
Charu Verma
Department of Biotechnology and Microbiology, Adhunik Institute of Education and Research, 9th Milestone, Duhai, Ghaziabad, Uttar Pradesh, India
Amit Kumar Mani Tiwari
Laboratory of Biochemistry, Department of Biotechnology, Faculty of Engineering and Technology, IFTM University, Delhi Road (NH 24), 244 102 Moradabad, Uttar Pradesh, India
LiveDNA: 91.10516
Sanjay Mishra
Laboratory of Biochemistry, Department of Biotechnology, Faculty of Engineering and Technology, IFTM University, Delhi Road (NH 24), 244 102 Moradabad, Uttar Pradesh, India
LiveDNA: 91.1019
Asian Journal of Biotechnology
Year: 2017 | Volume: 9 | Issue: 1 | Page No.: 1-23







ABSTRACT
Ripening of fruit is very important process but in some fruits, early ripening leads to a great damage during long distance transportation. There are various biochemical changes taking place during ripening such as changes in respiration, aroma, flavor, ethylene production and activity of cell wall degrading enzymes. Pectin methylesterase is one of the cell wall degrading enzymes, which acts on pectin, (a primary cell wall constituent) and releases methanol and hydrogen ions. The present compilation includes the studies on biochemical and molecular characterization of pectin methylesterase from Musa acuminata (banana). This study also deals with the in silico study reflecting inhibition of enzyme activity in context to delayed ripening in banana. It mainly deals with the identification of a PME1 gene from Grand nain variety of banana, the expression of which is related to the process of ripening. Using cross species analysis, researchers found that banana has maximum homology with carrot. The validation and verification of the designed model has been performed and found to be of good quality and further used for docking studies. Docking results suggested that green tea catechin and salicin were the best inhibitors having good interaction energies which bind at third motif, Asp381 residue on active site of PME in Musa acuminata. It was found that green tea catechin was better than salicin probably because of being a natural inhibitor, antioxidant and thus inhibiting the activity of PME. Salicin is a chemical compound that inhibits ripening at some specific concentrations only, although the binding energies of both the inhibitors have been monitored to be approximately same. The information of binding sites of ligand provides new insights into the predictable functioning of relevant protein.
PDF Abstract XML References Citation
Received: September 20, 2016;
Accepted: November 07, 2016;
Published: December 15, 2016
How to cite this article
Charu Verma, Amit Kumar Mani Tiwari and Sanjay Mishra, 2017. Biochemical and Molecular Characterization of Cell Wall Degrading Enzyme, Pectin Methylesterase Versus Banana Ripening: An Overview. Asian Journal of Biotechnology, 9: 1-23.
DOI: 10.3923/ajbkr.2017.1.23
URL: https://scialert.net/abstract/?doi=ajbkr.2017.1.23
DOI: 10.3923/ajbkr.2017.1.23
URL: https://scialert.net/abstract/?doi=ajbkr.2017.1.23
INTRODUCTION
Fruits are important nutritional food for balanced diet in humans and play pivotal role for providing various regulating factors, which are essential for health. Among fruits, banana is one of the most important staple foods for people to supply the nutritional value.
Banana plant is an herbaceous, monocotyledon and perennial, belongings to Musaceae family1. The genome of banana is commonly triploid having AAA type e.g., Cavendish and having property of parthenocarpic2. Bananas are of two types, which are available throughout the year, namely the dessert banana and plantains3. The origin of dessert banana is Indo-Malaysia, whereas plantain are originated in East, West and Central Africa4-6. The genomes of different varieties vary from diploid to triploid6. Dessert bananas belong to AAA group like Gros Michel and Cavendish1,7, while plantain belongs to AAB group. Due to the susceptibility of Panama disease, Gros Michael variety having been the most popular earlier was replaced by Cavendish group4,8,9.
On the basis of banana production, the top producing countries are India, Brazil, Ecuador, China and Philippines. Bananas are rich sources of carbohydrates, vitamin C, vitamin B6 and potassium1. The ripe pulp and peel of banana fruit contain norepinephrine, dopamine and serotonin2. Dopamine and norepinephrine help in rising blood pressure, while gastric secretion is decreased by serotonin. Banana plants can also be used as an ornamental plant and for fibre6. There are certain other uses of different parts of banana such as food wrapping in large leaves of banana plant and stem for the purpose of flooring in some countries8.
During the ripening, fruit can be classified as climacteric or non-climacteric on the basis of their respiratory property. The rate of respiration and ethylene synthesis increases in climacteric fruits such as apple, banana and tomato. In contrast, there is not a sudden increase in respiration and also in ethylene synthesis in non-climacteric fruits such as citrus, cherry and strawberry.
PLANT CELL WALL AND ITS COMPOSITION
Cell wall is the outermost covering of a plant cell. It is the most important and distinguishing characteristic of plant cell. Animal cell lacks the cell wall and hence these cells are more flexible required for movement and locomotion. The plant cell wall plays various vital roles such as (i) Protection of all intracellular components, (ii) Providing rigidity and texture to the plant cell, containing a medium for transport and circulation of water, minerals and other nutrients and (iii) Resistance to the entry of pathogens to protect the plant from various infections and diseases. The thickness and composition of the cell wall may vary significantly.
The cell wall consists of three types of layers, namely middle lamella, primary cell wall and secondary cell wall. Middle lamella is the first layer formed during cell division. It is the outermost wall of the cell and establishes connections to the adjacent cells. The composition of this layer is mainly the pectic components and proteins. After the formation of middle lamella, there is a synthesis of primary cell wall, which consists of rigid cellulose microfibrils present in the matrix of pectic compounds, hemicellulose and glycoproteins. After cell enlargement, secondary cell wall is formed, which is the most rigid and thus providing strength. It is composed of cellulose, hemicellulose and lignin.
The principle components of the plant cell wall are the polysaccharides. These polysaccharides are made up by the polymerization of sugar molecules by glycosidic linkages. In plant cell, various polysaccharides chains are integrated to each other and form a network. Plant cell wall consists of mainly three types of polysaccharides, namely, cellulose, hemicellulose and pectic substances. Besides, some structural proteins are also present.
Cellulose: Cellulose is crystalline, formed by the aggregation of fibers by β-1, 4 linked glucan chains2. These cellulose fibers are responsible for providing the extensive strength. In cellulose, the β-D-glucose are linked by glycosidic linkages of C-1 of one residue to the C-4 of second adjacent residue. It consists of nearly 1,000-10,000 β-D-glucose residues. It is the major component of primary and secondary cell wall. In cellulose, hydrogen bonds are responsible for micro fiber formation and millions of micro fibers are interconnected to form fibers containing millions of hydrogen bonds having very high bonding energy that provides the basis of high tensile strength of cellulose3.
Hemicellulose: Hemicellulose is mainly present in primary cell wall. It is also observed in secondary cell wall. It is a polysaccharide, which constitutes a large number of sugars like xylose, arabinose and mannose. Hemicellulose along with xylose sugar called as xyloglucans whereas with arabinose known as arabinoglucans. The molecules of hemicelluloses are hydrophilic in nature. The backbone of all hemicelluloses contains straight β-1, 4 linkages. The chain of hemicelluloses does not extend self-aggregation like in cellulose due to some structural features and thus is not available for hydrogen inter-chain bonding. The chain of hemicelluloses cannot self-aggregate to form fibers like in cellulose but sometime can form bonded with each other10,11.
Structural proteins: The primary cell wall contains various proteins. Glycoproteins are known to contain carbohydrates side chain. In the composition of cell wall protein, hydroxyproline amino acid is present abundantly. Glycoproteins are water soluble and can form hydrogen bonds with cell wall polysaccharides10. Proline and lysine are also high in cell wall protein. The peptide chain contains positive charge due to the presence of ![]() of lysine residue, which binds with the negative charged groups on pectic acid. The other example of structural cell wall protein is extensin, which can form covalent interactions with other extension proteins by tyrosine amino acid residue. The presence of thick and hard cell wall is also due to the rich content of extensin and the concentration of this protein changes with development11.
of lysine residue, which binds with the negative charged groups on pectic acid. The other example of structural cell wall protein is extensin, which can form covalent interactions with other extension proteins by tyrosine amino acid residue. The presence of thick and hard cell wall is also due to the rich content of extensin and the concentration of this protein changes with development11.
Pectic substances: A group of closely related polysaccharides present in the plant cell wall is called as pectic substances or pectin. Pectin is mainly responsible for the rigidity of tissue. It is a complex macromolecule composed of about 17 different monosaccharides with different linkages12,13. It contributes a major role in cell wall formation14. Pectic substances are made up about one-third of dry substances of primary cell wall of fruits and vegetables and a large proportion of middle lamella15. In middle lamella pectic concentration is the highest and then gradual decreases from primary cell wall to secondary and then plasma membrane16.
The mechanical strength of the cell wall and adhesion between the cells are provided by the pectic substances17. Several properties of cell wall like porosity, surface charge, pH and ion balance are influenced by pectin and so important in ion transport in the cell wall18. They are also known to participate in activating the defense mechanism as they remove the accumulation of phytoalexin having anti-microbial activity19. It also helps in the induction of lignifications20 and accumulation of protease inhibitors in plants. In food products such as jam, yoghurt, drinks, fruit milk drinks and ice-cream, pectin is frequently used as a thickening, gelling and stabilizing agent21. A large proportion of pectin used in food industry is obtained from the source of citrus or apple peel isolated at high temperature and low pH and is a hamogalacturonan22. It has been studied that the soup of carrot contains pectin derived oligosaccharides, which prevents various pathogenic microorganisms to get attached to intestinal mucosa in vitro and it is an important step in pathogenesis of gastrointestinal infection23,24. Pectins have immune regulatory effects in intestine to alter the activity of microbes and thus changing the morphology of the wall of small intestine25,26 to decrease the level of blood cholesterol27 and the glucose absorption in the serum of diabetic and obese patients28.
Water insoluble pectic substances are known as protopectin. A chain of polygalacturonic acid with low content of methoxyl group called as pectic acid whereas with high methoxyl content is called as pectinic acid. Pectin is a water soluble pectinic acid of different methyl ester group. Sometimes pectin is used in place of pectic substances. The predominant component of pectin is D-galacturonic acid (GalA) residue linked by α (1-4) linkages, on which varying proportion of acid groups are present as methoxyl ester28. The homogalacturonan regions are called as "Smooth region" whereas the highly branched part is referred to as ‘hairy’ or ramified region, in which many neutral sugars are present. There are various pectin structural elements containing the same repeating unit but the structure and number of these units varies29-31. Some pectin structural elements are discussed in this study.
Homogalacturonan (HG): The main type of pectin in the cell wall is homogalacturonan constituting about 60% of the total pectin32,33. The backbone of HG is made up of a polymer of α-1,4 linked GalA residue34. In this backbone, the GalA may be mthyl esterified35 at C-6 and O-acetylated at O-2 or O-336. Methyl esterification of pectin is related to changes its structure. The molecules of HG are interspersed with L-rhamnose residue, which results in a kink37.
Xylogalacturonan: Xylogalacturonan results by the substitution of HG with β-D-Xylp-(1->3) single unit side chain37-39. In XGA, a part of GalA residue is methyl esterified and among the substituted and un-substituted GalA residue, these methylesters are distributed equally38,40. The XGA is mainly found in reproductive tissues like fruits and seeds37,38. Recently, it has been documented that XGA is present in many tissues of Arabidopsis thaliana41.
Rhamnogalacturonan I (RGI): The backbone of RGI consists of (->2)-α-L-rhap-(1->4)-α-D-GalpA(1->)18. The pectin oligosaccharides molecules in sugar beet contain a chain of length upto 20 residues alternating Rha and GalA units. In RGI, the rhamnosyl residue molecule can be substituted at O-4 with the side chain of neutral sugar42. The side chain is composed of galactosyl or arabinosyl residue. The GalA residue of RGI are presumably not methyl esterified because RGI is not degraded under β eliminative condition43, but a fraction of RGI has been reported to contain 40% methyl esters44. In RGI, the GalA residues may be highly O-acetylated on O-2 or O-3 position45,46. Highly branched region of pectin are known as ‘hairy’ or ‘ramified’ region.
Rhamnogalacturonan II (RGII): The RGII is likely to be released by the action of endopolygalacturonase (PG) and is much conserved structure in plant kingdom. The structure of RGII is characterized by a distinct region within HG, which contains cluster of four different side chains with peculiar sugar residues. These side chains are attached to HG fragment of about 9 GalA residue and some of them are methyl esterified12,32,47.
Arabinan and arabigalacturonan: The backbone of arabinan composed of 1,5 linked α-L-Araf-(1->2)-α-L-Araf-(1->3) or α-L-Araf-(1->3)-α-L-Araf-(1->3) side chain12. Arabinogalacturonan is of two types, namely AGI and AGII. The AGI is composed of a 1,4 linked β-D-Galp backbone with α-L-Araf residues attached to O-3 of the galactosyl residue12,45. The chain of AGI can be terminated with α-L-Arap-(1->4) at the non-reducing end48.
