biol. Chem., 193: 265-275.
CrossRefPubMedDirect Link - Perlmann, P. and M. Troye-Blomberg, 2002. Malaria and the immune system in humans. Chem. Immunol., 80: 229-242.
PubMed - Petrik, M.S., M.C. Wong, R.C. Tabata, R.F. Garry and C.A. Shaw, 2007. Aluminum adjuvant linked to gulf war illness induces motor neuron death in mice. Neuromolecular Med., 9: 83-100.
PubMed - Rahlfs, S., R.H. Schirmer and K. Becker, 2002. The thioredoxin system of Plasmodium falciparum and other parasites. Cell Mol. Life Sci., 59: 1024-1041.
PubMed - Stevenson, M.M. and E.M. Riley, 2004. Innate immunity to malaria. Nature Rev. Immunol., 4: 169-180.
CrossRef