The chain AGII consists of 1,3 linked β-D-Galp backbone alongwith the side chain of α-L-Araf-(1->6)-(β-D-Galp-(1->6)n, where n = 1,2 or 3 (1, 3, 54). The AGII is also associated with proteins of about 3-8% called as arabinogalactans proteins (AGPs). The protein region is mainly consists of proline, hydroxyproline, alanine, serine and threonine49. Pectin andAGII are fractionated partially as it is difficult to separate those50. It has been reported that a small fraction of carrot tap root cell wall AGPs is linked to pectin51.
CELL WALL DISAGGREGATION AND PROCESSING OF FRUIT SOFTENING
The amount of pectins is higher than proteins in fruit cell wall. Pectins are major constituents of primary cell wall and middle lamella52. A network of pectins is present in between the spaces of cellulose and hemicelluloses cross linkages52. Pectins are group of heteropolysaccharides containing galacturonic acid. The structure of galacturonic acid may be linear known as homogalacturonan or branched called as rhamnogalacturonans.
The cell wall disintegration takes place due to the solubilization of pectins and hemicelluloses. Diverse groups of cell wall modifying proteins are involved in dismantling of polysaccharides. This class of proteins is localized in cell wall called as expansins53. The role of expansion proteins has been monitored in disrupting the hydrogen bonds between cellulose and xyloglucans54. Thus, in fruit softening, expansion proteins also play a key role55.
Fruits are very important for providing various essential nutrients for normal human health. Fruit softening is one of the most significant indicators of ripening. Fruit softening is a developmental program during ripening, which results in changing the texture56. The process of fruit softening is generally due to the effects of various modifications in cell wall, which results in the loss of rigidity, ultimately affecting their texture. During ripening, various biochemical events take place such as (i) Chlorophyll degradation, leading to yellowing of the peel, (ii) Starch conversion into sugars and (iii) Change in flavor and aroma. Ripening of fruit is also regulated by the production of ethylene. The quality of fruits and vegetables are also maintained by calcium57. It plays an important role in firming the fruit by β-elimination. Tissue softening is also due to the decrease in intermolecular bonding between polymers of the cell wall. The modification of fruit cell wall is greatly achieved b y the activity of hydrolytic or cell wall degrading enzymes such as polygalacturonase (PG), pectin methylesterase (PME), Pectate Lyase (PL), rhamnogalacturonase (RGase).
Softening of fruit involves generally cell wall loosening accompanied by depolymerization of hemicelluloses and action of polygalacturonase and other cell wall degrading enzymes52.
CELL WALL DEGRADING ENZYMES
During ripening, softening of fruit is related to changes in the composition of cell wall due to various factors, mainly, pectin degradation by hydrolytic enzyme. There are some cell wall degrading enzymes such as PG, PME, PL, RGase, arabinase and galactanase, involved in pectin solubilization. The principle hydrolytic enzymes are PME and PG.
The PME is mainly involved in de-esterification by removing methanol from C-6 position of galacturonic acid of pectins. The activity of PME was observed in fruits such as apple, banana, grape, tomato, strawberry58. The PME removes methyl esters, which are then depolymerized by PG and results in reduction of tissue rigidity and adhesiveness59. Fruit firmness is increased by the action of PME due to the demethylation of endogenous pectins and subsequent chelation of divalent cations by ionized carboxyl groups on adjacent pectic acids60. The PME is found in both plants and micro-organisms61. The PME occurs in both exogenous and endogenous forms. In fungi, PME acts on homogalactuonans randomly whereas linearly in plants62. The degree of methyl esterification of pectin and pH are responsible for PME activity. There are many isoforms of PME encoded by multigene family63,64. At acidic pH, some isoforms can act randomly but at alkaline pH, act linearly. PG is mainly responsible for solubilization of pectins. It causes depolymerization by hydrolysing α-1,4 glycosidic bonds present in galacturonic acid. The activity of PG is increased during ripening by the presence of soluble pectins65. The PME is responsible for de-esterification followed by action of PG on de-esterified portion of galacturonan. In climacteric fruits, an increase in PG activity during ripening is corelated with the maximum loss of rigidity or firmness66. For the study of textural regulation in fruit ripening, PG was the first to be cloned from tomato by transforming with PG antisense, which results in increase in texture quality, ultimately their shelf-life increasing67.
Pectate Lyase (PL) is also a hydrolytic enzyme, which plays an important role in breaking the glycosidic bond by trans β-elimination. Rhamnogalacturonase (RGase) catalyzes the hydrolysis of glycosidic bond between galacturonic acid and rhamnose in RG chain46. Cellulases as well as hemicellulases are also involved in fruit softening.
BIOCHEMICAL CHANGES DURING RIPENING
The key feature in fruit deterioration is the rate of fruit softening. The shelf life of fruits becomes shorter on excessive softening and leads to a huge damage during transportation and storage. In tomato fruit, a large number of genes have been reported to be involved in degradation of cell wall. The suppression genes encoding the cell wall degrading enzymes such as PG, PME, PL and β-glucanase do not have a major effect on firmness68,69. About 40% of tomato fruit softening is achieved by down-regulating TBG4β-galactosidase gene70. Cell wall proteins known as expansins disrupting hydrogen bonds between cellulose microfibrils and polysaccharides lead to cell wall loosening. A protein in tomato fruit is expressed in fruit ripening is encoded by LeExp1 (tomato expansin1) gene. There is a strong reduction of softening throughout ripening due to the down-regulation by changing glycan interface, facilitating the access of cell wall hydrolases71. Strawberry, which is a non-climacteric fruit, becomes firmer and reduces softening by suppression of pectatelyase m-RNA72. Many genes are involved in fruit softening, encoding various cell wall degrading enzymes and non-enzymatic proteins. Each of proteins and their isoforms play a significant role in textural changes and fruit softening.
There are several factors involved in the attractiveness of fruit. One of the important examples of such factor is pigments, which accumulate in the skin during the process of ripening, in some climacteric fruits73. Pigments are known to accumulate in the pulp tissue too. Carotenoids and anthocyanins are the most important pigments of fruit because they also involve as source of vitamin A and antioxidant compounds for human health. Carotenoids consist of carotenes like lycopene and β-carotene and xanthophylls like lutein. They are obtained from terpenoids and synthesized at a high rate in the fruit during transition from chloroplast to chromoplast. Several genes have been cloned and are involved in the synthesis of carotenoids74. On the other hand, anthocyanins belong to the flavonoid subclass of phenolic compounds.
Many genes and their enzymes of flavonoid biosynthetic pathway have been isolated and characterized75. Anthocyanins are important for wine quality in grapes. Ethylene stimulates the colouration of berry and is involved in the regulation of anthocyanin biosynthesis genes76. There are various factors and signals, which affect the anthocyanin accumulation and gene expression, including photochrome and light. Many stresses such as low temperature and wounding are also involved77. The quality of fruits and vegetables are strongly affected by aroma volatiles. Aroma is a complex mixture of large number of compounds. Each product has a different aroma. Most important classes of aroma are monoterpenes, sesquiterpenes and compounds derived from lipids, sugars and amino acids. The rate of ripening and duration of storage life and other ripening processes in climacteric fruit is controlled by the ethylene. Therefore, various efforts have been made to increase the shelf life of fruits by generating the genotype of incidently reduced ethylene synthesis, because ethylene regulates various genes of aroma biosynthesis78.
Ripening involves the transition from an inedible state of fruit to the prerequisite characteristics of food quality79. It involves a series of various physiological changes in mature plant organ. When an appropriate level of ethylene is reached in mature cells, initiation of ripening takes place80. During the ripening of fruit, there are various biochemical changes to occur81 and are as follows.
Cell membrane changes: There are a number of vital roles of membranes such as energy transfer, hormone binding, signal transduction, interaction between plant and pathogens and transport of ions and other solute molecules. Membranes are thin, flexible and sheet like structures enclosing all the cell organelle82. Membranes are made up of lipid bilayer in which the integral and peripheral proteins are embedded83. Phospholipids can freely rotate in this fluid mosaic of membrane and the fluidity is influenced by the presence of sterols and proteins. Fluidity of membrane also enhances by increasing the content of unsaturated fatty acids.
During the ripening of banana fruit, the content of unsaturated fatty acids decreases in the pulp and peel, which results in increase in microviscosity82,83. During senescence, there are some visible changes in lipid bilayer such as degradation of lipids, which leads to impaired functioning. Bilayer may exist in different phases. It can change from a liquid to a solid or gel phase. On ripening of fruit the permeability of the membrane is changed and thus responsible for the leakage of solutes leading to an increase in free space. It has been reported that there is an increase in membrane permeability of banana pulp as a post climacteric respiration event84. As banana fruit ripens, the leakage of ions becomes increasing. There are some harmful changes in membrane during the senescence such as the content of phospholipids and proteins decreases and an increase in saturated: unsaturated fatty acid ratio. An increase in short chain fatty acids like octanoic, nonanoic and decanoic acid has been observed to correlate to an increase in ethylene sensitivity in banana ripening fruit and petals of petunia84.
Respiratory changes: In plants, the process of respiration includes the breakdown of complex molecules like starch, sugar and acids into simpler molecules CO2 and H2O85. Certain pathways are known to involve in the complete process of respiration are glycolysis, citric acid cycle or TCA cycle, pentose phosphate pathway and an Electron Transport Chain (ETC)83.
Glycolytic pathway is the first step in process of respiration and occurs in the cytosol of cell. In glycolytic pathway, glucose molecule is broken down into two molecules of pyruvate as shown in Fig. 1. The key enzyme in this pathway is phosphofructokinase (PFK) that breaks fructose-1,6-bisphosphate into two molecules of trioses85. During glycolysis, the net gain is of two molecules of ATP and two molecules of NADH. The overall glycolytic pathway can be summarized as follows:
Glucose+2ADP+2Pi+2NAD+ → 2 pyruvate+2ATP+2NADH+2H++2H2O
Additionally, TCA cycle also known as citric acid cycle, occurring in mitochondria of the cell has been known to be connected with respiratory phenomenon while fruit ripening and softening. In this cycle, the pyruvate from glycolytic pathway is broken down to CO2 in various steps.
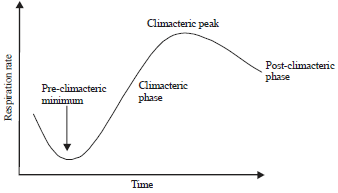 | |
| Fig. 1: | Graph of climacteric pattern of respiration on ripening of fruit |
This pathway involves the generation of one ATP molecule. Three molecules of NAD are reduced to NADH and one molecule of FAD to FADH283,86.
Electron transport chain: This metabolic chain takes place in the cristae of mitochondria. It involves the transfer of hydrogen atoms from organic acids in TCA cycle and production of ATP from NADH2 and FADH2. Each NADH2 molecule gives 3 ATP molecules whereas each FADH2 molecule produces only two ATP molecules. The function of ETC is known to trap energy in a usable form and also recycle NAD and FAD, required for the functioning of various pathways. The binding of cyanide to the iron atom of cytochrome oxidase inhibits the electron transport system86. In some plants such as banana and avocado, cyanide resistant respiration can occur by the action of an alternative oxidase. In this pathway, the electron transport from NADH to ubiquinone followed by their transfer to a flavoprotein and then to the alternate oxidase and then ultimately to oxygen, to form water. Only one molecule of ATP is generated through this process instead of three. Energy is not conserved and mostly releases in the form of heat. A higher rate of respiration is related to more perishable85. An increase in the temperature of commodity due to the heat released by respiration causes an increase in metabolism rate and decrease in longevity.
Respiration in climacteric fruits: A climacteric pattern of respiration is seen in some fruits during the process of ripening83. There are four phases of climacteric pattern of respiration i.e., pre-climacteric, climacteric, climacteric peak and post-climacteric87 (Fig. 1). This respiration profile of climacteric fruit is related with the release of high level of CO2 and production of ethylene during ripening. The climacteric pattern of respiration is exhibited by banana in four phases.
In pre-climacteric phase, the color of fruit is green, hard in texture, inedible and contains a high content of starch and the rate of respiration is low. During climacteric phase, the flavor of fruit starts to develop by conversion of starch into sugars. The texture starts to become soft and less chlorophyll is visible. The rate of respiration becomes maximum at climacteric peak and at this stage, the fruit is right for consumption. At the post climacteric phase, there is a decrease in the rate of respiration and the fruit becomes overripe. At this stage the fruit is tasteless and the pulp turns to mealy.
Color changes: The color changes are first visual indication of ripening and thus determine the eating quality88. Banana ripening is also related with the color changes i.e., from green to yellow and then turns to brown on over ripening. The color changes in banana starts in the center of fruit and then proceeds towards the edge88. Changes in fruit color are mainly due to the presence of pigments located in chloroplast, chromoplast and vacuoles of the cell88. Anthocyanin pigments are water soluble and present in vacuoles whereas the carotenoids pigments are water insoluble and located in chromoplas88. The peel color of banana changes due to the presence of several pigments such as chlorophylls, carotenes and xanthophylls located in chloroplast and chromoplast.
On ripening of fruit, the color changes are related with the chlorophyll breakdown and the unmasking of carotenoid pigments83. On rise of climacteric phase, the activity of chlorophyllase increases to breakdown of chlorophyll. In addition, there are some other factors playing a vital role in changing the color of fruit such as light, temperature, oxygen and time of harvest. It is not mandatory that color change is always an indication of ripening of fruit e.g., in apples83.
Changes in organic acids during ripening: The flavor of the fruit is also associated with organic acids. On fruit ripening, concentration of some specific organic acids decreases and thus leads to decrease in acidity83,86. Malic acid, oxalic and citric acid are mainly present in banana ripening. The main component of organic acid in fruits like banana, apple, cherry, peach and pears is malic acid83,86. On banana ripening the level of malic acid increases, while there is a metabolism of oxalic acid and due to their role in TCA respiratory cycle as the substrate, there is a loss of acidity83,86.
Carbohydrate changes: Starch is the main storage form of carbohydrates in green banana comprising about 20-25% of weight of the tissue pulp89. When the fruit starts to ripen, the starch is hydrolyzed and converted into free sugars, with remaining only 1-2% starch in the ripe fruits. The content of ripen banana consists of 60% sucrose, 14% fructose and 20% glucose. Due to the presence of these sugar molecules, edibility of fruit increases and thus is responsible for the sweet flavor of fruit. When the starch is degraded into sugars, the activity of starch biosynthesis enzymes decreases whereas the enzymes involved in starch hydrolysis increase89. The conversion of starch into sugar is catalysed by the activity of enzymes, namely, amylases and starch phosphorylase. In banana, the breakdown of starch is catalysed by starch phosphorylase rather than amylase89.
Production of aroma components during ripening: At the time of ripening of fruit, their characteristic aroma is produced by the complex mixture of volatile compounds90. A certain number of volatile compounds are produced by each fruit such as at least 200-350 are produced by a ripe banana90. After duration of 4 days of climacteric respiration, volatiles are produced. Aroma occurring during the course of fruit ripening is due to the presence of various classes of volatiles such as aldehyde, ketones, esters, terpenoids and sulphur containing compounds90. Stimulation of synthesis of these compounds takes place on exposure of banana fruit to exogenous ethylene. As fruit ripens, there is a gradual increase in the complexity and concentration of volatile compounds91. In banana fruit, main volatile compounds are esters, alcohols, phenols, aldehyde and ketones. These volatile compounds are obtained from amino acid and fatty acid (Fig. 2). About 70% of esters are present in volatile compounds, in which the predominating fractions are of acetates and butyrates. The production of aroma compounds is decreased by providing the chilling injury91.
Ethylene: A Russian plant physiologist Dimitry Neljubov discovered the effect of ethylene on plant tissue92. The chemical formula of ethylene is C2H4 with molecular weight83 28.5. Under normal conditions, it is present in the gaseous state and helps in the regulation of plant growth, development and senescence93. Ethylene is commercially available in compounds such as ethephon and ethrel, which is used to ripen fruit like bananas, tomatoes, apples and pear93. There are some sources of ethylene present in atmosphere such as from combustion engines or heaters in the form of fumes, smoke, natural gas leaks93. Ethylene is also produced naturally in all plant organs such as roots, stem, leaves, buds, tubers, bulbs and flowers94. Production of ethylene can also be stimulated as a result of biotic and abiotic stresses. Ethylene is responsible for ripening and also plays an important role in the activities of adventitious root formation, flower induction, fruit abscission, flower opening and senescence of leaf and flower94.
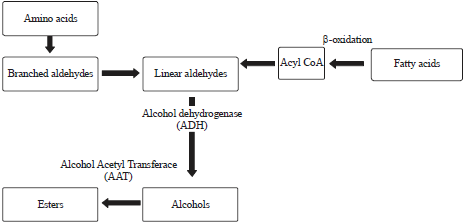 | |
| Fig. 2: | Pathway illustrating the production of aroma compounds |
At all developmental stages of fruit, the ethylene is present at low concentration but during the ripening of climacteric fruit, the activity of ethylene becomes more dominant. The release of ethylene into the plant tissue is responsible for a change in fruit quality such as color, texture and flavor95. For production of ethylene in plants, there are two systems, namely system 1 and system 2. System 1 is present throughout the development in both climacteric as well as non-climacteric fruit whereas system 2 is activated during ripening in climacteric fruit. Ethylene possesses the autocatalysis property and thus can stimulate its own synthesis. Through autocatalysis, the tissue can synthesize large amount of ethylene in response to application of low concentration of this gaseous hydrocarbon. Ethylene present in the atmosphere or in plant tissue is responsible for positive feedback and thus results in increased production of this hormone96.
First ethylene responsive genes isolated from tomato, include genes encoding cell wall degradation, pigment synthesis and ethylene production. On the basis of expression studies it reveals that ethylene responsive genes can be up-regulated, down-regulated or transiently induced following hormonal treatment for short duration encourages the view that ethylene can act as positive or negative regulator of gene expression97,98. Many of the regulatory proteins encoded by early ethylene responsive genes involved in various processes such as transduction pathways, regulation of transcriptional or post-transcriptional factors, shows that ethylene control operates in a complex multilevel. It has been documented that ethylene have several effects on different developmental processes such as germination, abiotic stress response, fruit ripening, abscission of leaves, root nodulation, programmed cell death and pathogen attack99,100.
To understand various roles of ethylene responses to plant, there is a signal transduction pathway. The first transcription regulator is an EIN3. So, it can be concluded that the diversity of ethylene responses may arise from the expression of Ethylene Response Factors (ERFs), transcription regulator proteins present at the downstream of EIN3. The ERFs are involved in the largest families of transcription factors in plants101 and thus are available for different channels of hormone signaling for a variety of responses102. A conserved motif of cis-acting element called as GCC box is present in the promoter of ethylene responsive genes, upon which trans-acting factors encoded by ERF genes are bound. The ERFs belong to a large multigene family, which contains diverse binding activities and multifunctionality.
Fruit ripening processes involve complex multi-hormonal control and are genetically regulated processes. The effect of phytohormones on plant development consists of a chain of signal transduction pathways, known to activate specific transcription factors and ultimately regulate the expression of target genes, shown in Fig. 3.
CELL WALL HYDROLASES
On the ripening of fruit, firmness decreases and osmotic properties of the cell also vary. The loss of firmness is also resulted due changes in cell wall polysaccharides103. There are some chemical changes in the cell wall which focus mainly on changes in pectic substances103. Pectins are present in the form of polysaccharides, which are easily soluble in water and hence get de-esterified and depolymerized by certain enzymatic reactions. The addition of Ca2+ ions is related to decrease of textural softening due to having an ability to form calcium bridges between pectic polysaccharides chain103. The reduction in fruit firmness is due to acid hydrolysis of glycosidic bonds in the cell wall.
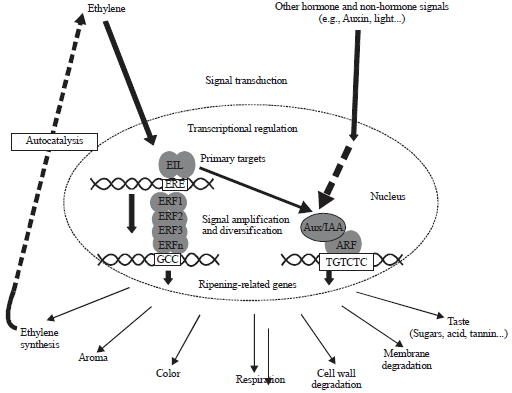 | |
| Fig. 3: | Hormonal dependent transcription regulation of fruit ripening104 |
The most stable linkage in between galacturonan in pectin is glycosidic103. The hydrolysis of galacturonans is faster in acidic pH as compared to neutral sugars because the uronic acid residues are lost whereas neutral sugars are not and they are bound with the pectin103.
Thus, in acidic conditions, the fruit softening during ripening is due to hydrolysis of pectin105. There is a gradual solubilization of protopectin to form pectin, which leads to loss of firmness on ripening of fruit.
During the process of ripening of fruit after pectin solubilization, depolymerization and de-esterification take place. By the action of carbohydrate hydrolyzing enzymes, the composition of cell wall changes, thus resulting in fruit softening. Many of these hydrolytic enzymes are present in low level and are constitutive throughout the fruit development105, but mostly the activity of these enzymes are increased on fruit ripening. Among all hydrolytic enzymes, fruit softening is mainly due to pectin degrading enzymes and results in loss of firmness. An increase in the activity of polygalacturonase relates with the ripening of fruit105. After the activation of polygalacturonase (PG), pectin polymers are solubilised, because this enzyme helps in the breakdown of insoluble complex polysaccharides by reducing the chain length interconnection by calcium106.
During ripening, the changes in pectin occur in many fruits like strawberry107, kiwifruit108, apricot109 and olive fruit110. There is a positive relationship between the presence of polygalacturonase and fruit softening observed in many fruits like papaya110 and mango111. Activities of PG and PME have been reported to increase in peach, tomato and pear65. Process of solubilisation of polyuronide is different in apple and strawberry due to the absence of endo-PG28. The activity of PG was not detected in plum fruit112. Although PG is important for pectin degradation but it is not only the factor for tissue softening during ripening113. The hydrolysis of neutral sugar side chains helps in weakening the network of polymerization of cell wall and contributes to the textural softening114. Antisense RNA technology is a new approach to elucidate the role of enzymes in cell wall degradation responsible for fruit softening. This was the first molecular approach concerning with delaying of fruit ripening115. Firmness and shelf life of tomatoes can be increased by suppression of PG gene with antisense RNA technology. The suppression of PME in tomatoes results in increase in solid content68. The genes coding for PG, PME and other enzymes have been cloned in tomato113 and in other fruits115. Pectin degrading enzymes are of various types on the basis of their mode of action. Some cell wall hydrolytic enzymes are pectatelyase, polygalacturonase, pectin methylesterase, rhamnogalacturonase and arabinase.
Polygalacturonase (PG): The main enzyme responsible for pectin dissolution in vivo is polygalacturonase enzyme116. It plays an important role in hydrolysis of pectic acid or galacturonic acid by hydrolyzing α-1,4-glycosidic bonds between them. The PGs are divided into two forms on the basis of their mode of action i.e., exo-PG and an endo-PG. Exo-PG is responsible for the hydrolysis of glycosidic bonds between the de-esterified galacturonans from non-reducing end, resulting in release of galacturonic acid as main product116. Degree of polymerization depicts the rate of hydrolysis and with increase in molecular size of substrate, it increases116. Exo-PG helps in producing large reducing groups. Pectate degradation does not occur by the activity of this enzyme. So, it is not involved in fruit ripening. Endo-PG plays an important role in ripening as it causes the depolymerization of pectic acid randomly. As the length of the chain decreases, the rate of hydrolysis also decreases.
During ripening of fruit, the rate and extent of textural softening is directly related to the composition of PG i.e., if an -endo or both -endo and -exo PG are present, a significant softening takes place and if only -exo PG is present, restricted softening occurs117. The PG is responsible for the dissolution of middle lamella. On ripening, presence of soluble pectin results by increased activity of PG28. The PG acts on glycosidic bonds of de-esterified galacturonan chain. The PG was first observed in ripen tomato and it is the richest source of polygalacturonase enzyme106. It has been reported that at the climacteric stage, the activity of PG is increased in mango118, capsicum116 and banana119. At the time of ripening, the texture of climacteric fruit changes and a rapid increase in polygalacturonase is directly related to the maximum loss of firmness66. The purification and characterization of PG as well as biochemical specificity of isoenzymes of PG were reported from banana120,121 and strawberry107.
Pectatelyase (PL): The PL enzyme, also known as trans-eliminase, breaks the glycosidic bond by trans β-elimination. It removes hydrogen from C4 and C5 position of the aglycone of substrate122. Pectin undergoes de-esterification in alkaline medium and then degrades by β-elimination reaction. PL catalyses the cleavage of deesterified or esterified galacturonate units by a trans β-elimination of hydrogen from the C4 and C5 positions of galacturonic acid. The PL also exists in two forms i.e., exo-PL lyase acting from the non-reducing end, whereas endo-PL lyase acts randomly on de-esterified galacturonans.
Arabinase: This enzyme is of two types, namely, arabinofuranosidase (EC 3.2.1.55) and endo-arabinase (EC 3.2.2.99). It increases the polymer-polymer interaction and decreases the degree of branching122. Endo-arabinase acts on linear arabinan randomly resulting in the production of short length oligomers. On the other hand, arabinofuranosidase is responsible for degrading branched chain into linear chain of arabinan by splitting of terminal α-1,3 linked arabinofuranosyl side chains and then breaks α-1,5 links at the non-reducing end of linear arabinan. This enzyme hydrolyses the non-reducing group from a large number of polysaccharides containing arabinose such as arabinogalactans, arabinoxylans and arabinans.
Rhamnogalacturonase (RGase): This enzyme catalysed the hydrolysis of glycosidic bonds between galacturonic acid and rhamnose units in RG backbone, the "Hairy regions" of many fruit pectins31. The products are the alternating units of galacturonic acid and rhamnose units. It forms the non-reducing end31. The activity of RGase increases when the side chains are removed and the esters group are de-esterified31. The activity of RGase is interrupted by o-acetyl group. So, they perform with rhamnogalacturonan acetylesterase, which removes the acetyl group from hairy portion of pectin28. The activity of RGase is more on de-esterified pectin31. Thus, for the intense activity of glycanase such as PG and RGase, de-esterification of pectin is the most important step which is catalysed by the enzyme pectin methylesterase (PME).
Pectin methylesterase (PME): The PME (EC 3.1.1.11) is a pectinase enzyme, which acts on pectin substrate. On the basis of mode of action on pectin substrate, pectinases classified into two groups are deesterifying and depolymerizing enzymes123,124. The PME catalyzes the demethyl esterification of pectin and liberated carboxyl groups and methanol (Fig. 4).
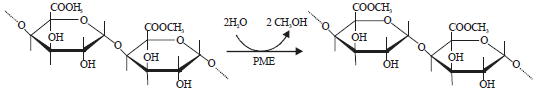 | |
| Fig. 4: | PME causes de-esterification of pectin125 |
The PME removes methanol from highly esterified pectins and convert them into low ester pectin, which becomes later hydrolysed by PG, attacking glycosidic linkages next to a free carboxyl groups126. Depolymerizing enzymes are known to act on different forms of pectin such as polygalacturonase (PG), pectatelyase (PL). The backbone of pectin consists of a smooth homogalacturonan, (1à4) linked α-D-galacturonic acid, which is methyl esterified at C-6 position. The PME is also related to increase in the firmness of fruits and vegetables by demethylation of endogenous pectin and chelation of divalent cations by ionized carboxyl groups on adjacent pectic acids60. The activity of PME has been detected in some bacteria, fungi, yeast and higher plants. The PME has been identified in several higher plants such as apple, banana, citrus (lime, orange, grape fruit), mango, papaya, cherry, berries, grape, pear, plum, beans, carrot, cauliflower, onion, pea, potato, radish and tomato127.
In the cell wall of dicot plants, about 35% of the dry weight consists of pectin. There are two phenomena involved in the firming effect of pectin in tissues i.e., the formation of free carboxyl groups in fresh tissue, which increases the strength of calcium binding between polymers of pectin while in heated tissue, calcium binding strength is increased but there is a decrease in the susceptibility of pectin to depolymerization by β-elimination. Generally the optimum pH for plants and bacteria is between 6 and 8 whereas some fungal PME have been to show pH optima between128 4 and 6.
The expression and characterization of PME in coffee fruit have been studied comprehensively129. Enzymatic activity of PME has been measured at different stages of maturation in coffee fruit, which showing PME activity to be increased progressively from the beginning of ripening process129. Five isoforms of PME have been identified in coffee and the transcription level of CaPME4 has been observed to increase in pericarp whereas decrease in endosperm129. The expression of CaPME4 transcript was effective in branches roots and flower tissues, indicating that it plays a significant role in pectin degradation and contributes to fruit softening. Recently, the continuum electrostatic calculations have been observed on the molecular dynamics trajectory of a PME-HG-disaccharide complex130. It has been reported that when methyl ester group of disaccharide were arranged at the carboxylate product of demethyl esterification, a net unidirectional sliding of disaccharide model was observed along the binding groove of enzyme130. It was also shown that during the translocation, the changes in electrostatic binding energies and protein dynamics reflect the mechanism of rectifying Brownian motion of enzymes130.
The PME has been reported in all higher plants particularly in citrus fruits131. It can also be produced by some pathogenic bacteria and fungi132. The PME is a cell wall bound enzyme and exists in two or more isoforms in higher plants but have different molecular weight, biochemical activity, physical and chemical characteristics133. The kinetic properties of enzymes are highly affected by the source e.g., the removal of pectin methyl ester group linearly by plants PMEs and bacterial PMEs from Erwinia chrysanthemi. In this type of action pattern, the binding of enzyme responsible for conversion of all subsequent substrate sites on the polymer chain, whereas fungal PMEs from Aspergillus niger acted on pectin with multiple chain mechanism. In this pattern, for each complex of enzyme and substrate molecules, the enzyme catalyses limited number of substrate site randomly134. It was reported that acidic PMEs were related with random de-esterification, whereas alkaline PMEs being related with linear de-esterification135. The mode of de-esterification of PME depends on the degree of methyl esterification of pectin and pH of the medium125.
Mode of action and regulation of PME: It has already been reported that the various isoforms of PME detected in cell walls are encoded by multigene family125. The sequencing of Arabidopsis genome, involves in the identification of 67 genes in relation with PME136. The PME genes encode some peptide motifs as pre-pro-proteins. The pre- region behaves as a signal peptide, which is essential for the targeting of proteins into the endoplasmic reticulum. Secretion of pro-PME leads to the apoplasm through the cis, medial and transgolgi cisternae and in the cell wall, only mature PME i.e., without pro-region is found. Through the study of systematic sequencing of Arabidopsis genome136, PME genes can be classified into two groups. The first group of gene consists of only two or three introns and a long pro-region whereas the genes of other group contain five or six introns and a small or non-existant pro-region125. These two groups of PME are designated as type I and type II respectively.
The structure of type II sequence is closely resembled to PMEs, occurring in pathogenic organisms, namely, bacteria, fungi, which are responsible in cell wall soaking during plant infection. It has been stated that cell wall extracted PMEs do not have a pro-region137. In mature PME, the cleavage of pro-region might occur in early or before excretion of mature PME or later into the apoplasm. The role of pro-region is exactly not known but there are some possible hypotheses. It may be involved in the biological functions of PMEs, in PMEs targeting and as an intramolecular chaperones, which help in protein folding or conformational change of PME138 or may be involved in the inhibition of enzyme activity137. The isoforms of PME, which are involved in microsporogenesis and pollen tube growth are type II related to bacterial PME138.
The mode of action of PME after integration into the cell wall is different. The main accepted hypothesis states that PMEs acts on homogalacturonan either randomly (as in fungi) or linearly (as in plants) along the chain of pectins137,138. When PMEs act on homogalacturonans randomly, demethyl esterification releases proton, which promotes the activity of endopolygalacturonases (endo-PG)137 and results in cell wall loosening. On the other hand, when PMEs act on homogalacturonans linearly, it produces blocks of free carboxyl groups, which interacts with calcium and creating a pectate gel137. The activity of endo-PG is limited in this gel and hence contributes to cell wall stiffening. The random demethylesterification depends on acidic PMEs while linear demethylesterification depends on alkaline PME138.
FUNCTIONS OF PME
In plant growth: There are various physiological processes of vegetative and reproductive plant development, in which PMEs play important role such as in cell wall extension and stiffening139, separation of cellular compartments140, germination of seed141, leaf growth142, growth of internode stem143, elongation of root tip144, dehiscence of dry fruit and in fruit ripening145-148. In tomato fruit, the activity of PME changes to modify the cell wall structure of the pericarp52. The PME is also involved in wood development. There are five different PME encoded genes, which are highly expressed in the xylem tissues of Arabidopsis. A large number of transcripts encoded by PME gene are found in poplar wood forming tissues149 and is tightly regulated within the cambial meristem tissue. The PME is also involved in the regulation of length of fibre by improving the cellular adhesion between the fibres and hence their apical elongation is inhibited. PME also plays a major role in modification of pectin in xylem during secondary wall deposition with the help of polygalacturonases and pectatelyases149,150. The significant role of PME is de-esterification of HGA, which is essential for the lignifications of xylem, e.g., de-esterified HGA calcium bridge is bound with peroxidase class III, which may start the polymerization of lignin151. PME helps in the de-esterification of HGA and ultimately initiates the process of lignifications at cell junctions in woody tissues152. Pectin molecules interact with lignin and affect their polymerization in vitro, which depicts the function of PME in regulating such interactions in vivo. The PME plays a significant role in reproductive development mainly in pollen formation and in pollen tube growth153. It has been shown that QUARTET1 is encoded by AT5G5590 in Arabidopsis is expressed in pollen and helps in the separation of pollen tetrad during floral development and when it is expressed in E. coli, it has PME activity154.
It has been suggested that in tobacco plant the control of degree of pectin esterification control oscillatory pollen tube growth155,156. In some symbiotic stages, various pollen specific PME proteins are present in roots, which are observed by the study of polar tip growth in the development of pollen tube and during rhizobial infestation of medicago, by the presence of infection threads157. The expression of PMEs, which are nodule specific as well as plant tissue specific having different similarities during the development of nodules, with higher activity in primordial than in mature nodules157,158.
In plant defense system: There is a physical barrier between internal compartments of plant cell and environment, plant defense mechanism or response is related with changes in cell wall159. It has been revealed that the expression of about 75% PME sequences in Arabidopsis differ in their response to both biotic and abiotic stresses e.g., the regulation of some PME transcripts is by wounding, ethylene160, cold161, oligogalacturonides (OGAs)162 and phloem feeding insects162. In over-expressing plant lines and in antisense, the activity of PME has been observed to up and down and the change in degree of methylesterification (DM) of pectin have been related with changes in susceptibility of plants to pathogens and abiotic stresses163,164. The PMEs may be involved either directly by the interaction with virulence factors or indirectly by catalyzing the reaction products. The functional plant PME binding is required to the Movement Proteins (MPs) of some viruses for the transport of them to the new cells with the help of plasmodesmata165. Generally PMEs are related with cell wall embedded plasmodesmata, MPs co localize with plasmodesmata and PME like proteins bind to MPs but the mechanism that allows PME for viral movement is not clear. The activity of PME enzyme produces three types of compounds, which are involved in defense mechanism in plants.
The first one is the production of methanol that has been observed in the interaction between plant and herbivores166,167. The rapid increases in the release of methanol have been observed in tobacco after wounding and in the presence of oral secretions of caterpillars, they get enhanced, which leads to the activity as well as expression of PME becomes up regulated168. As the activity of PME depends on pH, the release of methanol is also pH dependent. The methanol may either diffuse out or may be broken down into formate, but act as a substrate for the enzyme formate dehydrogenase, which is up regulated by environmental stresses such as cold, wounding, drought in leaves of potato169 and by the spray of methanol. The regulation of PME genes have also been found byaphids present in phloem of celery168 that spectacles that the methanol produced by PME plays a role in the signaling of plant stress.
Secondly, the activity of PME leads to release of carboxylic group, a negatively charged particle and protons in the matrix of pectin144, which modifies the charge of cell wall, pH of apoplasm and the activity of proteins of apoplasm. By the activity of PME, negatively charges produced, that facilitates the binding of cations, which either inhibits or favors the growth of plant depending on conditions144. It has been shown that the activity of PME can also change the content of aluminium, which is a toxic metal in soil that inhibits the elongation of roots163. With the help of this study, it has been shown that the amount of aluminium in plant increases on the over-expression of PME activity which results in the decrease in degree of pectin methylesterification. Like other cations, namely, calcium or magnesium, aluminium is also capable to enhance the activity of PME in vivo. The high activity of PME in cell wall results in the high amount of unmethylesterified HGA, which has been positively related with susceptibility of plants to biotic and abiotic stresses163.
Finally, third category includes various compounds, which are derived from pectin, involving in plant defense system. The glycosidic bonds of polygalacturonic acid can be cleaved by glycosidic hydrolases e.g., polygalacturonases which allows the action of pectatelyase and formation of oligogalacturonic acid. The pectin might be depolymerizing by pectin lyases through breaking the bonds between methylesterified galacturonide residues. The defense of plants induced by oligogalcturonides has been studied and its affectivity depends on the degree of oligogalacturonide methylesterification159,162. By the activity of PME, there is an acidification of apoplasm, which leads to the formation of oligogalacturonides by pectolytic enzymes. Exogenous pectolytic enzymes are also involved in plant-pathogen interactions. There are various secretions by pests and pathogens of various enzymes including PME through plants when attack by bacteria, fungi and insects138,159,170 e.g., a pathogenicity has been determined by the functional PME from the bacteria Botrytis cinerea171. From cell wall digest of T. aestivum, there is a release of dimmers or trimers of galacturonic acid which are endogenous suppressor of defense mechanism. So, in the interactions of plant and pathogens, some compounds, derived from HGA, act as elicitors while others participate as defense suppressors, depending on the pectolytic enzymes involved159,162,172. The susceptibility of a plant to the stress may be modulated by the mode of PME action. Patterns of methylesterification of HGA are found to be significantly variations in between stem rust fungus resistant, in which blockwise distribution of methyl esters takes place and susceptible lines of wheat, in which non-blockwise distribution occurs.
The activity of PME is significant for the virulence of necrotrophic fungal pathogen Botrytis cinerea. The PMEs from Arabidopsis thaliana are involved in Pathogen Triggered Immunity (PTI) and immune responses to Alternaria brassicicola, a necrotrophic fungus and to Pseudomonas syringae bacteria173. It has been reported that after the inoculation with either pathogen and during PTI, the activity of plant PME is increased173. The activity of pathogen induced PME does not necessitate ethylene signaling but depending on Jasmonic Acid (JA) signaling173. About 66 PME genes have been reported in Arabidopsis, suggesting the extensive genetic redundancy.
It has been studied that during the interaction of Fusarium and banana, PME modifies the pectin174. Currently two varieties of bananas have been studied for the expression and activity of PME in such a way that one variety is highly resistant whereas other one is highly susceptible to Fusarium174. It has been observed that before the treatment of pathogen, the activity of PME is higher in resistant cultivar than susceptible cultivar174. It was also showed that when the plants were wounded, there was a significant increase in the expression of PME and a decrease in degree of methylesterification in susceptible cultivar but not in the resistant variety174.
In pollen tube growth: When pollen tube growth media along with orange peel PME was studied, the results showed that the supply of exogenous PME inhibited the growth in vitro154,156,175 and cause the apical cell wall thickening and dissipation of the intracellular tip-focused Ca2+ gradient155,156. On the exposure of pollen tubes with the enhanced activities of extracellular PME changes the apical methylesterified pectins into deesterified forms175, which ultimately form a calcium pectate gel at the tip, preventing elongation and causing cell wall thickening. Thus, these results indicate that the composition of pectin by PME is an important component in controlling the growth of pollen tube at the apex. When various mutant tubes grown in vitro, they burst by decrease in the overall activity of PME, which leads to an imbalance of apical wall and results in failure to resist the internal turgor pressure. The in vivo growth of mutant pollen tubes initially are not affected on the surface of stigma cells but the growth is rapidly decreased in the style. The rate of pollen tube growth oscillates between in vitro176 and in vivo177. At the tip, the mechanism of regulation of PME activity consists of a negative feedback, where the locally generated pH decreases by the release of protons during de-esterification, reducing the activity of PME. Polygalacturonases and pectatelyase enzymes are also expected to be activated at low pH. The synergistic effect of inactivating PME and activating PG and PL causes the loosening of cell wall, facilitating a growth pulse. Due to the saturation of negative charges, there is an increase in pH and responsible for reactivation of PME and further inactivation of PG and PL, which leads to stiffening of apical wall and results in decrease in growth rate. Thus, pectin associated enzymes with PME, play a significant role in controlling oscillations in the growth of pollen tube177.
There are a number of evidences in the support of this hypothesis. One of them is the thickness of pollen tubes in lily where the frequency of oscillations is shown to be same as growth, becomes more thickened in anticipation of increase of growth rate176. In other case, the transcript of PG and PL are highly expressed in pollen of Arabidopsis178 that performs a role in pollen germination and tube growth. It has been reported that the pollen germination and tube growth is stimulated by the application of PG from Aspergillus niger175. Besides the regulation of PME activity is controlled by endogenous oscillation of intracellular H+. There are two domains in pollen tubes, which oscillate in pH intensity i.e., at the apex, acidic domain while at the base, an alkaline band of clear zone179. At the apex, the acidic tip is maintained by the influx of proton whereas by the efflux of proton, alkaline band is to be maintained through the action of proton ATPase located in the cell membrane. Due to the fluxes of these protons, the pH of specific cell wall region is affected and thus probably influencing the PME activity in these regions.
Some specific proteases may be activated by the change in pH, which processes the pro-region and releases the active PME domain. It has been observed that at the tip of pollen tube there is an intracellular Ca2+ gradient and its concentration also oscillates. This gradient can be maintained by the influx of Ca2+ from extracellular region across the plasma membrane. Thus, it has been concluded that through the same activated cation channels, both Ca2+ and H+ can enter at the tip176. These generated ion fluxes may affect the activity of PME.
MOLECULAR MECHANISM AND SIGNAL TRANSDUCTION DURING FRUIT RIPENING
The development of fruit requires a network of interacting genes and signaling pathways. Fruit ripening constitutes several biochemical processes, including change in color, sugar, acidity, texture and aroma and are important for the quality of fruit. At the end stages of ripening, some changes occur, which are related with the senescence caused membrane destruction and leads to cell death. Thus, fruit ripening can be assumed as the first step of programmed cell death. During fruit ripening, all biochemical as well as physiological changes take place by the coordinated expression of fruit ripening related genes. This includes encoding the genes for enzymes responsible for biochemical and physiological changes, functioning of regulatory proteins involving in signaling pathways and in the machinery of transcription ultimately regulating the expression of gene. The genetically regulated process of fruit ripening involves the activation of a large number of primary and secondary metabolic pathways, which involve in sensory and nutritional quality of fruit. The complete process of expression of ripening related genes are under control of hormonal and environmental signals, in which ethylene plays a significant role.
The overall molecular pathways during the process of ripening are shown in Fig. 5. There are two different systems for ethylene biosynthesis. System 1 is related with the low production of ethylene in pre-climacteric stage of climacteric fruit and is present throughout the development of non-climacteric fruit. On the other hand, system 2 is an autostimulated large production of ethylene and known as "Autocatalytic synthesis" and is specific for climacteric fruit. Thus, in climacteric and non-climacteric fruits, the major difference in relation with ethylene is the presence or absence of autocatalytic ethylene production88. In ethylene biosynthesis process, two main enzymes involved are ACC synthase (ACS), which converts SAM into ACC and ACC Oxidase (ACO), converting ACC into ethylene. The ACS and ACO genes are encoded by multigene family of 9 and 5 members respectively in tomato, along with their differentially expressions in fruit ripening and development180.
It was also studied that LeACO1 and LeACO4 genes are up-regulated at the time of ripening and continue being active throughout the process. The LeACO3 shows a transient activation, while LeACS6 and LeACS1A are expressed at the pre climacteric stage of system 1, LeACS4 and LeACS1A are most active genes at the time of transition of ripening180,181. LeACS4 expresses highly during climacteric phase and the expression of LeACS1A declines. At the time of ripening, the increased production of ethylene results in the induction of LeACS2 and the expression of LeACS6 and LeACS1A inhibit. Thus, ACS gene can be considered as critical for the switch from system 1 of pre-climacteric stage to system 2 of climacteric stage182. By using an antisense strategy in transgenic plants, down-regulating ACO and ACS genes is required for ethylene to trigger the ripening of climacteric fruit.
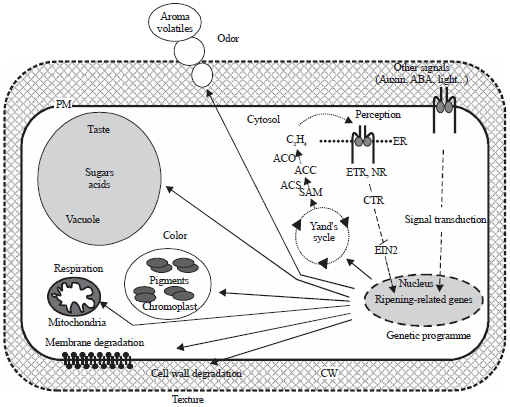 | |
| Fig. 5: | Molecular mechanism controlling the ripening of climacteric fruit182 |
In tomato, delayed ripening was monitored by ethylene-suppressed lines183 and in other fruits such as apple184 and melon185. In some climacteric fruit such as melon, ethylene-independent ripening pathway leads to softening, flesh color and accumulation of sugar occurs in ethylene suppressed fruit186. Based on these results, it has been concluded that climacteric (ethylene dependent) and non-climacteric (ethylene independent) regulation coexist in climacteric fruit187. In grape and berries, the supply of an exogenous ethylene stimulates the expression of genes related with the production of anthocyanin and involves in the regulation of vascular flux, acid content and some steps of volatile aroma synthesis188.
Ethylene perception has been shown on the model plant Arabidopsis and molecular genetics are implemented. The ethylene receptor genes were obtained by positional cloning189. The first isolated plant hormone receptor was ethylene receptor, reflecting towards the other components involving in ethylene transduction pathway189. The ethylene receptors are encoded by small multigene family, e.g., in tomato, six genes have been isolated for ethylene receptors, which are found to be expressed in all tissues of plant. Out of these six genes, three genes show a net increase on ripening whereas two genes are expressed constitutively. It has been demonstrated in recent studies that during fruit ripening, ethylene receptors are degraded rapidly and the rate of transcription remains high and that level of receptor determines the timing of ripening181.
In tomato fruit, the suppression of ethylene receptor LeETR4 leads to early ripening181. Another important component of ethylene signaling is CTR1 gene (Constitutive Triple Response), isolated first from Arabidopsis acts as a negative regulator of ethylene transduction pathway181. First isolated gene from fruit tissue of tomato has been recognized as CTR1 gene (S-1 CTR1)190 and its transcripts levels are known to be up-regulated during fruit ripening with increase in ethylene production.
BIOCHEMICAL AND in silico CHARACTERIZATION PECTIN METHYLESTERASE FROM G9 VARIETY OF Musa acuminata FOR DELAYED RIPENING
Amongst important cell wall degrading enzymes, pectin methylesterase (PME) has been known to act as a cell wall hydrolyzing enzyme, responsible for demethylesterification of cell wall polygalacturonan. A recent study has been successfully accomplished on the biochemical and molecular characterization of PME from Grand nain variety of Musa acuminata (banana)191. This study also deals with the in silico study reflecting inhibition of PME activity in context to delayed ripening in banana. It mainly deals with the identification of a PME1 gene from Grand nain variety of banana. The expression of this gene is related with the process of ripening. The expression of PME1 gene was observed to be peaked on 3rd day in ethylene treated samples of banana but the activity in untreated samples called control was rather slow and then there was a sudden decrease in their activity in both treated as well as untreated samples191. With the help of in silico study, authors observed that banana has maximum homology with carrot by using cross species analysis192. The designed model has been reported to be of good quality on the basis of its verification and validation and thus appropriate for docking. The information of binding sites of ligand provides new insights into the predictable functioning of relevant protein193.
CONCLUSION AND FUTURE RECOMMENDATIONS
How to delay fruit ripening is an important biological interest. During transport the fruits get harmed due to their early ripening. On fruit ripening there are many biochemical changes takes place like respiration, ethylene production and activity of cell wall degrading enzymes etc. There are many cell wall degrading enzymes which play an important role in fruit ripening namely PME, PG, PL etc. The PME is one of the cell wall degrading enzymes which act on pectin (a primary cell wall constituent) and releases methanol and hydrogen ions. Using cross species analysis, we found that banana has maximum homology with carrot. The validation and verification of the designed model has been performed and found that it is good quality model. This model is further used for docking studies. Knowledge about the site at which a ligand binds provides an important clue for predicting the function of a protein and is also often a prerequisite for performing docking computations in virtual drug design and screening. Docking results suggested that green tea catechin and salicin are the best inhibitors having good interaction energies which binds at third motif i.e., Asp381 residue on active site of PME in Musa acuminata. It was found that green tea catechin is better than salicin because it is a natural inhibitor, antioxidant and inhibits the activity of PME whereas salicin is a chemical compound inhibits ripening at some specific concentration only, although the binding energies of both the inhibitors have approximately similar. This review study provides new insights into increasing the more availability of banana through delaying its ripening by inhibiting especially the activity of pectin methylesterase.
SIGNIFICANCE STATEMENT
The major significance of this study is how to delay fruit ripening is an important biological interest. Never-the-less, this study provides new insights into increasing the more availability of banana through delaying its ripening by inhibiting especially the activity of pectin methylesterase.
REFERENCES
- Marin, D.H., T.B. Sutton and K.R. Barker, 1998. Dissemination of bananas in Latin America and the Caribbean and its relationship to the occurrence of Radophouls similis. Plant Dis., 82: 964-974.
Direct Link - Seymour, G.B., J.E. Taylor and G.A. Tucher, 1993. Biochemistry of Fruit Ripening. 1st Edn., Chapman and Hall, Dordrecht, The Netherlands, ISBN: 978-94-011-1584-1, Pages: 454.
Direct Link - Blake, J.D. and G.N. Richards, 1971. An examination of some methods for fractionation of plant hemicelluloses. Carbohydr. Res., 17: 253-268.
CrossRefDirect Link - McNiel, M., P. Albersheim, L. Taiz and R.L. Jones, 1975. The structure of plant cell walls: VII. Barley aleurone cells. Plant Physiol., 55: 64-68.
CrossRefDirect Link - Ridley, B.L., M.A. O'Neill and D. Mohnen, 2001. Pectins: Structure, biosynthesis and oligogalacturonide-related signaling. Phytochemistry, 57: 929-967.
CrossRefDirect Link - O'Neill, M.A., T. Ishii, P. Albersheim and A.G. Darvill, 2004. Rhamnogalacturonan II: Structure and function of a borate cross-linked cell wall pectic polysaccharide. Annu. Rev. Plant Biol., 55: 109-139.
CrossRefDirect Link - Van Buren, J.O., 1979. The chemistry of texture in fruits and vegetables. J. Texture Stud., 10: 1-23.
CrossRefDirect Link - Thakur, B.R., R.K. Singh, A.K. Handa and M.A. Rao,, 1997. Chemistry and uses of pectin-A review. Crit. Rev. Food. Sci. Nutr., 37: 47-73.
CrossRefPubMedDirect Link - Jarvis, M.C., 1984. Structure and properties of pectin gels in plant cell walls. Plant Cell Environ., 7: 153-164.
CrossRefDirect Link - McNeil, M., A.G. Darvill, S.C. Fry and P. Albersheim, 1984. Structure and function of the primary cell walls of plants. Annu. Rev. Biochem., 53: 625-663.
CrossRefDirect Link - Jin, D.F. and C.A. West, 1984. Characteristics of galacturonic acid oligomers as elicitors of casbene synthetase activity in castor bean seedlings. Plant Physiol., 74: 989-992.
CrossRefDirect Link - Robertsen, B., 1986. Elicitors of the production of lignin-like compounds in cucumber hypocotyls. Physiol. Mol. Plant Pathol., 28: 137-148.
CrossRefDirect Link - Laurent, M.A. and P. Boulenguer, 2003. Stabilization mechanism of Acid Dairy Drinks (ADD) induced by pectin. Food Hydrocolloids, 17: 445-454.
CrossRefDirect Link - Kastner, U., S. Glasl, B. Follrich, J.P. Guggenbichler and J. Jurenitsch, 2002. [Acid oligosaccharides as the active principle of aqueous carrot extracts for prevention and therapy of gastrointestinal infections]. Wiener Medizinische Wochenschrift, 152: 379-381, (In German).
CrossRefPubMedDirect Link - Lim, B.O., K. Yamada, M. Nonaka, Y. Kuramoto, P. Hung and M. Sugano, 1997. Dietary fibers modulate indices of intestinal immune function in rats. J. Nutr., 127: 663-667.
Direct Link - Langhout, D.J., J.B. Schutte, P. van Leeuwen, J. Wiebenga and S. Tamminga, 1999. Effect of dietary high- and low-methylated citrus pectin on the activity of the ileal microflora and morphology of the small intestinal wall of broiler chicks. Br. Poult. Sci., 40: 340-347.
CrossRefDirect Link - Trumbo, P., S. Schlicker, A.A. Yates and M. Poos, 2002. Dietary reference intakes for energy, carbohydrate, fiber, fat, fatty acids, cholesterol, protein and amino acids. J. Am. Dietetic Assoc., 102: 1621-1630.
CrossRefPubMedDirect Link - McNeil, M., A.G. Darvill and P. Albersheim, 1979. The Structural Polymers of the Primary Cell Walls of Dicots. In: Progress in the Chemistry of Organic Natural Products, Volume 37, Herz, W., H. Grisebach and G.W. Kirby (Eds.). Springer-Verlag, Vienna, Austria, ISBN: 978-3-7091-8547-6, pp: 191-249.
Direct Link - De Vries, J.A., A.G.J. Voragen, F.M. Rombouts and W. Pilnik, 1981. Extraction and purification of pectins from Alcohol Insoluble Solids from ripe and unripe apples. Carbohydr. Polym., 1: 117-127.
CrossRefDirect Link - O'Neill, M.A., S. Eberhard, P. Albersheim and A.G. Darvill, 2001. Requirement of borate cross-linking of cell wall rhamnogalacturonan II for Arabidopsis growth. Science, 294: 846-849.
CrossRefDirect Link - Mohnen, D., R.L. Doong, K. Liljebjelke, G. Fralish and J. Chan, 1996. Cell Free Synthesis of the Pectic Polysaccharide Homogalacturonan. In: Pectins and Pectinases (Progress in Biotechnology, Volume 14), Visser, J. and A.G.J. Voragen (Eds.). Elsevier Science BV, Amsterdam, ISBN-13: 9780080544601, pp: 109-127.
- McNeil, M., A. Darvill and P. Albersheim, 1980. Structure of plant cell walls. X. Rhamnogalacturonan I, a structurally complex pectic polysaccharide in the walls of suspension-cultured sycamore cells. Plant Physiol., 66: 1128-1134.
CrossRefDirect Link - Mort, A.J., F. Qiu and N.O. Maness, 1993. Determination of the pattern of methyl esterification in pectin. Distribution of contiguous nonesterified residues. Carbohydr. Res., 247: 21-35.
CrossRefDirect Link - Albersheim, P., A.G. Darvill, M.A. O'Neill, H.A. Schols and A.G.J. Voragen, 1996. An Hypothesis: The Same Six Polysaccharides are Components of the Primary Cell Walls of all Higher Plants. In: Pectins and Pectinases (Progress in Biotechnology, Volume 14), Visser, J. and A.G.J. Voragen (Eds.). Elsevier, London, UK., ISBN-13: 9780080544601, pp: 47-56.
- Schols, H.A., E.J. Bakx, D. Schipper and A.G.J. Voragen, 1995. A xylogalacturonan subunit present in the modified hairy regions of apple pectin. Carbohydr. Res., 279: 265-279.
CrossRefDirect Link - Le Goff, A., C.M.G.C. Renard, E. Bonnin and J.F. Thibault, 2001. Extraction, purification and chemical characterisation of xylogalacturonans from pea hulls. Carbohydr. Polym., 45: 325-334.
CrossRefDirect Link - Yu, L. and A.J. Mort, 1996. Partial Characterization of Xylogalacturonans from Cell Walls of Ripe Watermelon Fruit: Inhibition of Endopolygalacturonase Activity by Xylosylation. In: Pectins and Pectinases (Progress in Biotechnology, Volume 14), Visser, J. and A.G.J. Voragen (Eds.). Elsevier, London, UK., ISBN-13: 9780080544601, pp: 79-88.
- Zandleven, J., S.O. Sorensen, J. Harholt, G. Beldman, H.A. Schols, H.V. Scheller and A.J. Voragen, 2007. Xylogalacturonan exists in cell walls from various tissues of Arabidopsis thaliana. Phytochemistry, 68: 1219-1226.
CrossRefDirect Link - Colquhoun, I.J., G.A. de Ruiter, H.A. Schols and A.G.J. Voragen, 1990. Identification by N.M.R. spectroscopy of oligosaccharides obtained by treatment of the hairy regions of apple pectin with rhamnogalacturonase. Carbohydr. Res., 206: 131-144.
CrossRefDirect Link - Kravtchenko, T.P., I. Arnould, A.G.J. Voragen and W. Pilnik, 1992. Improvement of the selective depolymerization of pectic substances by chemical β-elimination in aqueous solution. Carbohydr. Polym., 19: 237-242.
CrossRefDirect Link - Rihouey, C., C. Morvan, I. Borissova, A. Jauneau, M. Demarty and M. Jarvis, 1995. Structural features of CDTA-soluble pectins from flax hypocotyls. Carbohydr. Polym., 28: 159-166.
CrossRefDirect Link - Carpita, N.C. and D.M. Gibeaut, 1993. Structural models of primary cell walls in flowering plants: consistency of molecular structure with the physical properties of the walls during growth. Plant J., 3: 1-30.
CrossRefPubMedDirect Link - Schols, H.A. and A.G.J. Voragen, 1994. Occurrence of pectic hairy regions in various plant cell wall materials and their degradability by rhamnogalacturonase. Carbohydr. Res., 256: 83-95.
CrossRefDirect Link - Ishii, T. and T. Matsunaga, 2001. Pectic polysaccharide rhamnogalacturonan II is covalently linked to homogalacturonan. Phytochemistry, 57: 969-974.
CrossRefDirect Link - Huisman, M.M.H., L.P. Brull, J.E. Thomas-Oates, J. Haverkamp, H.A. Schols and A.G.J. Voragen, 2001. The occurrence of internal (1→5)-linked arabinofuranose and arabinopyranose residues in arabinogalactan side chains from soybean pectic substances. Carbohydr. Res., 330: 103-114.
CrossRefDirect Link - Gaspar, Y., K.L. Johnson, J.A. McKenna, A. Bacic and C.J. Schultz, 2001. The complex structures of arabinogalactan-proteins and the journey towards understanding function. Plant Mol. Biol., 147: 161-176.
CrossRefDirect Link - Vincken, J.P., H.A. Schols, R.J.F.J. Oomen, G. Beldman, R.G.F. Visser and A.G.J. Voragen, 2003. Pectin-the Hairy Thing: Evidence that Homogalacturonan is a Side Chain of Rhamnogalacturonan I. In: Advances in Pectin and Pectinase Research, Voragen, A.G.J., H.A. Schols and R.G.F. Visser (Eds.). Kluwer Academic Publishers, Dordrecht, pp: 47-60.
- Immerzeel, P., M.M. Eppink, S.C. de Vries, H.A. Schols and A.G.J. Voragen, 2006. Carrot arabinogalactan proteins are interlinked with pectins. Physiologia Plantarum, 128: 18-28.
CrossRefDirect Link - Brummell, D.A. and M.H. Harpster, 2001. Cell wall metabolism in fruit softening and quality and its manipulation in transgenic plants. Plant. Mol. Biol., 47: 311-339.
CrossRefDirect Link - Cosgrove, D.J., 2000. Loosening of plant cell walls by expansins. Nature, 407: 321-326.
CrossRefDirect Link - Whitney, S.E.C., M.J. Gidley and S.J. McQueen-Mason, 2000. Probing expansin action using cellulose/hemicellulose composites. Plant J., 22: 327-334.
CrossRefDirect Link - Anjanasree, K.N. and K.C. Bansal, 2003. Isolation and characterization of ripening-related expansin cDNA from tomato. J. Plant Biochem. Biotechnol., 12: 31-35.
CrossRefDirect Link - Seymour, T.A., J.F. Preston, L. Wicker, J.A. Lindsay and M.R. Marshall, 1991. Purification and properties of pectinesterases of Marsh white grapefruit pulp. J. Agric. Food Chem., 39: 1080-1085.
CrossRefDirect Link - Poovaiah, B.W., 1986. Role of calcium in prolonging storage life of fruits and vegetables. Food Technol., 40: 86-89.
Direct Link - Alonso, J., N. Howell and W. Canet, 1997. Purification and characterisation of two pectinmethylesterase from persimmon (Diospyros kaki). J. Sci. Food Agric., 75: 352-358.
CrossRefDirect Link - Rexova-Benkova, L. and O. Markovic, 1976. Pectic enzymes. Adv. Carbohydr. Chem., 33: 323-385.
CrossRefDirect Link - Markovie, O. and R. Kohn, 1984. Mode of pectin deesterification by Trichoderma reesei pectinesterase. Experientia, 40: 842-843.
CrossRefDirect Link - Richard, L., L.X. Qin and R. Goldberg, 1996. Clustered genes within the genome of Arabidopsis thaliana encoding pectin methylesterase-like enzymes. Gene, 170: 207-211.
CrossRefDirect Link - Micheli, F., C. Holliger, R. Goldberg and L. Richard, 1998. Characterization of the pectin methylesterase-like gene AtPME3: A new member of a gene family comprising at least 12 genes in Arabidopsis thaliana. Gene, 220: 13-20.
CrossRefDirect Link - Abu-Sarra, A.F. and A.A. Abu-Goukh, 1992. Changes in pectinesterase, polygalacturonase and cellulase activity during mango fruit ripening. J. Hortic. Sci., 67: 561-568.
CrossRefDirect Link - Smith, C.J.S., C.F. Watson, J. Ray, C.R. Bird, P.C. Morris, W. Schuch and D. Grierson, 1988. Antisense RNA inhibition of polygalacturonase gene expression in transgenic tomatoes. Nature, 334: 724-726.
CrossRefDirect Link - Tieman, D.M., R.W. Harriman, G. Ramamohan and A.K. Handa, 1992. An antisense pectin methylesterase gene alters pectin chemistry and soluble solids in tomato fruit. Plant Cell, 4: 667-679.
CrossRefDirect Link - Brummell, D.A., M.H. Harpster and P. Dunsmuir, 1999. Differential expression of expansin gene family members during growth and ripening of tomato fruit. Plant Mol. Biol., 39: 161-169.
CrossRefDirect Link - Smith, D.L., J.A. Abbott and K.C. Gross, 2002. Down-regulation of tomato β-galactosidase 4 results in decreased fruit softening. Plant Physiol., 129: 1755-1762.
CrossRefDirect Link - Brummell, D.A., M.H. Harpster, P.M. Civello, J.M. Palys, A.B. Bennett and P. Dunsmuir, 1999. Modification of expansin protein abundance in tomato fruit alters softening and cell wall polymer metabolism during ripening. Plant Cell, 11: 2203-2216.
CrossRefDirect Link - Jimenez-Bermudez, S., J. Redondo-Nevado, J. Munoz-Blanco, J.L. Caballero and J.M. Lopez-Aranda et al., 2002. Manipulation of strawberry fruit softening by antisense expression of a pectate lyase gene. Plant Physiol., 128: 751-759.
CrossRefDirect Link - Hirschberg, J., 2001. Carotenoid biosynthesis in flowering plants. Curr. Opin. Plant Biol., 4: 210-218.
CrossRefDirect Link - Winkel-Shirley, B., 2001. Flavonoid biosynthesis. A colorful model for genetics, biochemistry, cell biology and biotechnology. Plant Physiol., 126: 485-493.
CrossRefDirect Link - El-Kereamy, A., C. Chervin, J.P. Roustan, V. Cheynier and J.M. Souquet et al., 2003. Exogenous ethylene stimulates the long-term expression of genes related to anthocyanin biosynthesis in grape berries. Physiologia Plantarum, 119: 175-182.
CrossRefDirect Link - Mol, J., G. Jenkins, E. Schafer, D. Weiss and V. Walbot, 1996. Signal perception, transduction and gene expression involved in anthocyanin biosynthesis. Crit. Rev. Plant Sci., 15: 525-557.
CrossRefDirect Link - El-Sharkawy, I., D. Manriquez, F.B. Flores, F. Regad, M. Bouzayen, A. Latche and J.C. Pech, 2005. Functional characterization of a melon alcohol acyl-transferase gene family involved in the biosynthesis of ester volatiles. Identification of the crucial role of a threonine residue for enzyme activity. Plant Mol. Biol., 59: 345-362.
CrossRefPubMedDirect Link - Thompson, G.A. and F.L. Goggin, 2006. Transcriptomics and functional genomics of plant defence induction by phloem-feeding insects. J. Exp. Bot., 57: 755-766.
CrossRefDirect Link - Hopkins, W.G. and N.P.A. Hunter, 2004. Introduction to Plant Physiology. 3rd Edn., John Wiley and Sons Inc., New York, ISBN: 978-0-471-38915-6, Pages: 576.
Direct Link - Whitehead, C.S. and C.A. Bosse, 1991. The effect of ethylene and short-chain saturated fatty acids on ethylene sensitivity and binding in ripening bananas. J. Plant Physiol., 137: 358-362.
CrossRefDirect Link - Saltveit, M.E., 2005. Aminoethoxyvinylglycine (AVG) reduces ethylene and protein biosynthesis in excised discs of mature-green tomato pericarp tissue. Postharvest Biol. Technol., 35: 183-190.
CrossRefDirect Link - Alexander, L. and D. Grierson, 2002. Ethylene biosynthesis and action in tomato: A model for climacteric fruit ripening. J. Exp. Bot., 53: 2039-2055.
CrossRefPubMedDirect Link - Do Nascimento, J.R.O., A. Vieira Jr., P.Z. Bassinello, B.R. Cordenunsi, J.A. Mainardi, E. Purgatto and F.M. Lajolo, 2006. Beta-amylase expression and starch degradation during banana ripening. Postharvest Biol. Technol., 40: 41-47.
CrossRefDirect Link - Blankenship, S.M. and J.M. Dole, 2003. 1-Methylcyclopropene: A review. Postharvest Biol. Technol., 28: 1-25.
CrossRefDirect Link - Watkins, C.B., 2006. The use of 1-Methylcyclopropene (1-MCP) on fruits and vegetables. Biotechnol. Adv., 24: 389-409.
CrossRefPubMedDirect Link - Serek, M., E.J. Woltering, E.C. Sisler, S. Frello and S. Sriskandarajah, 2006. Controlling ethylene responses in flowers at the receptor level. Biotechnol. Adv., 24: 368-381.
CrossRefDirect Link - Gupta, S.M., S. Srivastava, A.P. Sane and P. Nath, 2006. Differential expression of genes during banana fruit development, ripening and 1-MCP treatment: Presence of distinct fruit specific, ethylene induced and ethylene repressed expression. Postharvest Biol. Technol., 42: 16-22.
CrossRefDirect Link - Kesari, R., P.K. Trivedi and P. Nath, 2007. Ethylene-induced ripening in banana evokes expression of defense and stress related genes in fruit tissue. Postharvest Biol. Technol., 46: 136-143.
CrossRefDirect Link - Bleecker, A.B. and H. Kende, 2000. Ethylene: A gaseous signal molecule in plants. Annu. Rev. Cell Dev. Biol., 16: 1-18.
CrossRefDirect Link - Pirrello, J., F. Jaimes-Miranda, M.T. Sanchez-Ballesta, B. Tournier and Q. Khalil-Ahmad et al., 2006. Sl-ERF2, a tomato ethylene response factor involved in ethylene response and seed germination. Plant Cell Physiol., 47: 1195-1205.
CrossRefDirect Link - Riechmann, J.L., J. Heard, G. Martin, L. Reuber and C.Z. Jiang et al., 2000. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science, 290: 2105-2110.
CrossRefPubMedDirect Link - Rosado, A., A.L. Schapire, R.A. Bressan, A.L. Harfouche, P.M. Hasegawa, V. Valpuesta and M.A. Botella, 2006. The Arabidopsis tetratricopeptide repeat-containing protein TTL1 is required for osmotic stress responses and abscisic acid sensitivity. Plant Physiol., 142: 1113-1126.
CrossRefDirect Link - Stepanova, A.N., J. Yun, A.V. Likhacheva and J.M. Alonso, 2007. Multilevel interactions between ethylene and auxin in Arabidopsis roots. Plant Cell, 19: 2169-2185.
CrossRefDirect Link - Jones, B., P. Frasse, E. Olmos, H. Zegzouti and Z.G. Li et al., 2002. Down-regulation of DR12, an auxin-response-factor homolog, in the tomato results in a pleiotropic phenotype including dark green and blotchy ripening fruit. Plant J., 32: 603-614.
CrossRefDirect Link - Krall, S.M. and R.F. McFeeters, 1998. Pectin hydrolysis: Effect of temperature, degree of methylation, pH and calcium on hydrolysis rates. J. Agric. Food Chem., 46: 1311-1315.
CrossRefDirect Link - Nogata, Y., H. Ohta and A.G.J. Voragen, 1993. Polygalacturonase in strawberry fruit. Phytochemistry, 34: 617-620.
CrossRefDirect Link - Redgwell, R.J., L.D. Melton and D.J. Brasch, 1992. Cell wall dissolution in ripening kiwifruit (Actinidia deliciosa): Solubilization of the pectic polymers. Plant Physiol., 98: 71-81.
CrossRefDirect Link - Femenia, A., E.S. Sanchez, S. Simal and C. Rossello, 1998. Developmental and ripening‐related effects on the cell wall of apricot (Prunus armeniaca) fruit. J. Sci. Food Agric., 77: 487-493.
CrossRefDirect Link - Paull, R.E. and N.J. Chen, 1983. Postharvest variation in cell wall-degrading enzymes of papaya (Carica papaya L.) during fruit ripening. Plant Physiol., 72: 382-385.
CrossRefDirect Link - Roe, B. and J.H. Bruemmer, 1981. Changes in pectic substances and enzymes during ripening and storage of 'Keitt' mangos. J. Food Sci., 46: 186-189.
CrossRefDirect Link - Boothby, D., 1983. Pectic substances in developing and ripening plum fruits. J. Sci. Food Agric., 34: 1117-1122.
CrossRefDirect Link - Gray, J., S. Picton, J. Shabbeer, W. Schuch and D. Grierson, 1992. Molecular biology of fruit ripening and its manipulation with antisense genes. Plant Mol. Biol., 19: 69-87.
PubMedDirect Link - Smith, B.G. and P.J. Harris, 1995. Polysaccharide composition of unlignified cell walls of pineapple [Ananas comosus (L.) Merr.] fruit. Plant Physiol., 107: 1399-1409.
CrossRefDirect Link - Prasanna, V., T.N. Prabha and R.N. Tharanathan, 2007. Fruit ripening phenomena-an overview. Crit. Rev. Food Sci. Nutr., 47: 1-19.
CrossRefPubMedDirect Link - Huber, D.J., 1984. Strawberry fruit softening: The potential roles of polyuronides and hemicelluloses. J. Food Sci., 49: 1310-1315.
CrossRefDirect Link - Prabha, T.N. and N. Bhagyalakshmi, 1998. Carbohydrate metabolism in ripening banana fruit. Phytochemistry, 48: 915-919.
CrossRefDirect Link - Pathak, N. and G.G. Sanwal, 1998. Multiple forms of polygalacturonase from banana fruits. Phytochemistry, 48: 249-255.
CrossRefDirect Link - Pathak, N., S. Mishra and G.G. Sanwal, 2000. Purification and characterization of polygalacturonase from banana fruit. Phytochemistry, 54: 147-152.
CrossRefDirect Link - Whitaker, J.R., 1984. Pectic substances, pectic enzymes and haze formation in fruit juices. Enzyme Microb. Technol., 6: 341-349.
CrossRefDirect Link - Alkorta, I., C. Garbisu, M.J. Llama and J.L. Serra, 1998. Industrial applications of pectic enzymes: A review. Process Biochem., 33: 21-28.
CrossRefDirect Link - Micheli, F., 2001. Pectin methylesterases: Cell wall enzymes with important roles in plant physiology. Trends Plant Sci., 6: 414-419.
CrossRefDirect Link - Cacao, S.M.B., T.F. Leite, I.G.F. Budzinski, T.B. dos Santos and M.B.S. Scholz et al., 2012. Gene expression and enzymatic activity of pectin methylesterase during fruit development and ripening in Coffea arabica L. Genet. Mol. Res., 11: 3186-3197.
CrossRefPubMedDirect Link - Mercadante, D., L.D. Melton, G.B. Jameson and M.A. Williams, 2014. Processive pectin methylesterases: The role of electrostatic potential, breathing motions and bond cleavage in the rectification of brownian motions. PLoS ONE, Vol. 9.
CrossRef - Johansson, K., M. El-Ahmad, R. Friemann, H. Jornvall, O. Markovic and H. Eklund, 2002. Crystal structure of plant pectin methylesterase. FEBS Lett., 514: 243-249.
CrossRefDirect Link - Giovane, A., L. Servillo, C. Balestrieri, A. Raiola and R. D'avino et al., 2004. Pectin methylesterase inhibitor. Biochimica Biophysica Acta (BBA)-Proteins Proteomics, 1696: 245-252.
CrossRefDirect Link - Corredig, M., W. Kerr and L. Wicker, 2000. Separation of thermostable pectinmethylesterase from marsh grapefruit pulp. J. Agric. Food. Chem., 48: 4918-4923.
CrossRefPubMedDirect Link - Denes, J.M., A. Baron, C.M. Renard, C. Pean and J.F. Drilleau, 2000. Different action patterns for apple pectin methylesterase at pH 7.0 and 4.5. Carbohydr. Res., 327: 385-393.
CrossRefDirect Link - Arabidopsis Genome Initiative, 2000. Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature, 408: 796-815.
CrossRefPubMedDirect Link - Shen, Z., K. Pappan, N.S. Mutti, Q.J. He and M. Denton et al., 2005. Pectinmethylesterase from the rice weevil, Sitophilus oryzae: CDNA isolation and sequencing, genetic origin and expression of the recombinant enzyme. J. Insect Sci., Vol. 5.
Direct Link - Al-Qsous, S., E. Carpentier, D. Klein-Eude, C. Burel and A. Mareck et al., 2004. Identification and isolation of a pectin methylesterase isoform that could be involved in flax cell wall stiffening. Planta, 219: 369-378.
CrossRefDirect Link - Sobry, S., A. Havelange and P. van Cutsem, 2005. Immunocytochemistry of pectins in shoot apical meristems: Consequences for intercellular adhesion. Protoplasma, 225: 15-22.
CrossRefDirect Link - Ren, C. and A.R. Kermode, 2000. An increase in pectin methyl esterase activity accompanies dormancy breakage and germination of yellow cedar seeds. Plant Physiol., 124: 231-242.
CrossRefDirect Link - Hasunuma, T., E.I. Fukusaki and A. Kobayashi, 2004. Expression of fungal pectin methylesterase in transgenic tobacco leads to alteration in cell wall metabolism and a dwarf phenotype. J. Biotechnol., 111: 241-251.
CrossRefDirect Link - Saher, S., A. Piqueras, E. Hellin and E. Olmos, 2005. Pectin methyl esterases and pectins in normal and hyperhydric shoots of carnation cultured in vitro. Plant Physiol. Biochem., 43: 155-159.
CrossRefDirect Link - Pilling, J., L. Willmitzer, H. Bucking and J. Fisahn, 2004. Inhibition of a ubiquitously expressed pectin methyl esterase in Solanum tuberosum L. affects plant growth, leaf growth polarity and ion partitioning. Planta, 219: 32-40.
CrossRefDirect Link - Ferrandiz, C., 2002. Regulation of fruit dehiscence in Arabidopsis. J. Exp. Bot., 53: 2031-2038.
CrossRefDirect Link - Brummell, D.A., V. Dal Cin, C.H. Crisosto and J.M. Labavitch, 2004. Cell wall metabolism during maturation, ripening and senescence of peach fruit. J. Exp. Bot., 55: 2029-2039.
CrossRefDirect Link - Eriksson, E.M., A. Bovy, K. Manning, L. Harrison and J. Andrews et al., 2004. Effect of the Colorless non-ripening mutation on cell wall biochemistry and gene expression during tomato fruit development and ripening. Plant Physiol., 136: 4184-4197.
CrossRefDirect Link - Arancibia, R.A. and C.E. Motsenbocker, 2006. Pectin methylesterase activity in vivo differs from activity in vitro and enhances polygalacturonase-mediated pectin degradation in tabasco pepper. J. Plant Physiol., 163: 488-496.
CrossRefDirect Link - Geisler-Lee, J., M. Geisler, P.M. Coutinho, B. Segerman and N. Nishikubo et al., 2006. Poplar carbohydrate-active enzymes. Gene identification and expression analyses. Plant Physiol., 140: 946-962.
CrossRefPubMedDirect Link - Hertzberg, M., H. Aspeborg, J. Schrader, A. Andersson and R. Erlandsson et al., 2001. A transcriptional roadmap to wood formation. Proc. Natl. Acad. Sci. USA., 98: 14732-14737.
CrossRefDirect Link - Dunand, C., M. Tognolli, S. Overney, L. von Tobel, M. de Meyer, P. Simon and C. Penel, 2002. Identification and characterisation of Ca2+-pectate binding peroxidases in Arabidopsis thaliana. J. Plant Physiol., 159: 1165-1171.
CrossRefDirect Link - Wi, S.G., A.P. Singh, K.H. Lee and Y.S. Kim, 2005. The pattern of distribution of pectin, peroxidase and lignin in the middle lamella of secondary xylem fibres in alfalfa (Medicago sativa). Ann. Bot., 95: 863-868.
CrossRefDirect Link - Lairez, D., B. Cathala, B. Monties, F. Bedos-Belval, H. Duran and L. Gorrichon, 2005. Aggregation during coniferyl alcohol polymerization in pectin solution: A biomimetic approach of the first steps of lignification. Biomacromolecules, 6: 763-774.
CrossRefDirect Link - Francis, K.E., S.Y. Lam and G.P. Copenhaver, 2006. Separation of Arabidopsis pollen tetrads is regulated by QUARTET1, a pectin methylesterase gene. Plant Physiol., 142: 1004-1013.
CrossRefDirect Link - Bosch, M. and P.K. Hepler, 2006. Silencing of the tobacco pollen pectin methylesterase NtPPME1 results in retarded in vivo pollen tube growth. Planta, 223: 736-745.
CrossRefDirect Link - Bosch, M., A.Y. Cheung and P.K. Hepler, 2005. Pectin methylesterase, a regulator of pollen tube growth. Plant Physiol., 138: 1334-1346.
CrossRefDirect Link - Rodriguez-Llorente, I.D., J. Perez-Hormaeche, K. El Mounadi, M. Dary and M.A. Caviedes et al., 2004. From pollen tubes to infection threads: Recruitment of Medicago floral pectic genes for symbiosis. Plant J., 39: 587-598.
CrossRefDirect Link - Lievens, S., S. Goormachtig, S. Herman and M. Holsters, 2002. Patterns of pectin methylesterase transcripts in developing stem nodules of Sesbania rostrata. Mol. Plant-Microbe Interact., 15: 164-168.
CrossRefDirect Link - Vorwerk, S., S. Somerville and C. Somerville, 2004. The role of plant cell wall polysaccharide composition in disease resistance. Trends Plant Sci., 9: 203-209.
CrossRefDirect Link - De Paepe, A., M. Vuylsteke, P. van Hummelen, M. Zabeau and D. van der Straeten, 2004. Transcriptional profiling by cDNA-AFLP and microarray analysis reveals novel insights into the early response to ethylene in Arabidopsis. Plant J., 39: 537-559.
CrossRefDirect Link - Lee, J.Y. and D.H. Lee, 2003. Use of serial analysis of gene expression technology to reveal changes in gene expression in Arabidopsis pollen undergoing cold stress. Plant Pysiol., 132: 517-529.
CrossRefDirect Link - Moscatiello, R., P. Mariani, D. Sanders and F.J.M. Maathuis, 2006. Transcriptional analysis of calcium-dependent and calcium-independent signalling pathways induced by oligogalacturonides. J. Exp. Bot., 57: 2847-2865.
CrossRefDirect Link - Schmohl, N., J. Pilling, J. Fisahn and W.J. Horst, 2000. Pectin methylesterase modulates aluminium sensitivity in Zea mays and Solanum tuberosum. Physiologia Plantarum, 109: 419-427.
CrossRefDirect Link - Dorokhov, Y.L., O.Y. Frolova, E.V. Skurat, P.A. Ivanov and T.V. Gasanova et al., 2006. A novel function for a ubiquitous plant enzyme pectin methylesterase: The enhancer of RNA silencing. FEBS Lett., 580: 3872-3878.
CrossRefDirect Link - Chen, M.H., J. Sheng, G. Hind, A.K. Handa and V. Citovsky, 2000. Interaction between the tobacco mosaic virus movement protein and host cell pectin methylesterases is required for viral cell‐to‐cell movement. EMBO J., 19: 913-920.
CrossRefDirect Link - Penuelas, J., I. Filella, C. Stefanescu and J. Llusia, 2005. Caterpillars of Euphydryas aurinia (Lepidoptera: Nymphalidae) feeding on Succisa pratensis leaves induce large foliar emissions of methanol. New Phytol., 167: 851-857.
CrossRefDirect Link - Von Dahl, C.C., M. Havecker, R. Schlogl and I.T. Baldwin, 2006. Caterpillar-elicited methanol emission: A new signal in plant-herbivore interactions? Plant J., 46: 948-960.
CrossRefDirect Link - Divol, F., F. Vilaine, S. Thibivilliers, J. Amselem, J.C. Palauqui, C. Kusiak and S. Dinant, 2005. Systemic response to aphid infestation by Myzus persicae in the phloem of Apium graveolens. Plant Mol. Biol., 57: 517-540.
CrossRefPubMedDirect Link - Hourton-Cabassa, C., F. Ambard-Bretteville, F. Moreau, J.D. de Virville, R. Remy and C.C. des Francs-Small, 1998. Stress induction of mitochondrial formate dehydrogenase in potato leaves. Plant Physiol., 116: 627-635.
PubMedDirect Link - Hammond-Kosack, K.E. and J.E. Parker, 2003. Deciphering plant-pathogen communication: Fresh perspectives for molecular resistance breeding. Curr. Opin. Biotechnol., 14: 177-193.
CrossRefDirect Link - Valette-Collet, O., A. Cimerman, P. Reignault, C. Levis and M. Boccara, 2003. Disruption of Botrytis cinerea pectin methylesterase gene Bcpme1 reduces virulence on several host plants. Mol. Plant-Microbe Interact., 16: 360-367.
CrossRefDirect Link - D'Ovidio, R., B. Mattei, S. Roberti and D. Bellincampi, 2004. Polygalacturonases, polygalacturonase-inhibiting proteins and pectic oligomers in plant-pathogen interactions. Biochimica Biophysica Acta (BBA)-Proteins Proteomics, 1696: 237-244.
CrossRefDirect Link - Bethke, G., R.E. Grundman, S. Sreekanta, W. Truman, F. Katagiri and J. Glazebrook, 2013. Arabidopsis Pectin methylesterases contribute to immunity against Pseudomonas syringae. Plant Physiol., 164: 1093-1197.
CrossRefDirect Link - Ma, L., S. Jiang, G. Lin, J. Cai and X. Ye et al., 2013. Wound-induced pectin methylesterases enhance banana (Musa spp. AAA) susceptibility to Fusarium oxysporum f. sp. cubense. J. Exp. Bot., 64: 2219-2229.
CrossRefDirect Link - Parre, E. and A. Geitmann, 2005. Pectin and the role of the physical properties of the cell wall in pollen tube growth of Solanum chacoense. Planta, 220: 582-592.
CrossRefDirect Link - Holdaway-Clarke, T.L. and P.K. Hepler, 2003. Control of pollen tube growth: Role of ion gradients and fluxes. New Phytol., 159: 539-563.
CrossRefDirect Link - Iwano, M., H. Shiba, T. Miwa, F.S. Che and S. Takayama et al., 2004. Ca2+ dynamics in a pollen grain and papilla cell during pollination of Arabidopsis. Plant Physiol., 136: 3562-3571.
CrossRefDirect Link - Pina, C., F. Pinto, J.A. Feijo and J.D. Becker, 2005. Gene family analysis of the Arabidopsis pollen transcriptome reveals biological implications for cell growth, division control and gene expression regulation. Plant Physiol., 138: 744-756.
CrossRefDirect Link - Cardenas, L., A. Lovy-Wheeler, K.L. Wilsen and P.K. Hepler, 2005. Actin polymerization promotes the reversal of streaming in the apex of pollen tubes. Cytoskeleton, 61: 112-127.
CrossRefDirect Link - Barry, C.S., M.I. Llop-Tous and D. Grierson, 2000. The regulation of 1-aminocyclopropane-1-carboxylic acid synthase gene expression during the transition from system-1 to system-2 ethylene synthesis in tomato. Plant Physiol., 123: 979-986.
CrossRefDirect Link - Kevany, B.M., M.G. Taylor and H.J. Klee, 2008. Fruit-specific suppression of the ethylene receptor LeETR4 results in early-ripening tomato fruit. Plant Biotechnol. J., 6: 295-300.
CrossRefDirect Link - Bouzayen, M., A. Latche, P. Nath and J.C. Pech, 2010. Mechanism of Fruit Ripening. In: Plant Development Biology-Biotechnological Perspectives, Pua, E.C. and M.R. Davey (Eds.), Springer, Berlin, Heidelberg, ISBN: 978-3-642-02301-9, pp: 319-339.
CrossRefDirect Link - Picton, S., S.L. Barton, M. Bouzayen, A.J. Hamilton and D. Grierson, 1993. Altered fruit ripening and leaf senescence in tomatoes expressing an antisense ethylene-forming enzyme transgene. Plant J., 3: 469-481.
CrossRefDirect Link - Dandekar, A.M., G. Teo, B.G. Defilippi, S.L. Uratsu and A.J. Passey et al., 2004. Effect of down-regulation of ethylene biosynthesis on fruit flavor complex in apple fruit. Transgenic Res., 13: 373-384.
CrossRefDirect Link - Ayub, R., M. Guis, M.B. Amor, L. Gillot and J.P. Roustan et al., 1996. Expression of ACC oxidase antisense gene inhibits ripening of cantaloupe melon fruits. Nat. Biotechnol., 14: 862-866.
CrossRefDirect Link - Flores, F., M. Ben Amor, B. Jones, J.C. Pech, M. Bouzayen, A. Latche and F. Romojaro, 2001. The use of ethylene-suppressed lines to assess differential sensitivity to ethylene of the various ripening pathways in Cantaloupe melons. Physiologia Plantarum, 113: 128-133.
CrossRefDirect Link - Pech, J.C., M. Bouzayen and A. Latche, 2008. Climacteric fruit ripening: Ethylene-dependent and independent regulation of ripening pathways in melon fruit. Plant Sci., 175: 114-120.
CrossRefDirect Link - Mailhac, N. and C. Chervin, 2006. Ethylene and grape berry ripening. Stewart Postharvest Rev., 2: 1-5.
Direct Link - Leclercq, J., L. Adams-Phillips, H. Zegzouti, B. Jones and A. Latche et al., 2002. LeCTR1, a tomato CTR1-like gene, demonstrates ethylene signaling ability in Arabidopsis and novel expression patterns in tomato. Plant Physiol., 130: 1132-1142.
CrossRefDirect Link - Verma, C., S. Gupta, R.K. Singh and S. Mishra, 2013. In silico study of pectin methylesterase in Musa acuminata for delayed ripening. Sci. Int., 1: 175-182.
Direct Link - Verma, C., R.K. Singh, R.B. Singh and S. Mishra, 2015. Biochemical and in-silico studies on pectin methylesterase from G9 variety of musa acuminata for delayed ripening. Open Biochem. J., 9: 15-23.
CrossRefDirect Link